Introduction
Global statistics show that ~49% of malignant brain
tumors are glioblastomas and 30% are diffuse infiltrating low-grade
gliomas (1). The 2021 World Health
Organization (WHO) classification central nervous system (CNS)5 is
a new system that combines the histological features and molecular
phenotypes of tumors. According to this classification system,
gliomas can be divided into adult diffuse gliomas, diffuse
low-grade gliomas in children, diffuse high-grade gliomas in
children and localized astrocyte gliomas. Furthermore, gliomas are
divided into four grades based on the degree of malignancy: WHO
Grades I and II are classified as low-grade gliomas, whereas WHO
Grades III and IV are classified as high-grade gliomas (2). High-grade gliomas, such as
glioblastoma, are associated with poor outcomes and a 5-year
relative survival rate of <6.9% (3). Thus, early identification and
treatment are crucial for improving the prognosis of patients with
gliomas (4).
Traditional glioma treatment typically involves
surgery, chemotherapy and radiation. The biocomplexity of glioma
cells, including their heterogeneity, high proliferation rate and
infiltration, contributes to their high recurrence and drug
resistance. Despite continual improvement in the understanding of
the pathological mechanism of glioma, the quality of life of
patients remains dismal (5). Thus,
identifying the most effective targets and treatments for glioma is
essential.
In addition to the challenges of disease progression
and recurrence, psychological problems, such as anxiety and
depression, often characterize patients with gliomas (6). Individuals with anxiety disorders
frequently exhibit symptoms of depression, and those with
depression frequently experience anxiety disorders. The two
conditions can occur concurrently, and individuals can meet the
criteria for both. These two widely prevalent psychiatric disorders
substantially contribute to morbidity and mortality worldwide.
Comorbid depression and anxiety disorders occur in up to 25% of
general practice patients in Australia (7). Comorbid anxiety and depression are
particularly attributed to shared predisposition or the possibility
that one condition can manifest as an epiphenomenon of the other
(8). A previous study reported the
association of neuroinflammation, oxidative stress and nitrosative
stress with depression and its comorbidities (9). Moderate to severe depression and
anxiety were diagnosed in 28 and 36% of patients with glioma,
respectively (10). Although their
precise causes remain unknown, anxiety and depression remain severe
problems in patients with glioma (11). Furthermore, a retrospective study
illustrated that patients with glioma and depression are associated
with shorter overall survival (OS) when compared with patients
without depression, with median survival times of 7 and 11 months,
respectively. Preoperative depression has been identified as an
independent factor predicting poorer OS in patients with
glioma.
Ferroptosis is a novel form of iron-dependent
programmed cell death characterized by specific hallmarks, such as
glutathione depletion, decreased glutathione peroxidase activity
and the inability of lipid oxides to be processed through the
glutathione peroxidase (GPX) pathway, which is catalyzed by GPX4.
Ferroptosis is promoted by the oxidation of lipids by divalent iron
ions, resulting in the generation of reactive oxygen species (ROS)
(12). Recent studies have reported
that ferroptosis is a significant factor in certain malignancies
and degenerative disorders, including gliomas (13), triple-negative breast cancer
(14), colorectal cancer (15), liver cancer (16) and kidney cancer (17). Regulating ferroptosis is a potential
therapeutic approach towards the development of new treatment
modalities.
Iron metabolism
Iron is indispensable to life, having a crucial role
in numerous physiological activities. Specifically, iron-containing
enzymes are involved in key physiological activities, such as ATP
production, DNA synthesis and oxygen transport (18). Furthermore, iron serves an essential
role in brain development and function, and is involved in various
biological processes, such as embryonic neuronal development,
myelination, neurotransmitter synthesis and oxidative
phosphorylation (19). Iron is also
vital for the activity of enzymes involved in the production of
monoamines (such as dopamine, epinephrine, norepinephrine and
serotonin), which are associated with social-emotional development,
executive functioning and memory processes. However, iron
deficiency compromises the activities of iron-dependent enzymes in
all tissues. Furthermore, although iron promotes cell division and
growth, it can also lead to cell damage, as excessive iron
accumulation can be toxic, activating cell death signaling pathways
through oxidative stress (20).
Previous studies have demonstrated a relationship
between iron metabolism dysregulation and certain illnesses,
including cancer, neurological diseases and atherosclerosis
(21,22). To maintain adequate and safe iron
levels, cells express diverse coordinated proteins that strictly
regulate both intracellular and systemic iron metabolism. Among
these proteins, iron transporters play a crucial role in
controlling iron absorption, storage, distribution and overall iron
homeostasis (23).
Iron absorption
The apical brush border membranes of the small
intestine absorb both heme and non-heme iron. To facilitate this
process, ferric reductase duodenal cytochrome b converts non-heme
Fe3+ entering the colon cells to ferrous ions
(Fe2+). Divalent metal transporter 1 (DMT1), a
proton-coupled transporter found on the apical membrane of
intestinal epithelial cells, absorbs dietary non-heme iron (mainly
ferric, Fe3+) (24).
Iron-chaperone poly(C)-binding protein 2 (PCBP2) mediates the
transfer of iron to ferritin (25).
PCBP2 binds to DMT1 and ferroportin (FPN), encoded by the solute
carrier family (SLC)40 member A1 gene] to facilitate the export of
iron from the epithelial cell layer into the bloodstream (Fig. 1A) (24,26–30).
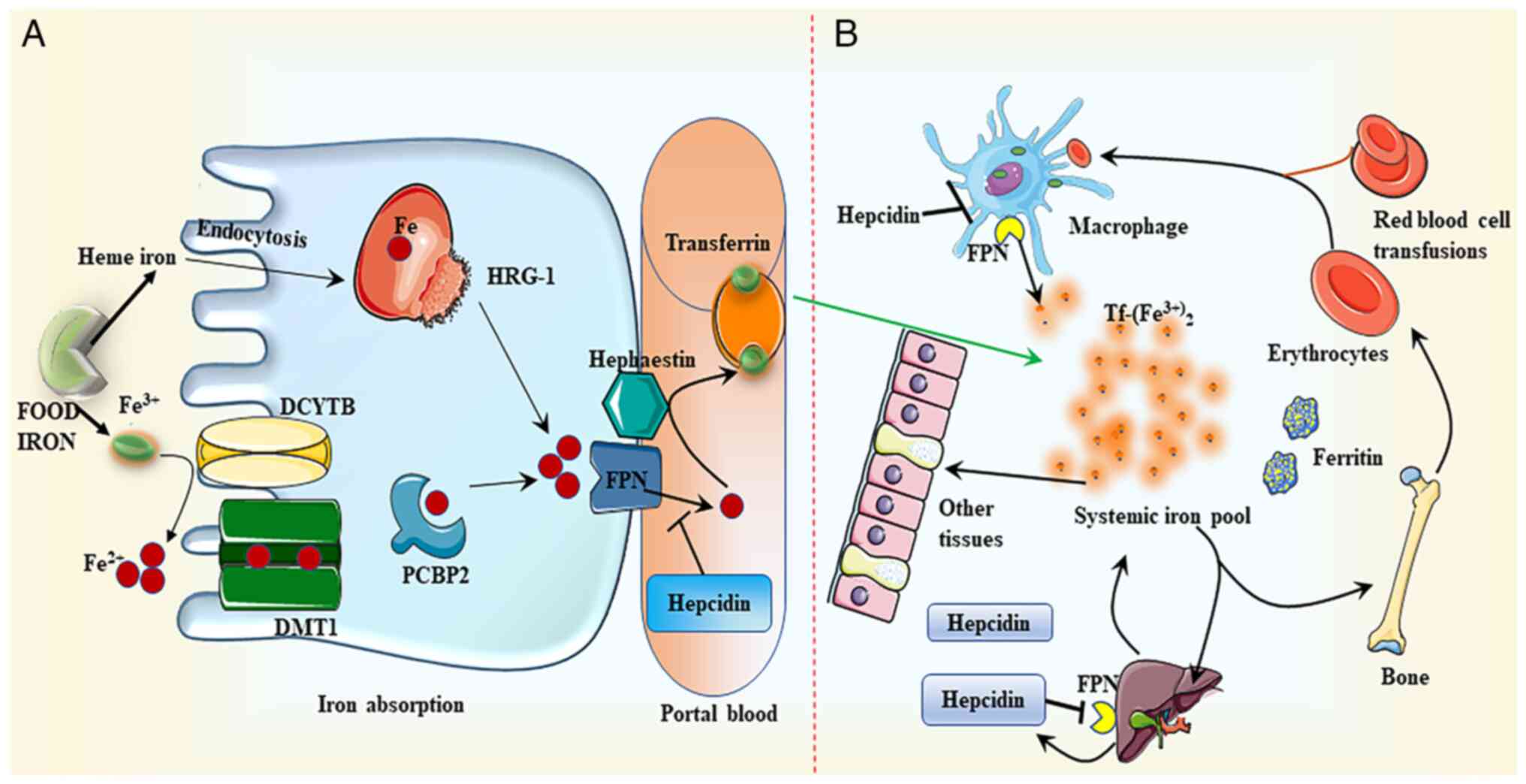 | Figure 1.Iron metabolism. (A) Iron absorption.
DCYTB mediates the conversion of dietary non-heme iron
(Fe3+) to ferrous ion (Fe2+), which is then
absorbed by DMT1 in the membrane of intestinal epithelial cells.
Following this intake, iron is either retained in ferritin or
transferred to the basement membrane by the iron-chaperone PCBP2,
where it is subsequently converted to Fe3+ by
hephaestin. Finally, iron is discharged to the portal circulation
through FPN. Tf attaches to the exported iron, which is then
transported to numerous peripheral tissues. HRG-1 may uptake heme
through endocytosis. Heme is degraded by heme oxygenase once it has
been absorbed. Similar to the transport of non-heme iron, iron
liberated from heme is transferred to portal vein blood through
FPN. (B) Iron distribution. Hepcidin regulates the production of
FPN by directly binding to FPN and promoting its breakdown, thus
facilitating the transport of iron to the portal vein. Iron is
mainly distributed in red blood cells, which transport oxygen in
the blood, and tissues, such as muscle, liver and bone marrow.
DCYTB, duodenal cytochrome b; DMT1, divalent metal transporter 1;
PCBP2, poly(C)-binding protein 2; FPN, ferroportin; HRG-1, heme
responsive gene-1; Tf, transferrin. |
Although the mechanisms of heme iron absorption are
not entirely clear, two types of carrier proteins are hypothesized
to be involved. Firstly, heme carrier protein 1 (HCP1) has
initially been associated with iron absorption. However, HCP1 has
been identified to exhibit high affinity for folate, and primarily
functions as a folate transporter instead of an iron transporter
(31). Secondly, the role of
heme-responsive gene 1 (HRG-1) has gained attention. HRG-1 exhibits
a high sensitivity to heme and may activate the endocytosis pathway
for heme trafficking into the cytosol (Fig. 1) (32). Next, heme is broken down by heme
oxygenase, producing Fe2+, which is then metabolized in
the same pathway as non-heme iron (33).
Iron transport
Cells primarily absorb plasma transferrin (Tf)-bound
iron through Tf receptor (R)1. The interaction between
Fe2+ and TfR1 at the plasma membrane induces
receptor-mediated endocytosis of the Tf/TfR1 complex. In the
nucleus, DMT1 transfers iron from the Tf/TfR1 complex to the
cytoplasm after reduction by the prostate epithelial transmembrane
protein, converting it to Fe2+. Tf/TfR1 is then recycled
to the cell surface and released into the plasma. After
transportation to the peripheral blood, hephaestin converts
Fe2+ to Fe3+. Iron is chelated and
redox-active in the labile iron pool (LIP) within the cell. Iron
from the LIP is transported to different cell regions to meet
metabolic demands or stored in ferritin (Fig. 1) (34).
The blood-brain barrier (BBB) and
blood-cerebrospinal fluid (CSF) barrier (BCSFB) separate the
central nervous system from the dynamic environment of the
bloodstream (35). The BBB is a
complicated multicellular system composed of endothelial cells and
a distinct basement membrane (36–38).
The BCSFB is located in the choroid plexus of the lateral, third
and fourth cerebral ventricles. A carrier system of choroid plexus
epithelial cells can facilitate material exchange between CSF and
blood. Thus, the transportation of iron across the BBB and BCSFB is
necessary for iron to enter the brain (39,40).
The Tf/TfR1 pathway is a potential route through
which iron transporters can cross the luminal (apical) membrane of
the BBB. Hephaestin (HEPH) and FPN1/ceruloplasmin (CP) mediate the
transport of iron through the luminal membrane (41,42).
The Tf/TfR1/DMT1 pathway is a key iron transport mechanism through
the BCSFB. Moreover, the FPN1/CP or FPN1/HEPH pathway mediates iron
export from the choroidal epithelium to the CSF. The degree of TfR1
expression in neurons is proportional to their iron needs. Neurons
express DMT1, which transfers iron from Tf to the cytoplasm
(43). Furthermore, DMT1 is
implicated in the iron uptake of hippocampal neurons during
maturation and memory formation (44,45).
Astrocytes in the central nervous system engage in
intracellular iron transport to maintain extracellular iron
balance. DMT1 may mediate this uptake, and FPN1 and CP have been
demonstrated to be highly expressed on astrocyte membranes; these
two proteins may be necessary for iron to leave astrocytes and
enter the extracellular brain region (46,47).
Lipid metabolism
Lipid peroxidation serves as a hallmark of
ferroptosis, a process that directly damages cell membranes and
leads to ferroptotic cell death. This buildup of lipid peroxides in
cell membranes is initiated by the inhibition of glutathione (GSH)
production, import of cysteine from the extracellular environment
and direct inhibition of GPX4 activity. Lipid peroxidation
predominantly occurs via enzymatic and non-enzymatic oxidation
(48). The lipoxygenase (LOX)
family, which oxidizes free and esterified polyunsaturated fatty
acids (PUFAs) to generate peroxide radicals, controls enzymatic
lipid peroxidation. Acyl-CoA synthetase long chain (ACSL) family
member 4 and lysophosphatidylcholine acyltransferase 3 facilitate
the binding of PUFAs to phospholipids, thereby producing PUFA
phospholipids (PUFA-PLs). PUFA-PLs are susceptible to lipid
peroxidation induced by arachidonic acid (A)LOXs, which eventually
results in the loss of the lipid bilayer and impairs cell membrane
function, thus increasing ferroptosis (49).
Radiotherapy can increase the expression of ACSL4,
thereby enhancing ferroptosis in glioma cells. Meanwhile, ACSL1 is
responsible for ferroptosis induced by conjugated linoleic acid. By
contrast, ACSL3 transforms monounsaturated fatty acids into
acyl-CoA esters, which bind to membrane phospholipids, thereby
preventing ferroptosis in cancer cells. Moreover, cancer-associated
fibroblasts secrete microRNA-522 through the exosome pathway, which
inhibits cancer cell ferroptosis by targeting ALOX15 and inhibiting
the accumulation of lipid-ROS. Additionally, vitamin E can suppress
ALOX activity (50). However,
although LOX activators alone do not directly cause ferroptosis, a
previous study reported that ALOXs overexpression enhanced
sensitivity to the small compound erastin. It has been suggested
that overexpression of wild-type ALOX15, rather than ALOX15 with an
N-terminal truncation, enhances iron deposition induced by Erastin
(51). In addition, LOX is crucial
for ferroptosis when GSH is depleted (49). In most cases, LOX may not be the
primary driver of ferroptosis, but it may contribute to the
initiation or spread of damage. The
15-LOX/phosphatidylethanolamine-binding protein 1 (PEBP1) complex
is produced through the interaction of LOX with PEBP1. Ferroptosis
is initiated by the ferroptosis signal molecule
15-hydroperoxy-eicosa-tetraenoyl-phosphatidylethanolamine (49) (Fig.
2).
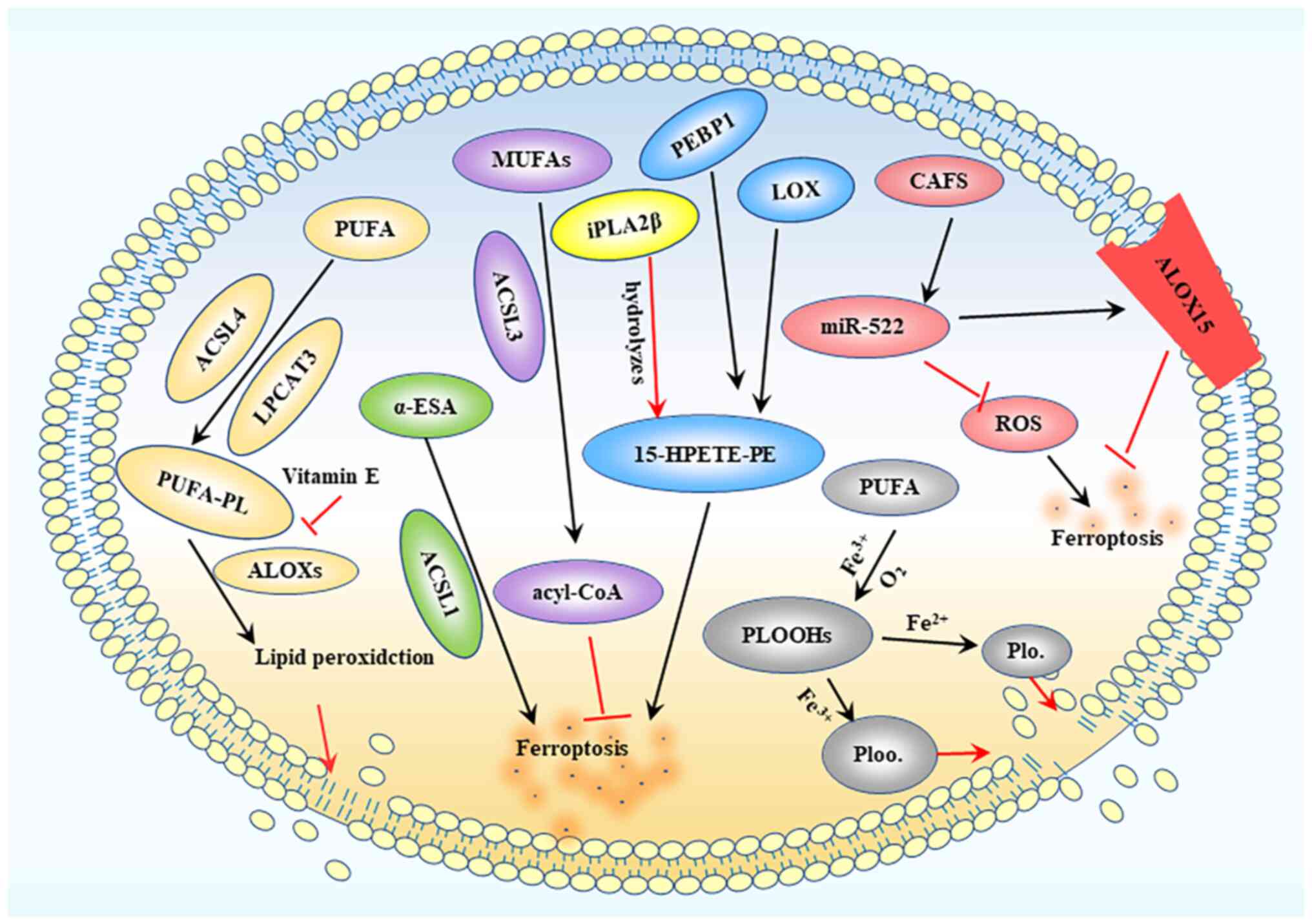 | Figure 2.Lipid peroxidation process. ACSL4 and
LPCAT3 mediate PUFA binding to phospholipids to produce PUFA-PLs,
while ALOXs further induce the production of lipid peroxides,
ultimately destroying the lipid bilayer. MUFA is converted into
acyl-CoA under the action of ACSL3, thereby inhibiting ferroptosis.
LOX interacts with PEBP1 to produce 15-HPETE-PE, leading to
ferroptotic death. CAFs produce extracellular vesicles containing
miR-522, which can inhibit ROS accumulation and target ALOX15 to
inhibit ferroptosis. ACSL, acyl-CoA synthetase long chain; LPCAT3,
lysophosphatidylcholine acyltransferase 3; PUFA, polyunsaturated
fatty acid; PUFA-PL, PUFA phospholipid; MUFA, monounsaturated fatty
acid; LOX, lipoxygenase; ALOX, arachidonic acid LOX; PEBP1,
phosphatidylethanolamine-binding protein 1; 15-HPETE-PE,
15-hydroperoxy-eicosa-tetraenoyl-phosphatidylethanolamine; CAFs,
cancer-associated fibroblasts; miR, microRNA; ROS, reactive oxygen
species; PLOOH, phospholipid hydroperoxide; iPLA2β,
calcium-independent PLA2β; α-ESA, α-eleostearic a. |
Ferroptosis
System xc−
Cysteine deficiency serves a significant role in the
induction of ferroptosis and is a major contributor to ferroptosis
in glioblastoma (52). In the cell
membrane, the amino acid transport system xc−
is composed of two key components, SLC7A11 (xCT) and SLC3A2 (also
known as 4F2 cell-surface antigen heavy chain). This system
facilitates the exchange of glutamate and cysteine. Once inside the
cell, cysteine 2 (Cys2) is converted into cysteine, which
stimulates the production of the GPX4 substrate GSH (53). GPX4, a fundamental regulator of
ferroptosis, may convert GSH to oxidized glutathione and reduce
lipid hydroperoxides to lipid alcohols. This mechanism is essential
for the prevention of lipid peroxidation and inhibition of
ferroptosis (54). Both GPX4
knockdown and inactivation result in ferroptosis (55). By blocking cysteine transport
through system xc−, the ferroptosis inducer
elastin can cause GSH depletion and GPX4 inactivation (56), whereas RAS-selective lethal 3
directly promotes ferroptosis by reducing GPX4 activity. Cysteine
is an essential limiting amino acid for the synthesis of
intracellular GSH, and GPX4 function is immediately affected by GSH
depletion. Thus, system xc− that is
responsible for Cys2 absorption is considered to be one of the most
important regulators of ferroptosis (Fig. 3).
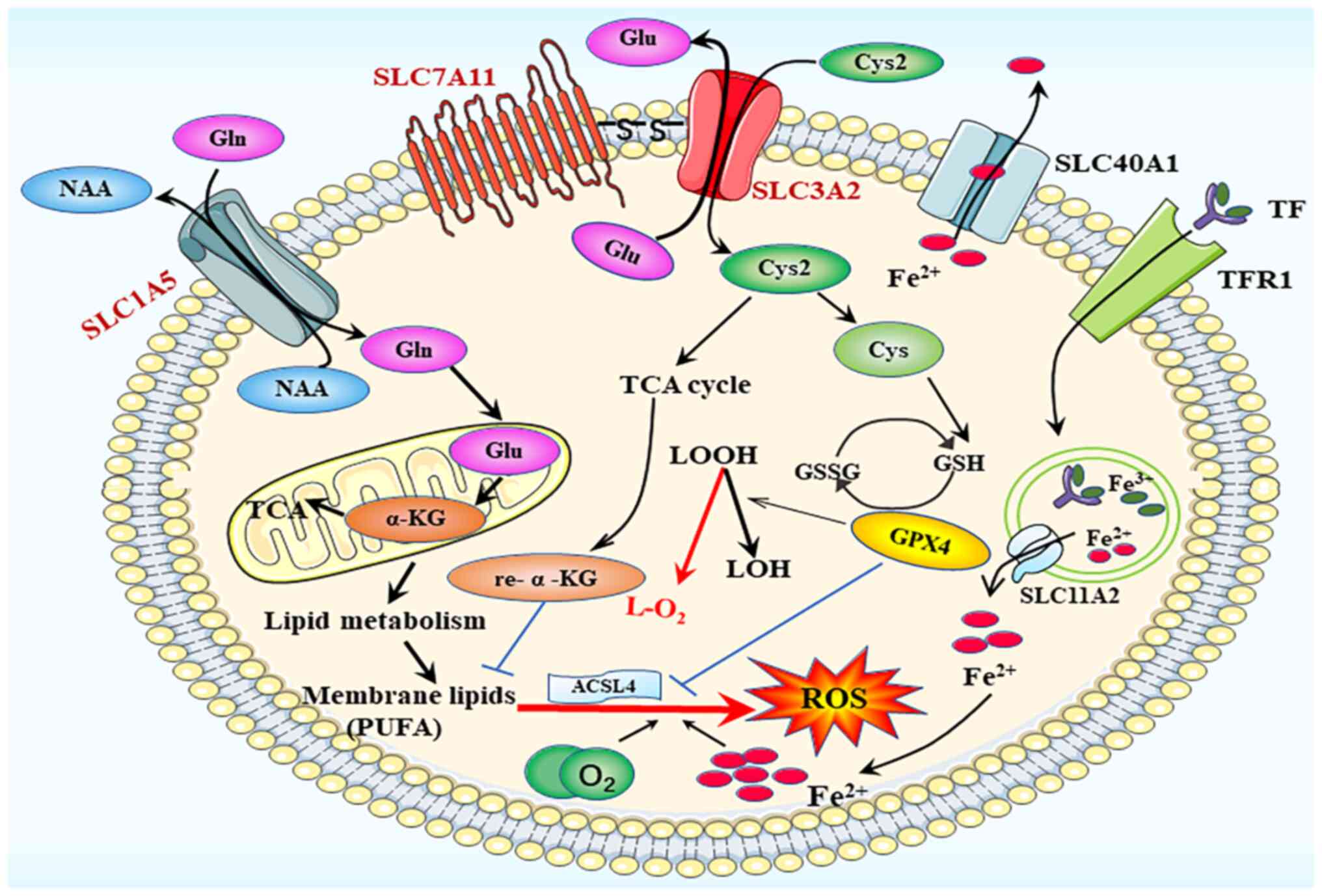 | Figure 3.Ferroptosis process. The main
participating system in ferroptotic death is the amino acid
transport system xc−, composed of the SLC1A5
and SLC3A2 families. SLC, solute carrier family; ACSL4, acyl-CoA
synthetase long chain family member 4; GPX4, glutathione
peroxidase; GSH, glutathione; GSSG, glutathione disulfide; ROS,
reactive oxygen species; TfR1, transferrin receptor 1; α-KG,
α-ketoglutarate; Cys, cysteine; Cys2, cysteine2; Glu, glutamate;
Gln, glutamine; LOH, lipid alcohol; LOOH, lipid hydroperoxidecid;
TCA cycle, tricarboxylic acid cycle. |
SLC1A5 and SLC3A2
SLC1A5 and SLC3A2 are essential proteins for the
transmembrane translocation of glutamine into cells (57). Once inside the cells, glutaminases
are transported to the mitochondria and glutamine is converted into
glutamate and ammonia. Glutamate can then be transformed into
α-ketoglutarate (α-KG), a crucial step in the tricarboxylic acid
cycle (Fig. 3) (58). Dihydrolipoamide dehydrogenase (DLD)
serves a crucial role in the promotion of ferroptosis, especially
in cases of cysteine deficiency or inhibition of cysteine import.
α-KG can stimulate DLD to generate hydrogen peroxide, and it can
also be converted into acetyl-CoA, thereby enhancing fatty acid
production and promoting lipid peroxidation-dependent ferroptosis
(59).
Gene transcriptional regulatory
network
Complex transcriptional regulatory networks affect
the cell vulnerability to ferroptosis. Several transcription
factors have been demonstrated to control certain
ferroptosis-related genes (60).
For instance, the transcription factors tumor protein p53 (TP53),
activating transcription factor (ATF)3, BTB domain and CNC homolog
1 (BACH1) and STAT1 upregulate SLC7A11 and downregulate nuclear
factor erythroid 2-related factor 2 (NFE2L2/Nrf2), ATF4 and aryl
hydrocarbon receptor nuclear translocator-like protein 1. The
intricate roles served by various transcription factors associated
with ferroptosis, including TP53 and NFE2L2, are shown in Fig. 4. TP53 upregulation has been shown to
suppress expression of the system x−c
transporter subunit SLC7A11 and sensitize cells to ferroptosis
(18).
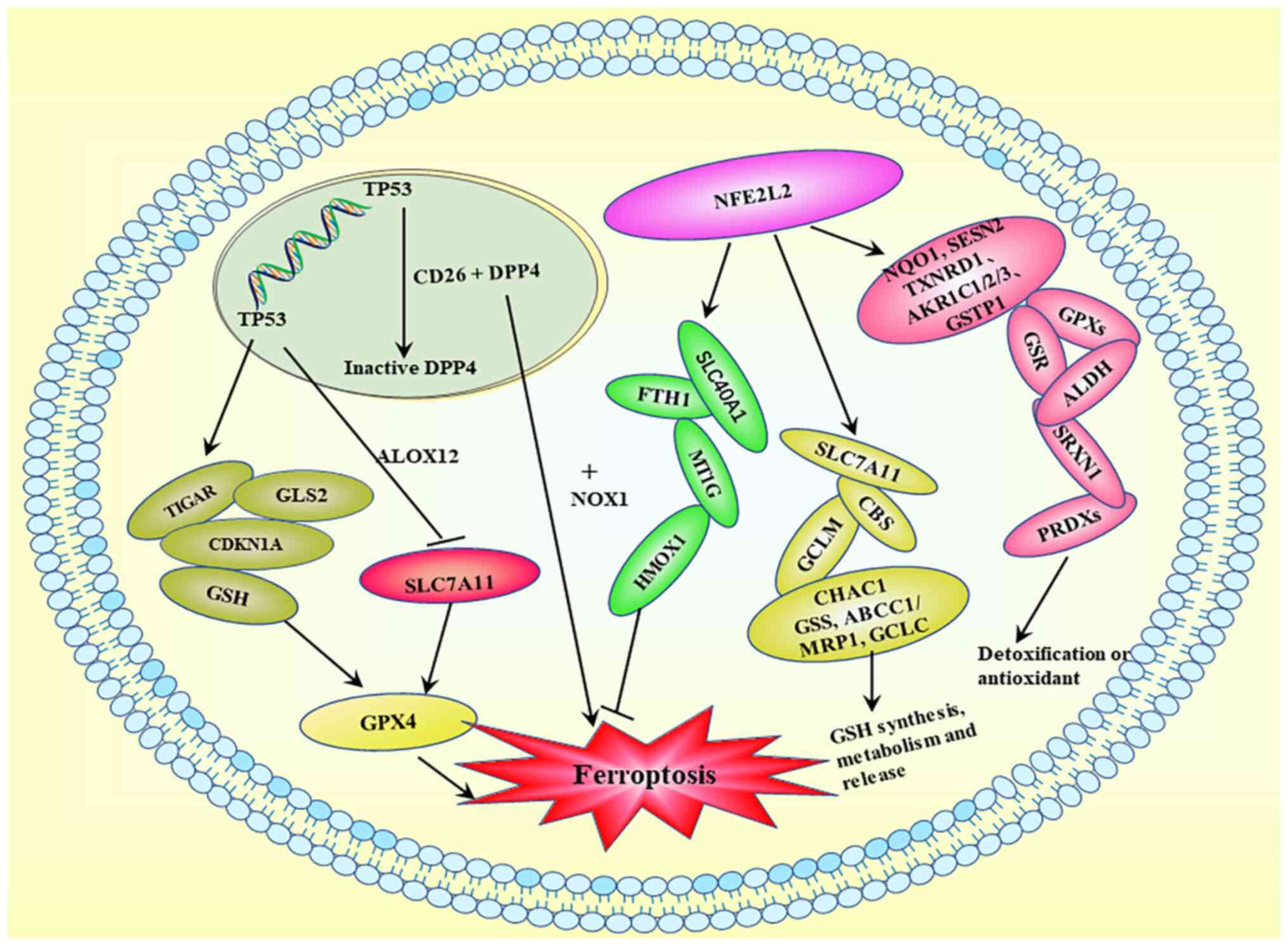 | Figure 4.Ferroptosis-related transcriptional
regulation. Activation of TP53 exerts a minimal effect on GSH
levels or GPX4 function, but it downregulates SLC7A11 and inhibits
cysteine absorption. Activation of TP53 results in the production
of GSH, CDKN1A, GLS2 and TIGAR. DPP4/CD26 interacts with TP53 in
the nucleus and is maintained in a dormant state, whereas TP53
deficiency promotes DDP4 cell membrane localization. DDP4 induces
ferroptosis by binding to NOX1 on the cell membrane. TP53, tumor
protein p53; GSH, glutathione; GPX, glutathione peroxidase; SLC,
solute carrier family; CDKN1A, cyclin-dependent kinase inhibitor
1A; GLS2, glutaminase 2; TIGAR, TP53-induced glycolytic regulatory
phosphatase; DPP4, dipeptidyl peptidase 4; NOX, NADPH oxidase;
NFE2L2, nuclear factor erythroid 2-related factor 2; ALOX,
arachidonic acid lipoxygenase; FTH1, ferritin heavy chain. |
NFE2L2 exerts multiple effects on ferroptosis via
transcriptional regulation. First, it inhibits ferroptosis by
upregulating iron metabolism genes, including FTH1, SLC40A1, HMOX1
and MT1G. Second, NFE2L2 activation enhances the expression of
SLC7A11, GCLM, CBS, CHAC1, ABCC1/MRP1, GCLC and GSS, which are
involved in GSH production, metabolism and release. Third, several
NFE2L2 target genes, including NQO1, TXNRD1, AKR1C1/2/3, SESN2,
GSTP1, GPXs, GSR, SRXN1, ALDH and PRDXs, participate in
detoxification or antioxidant activities, which may reduce
ferroptosis sensitivity. Therefore, NFE2L2 serves a crucial role in
the transcriptional regulation of ferroptosis, primarily via the
activation of genes that counteract cellular damage (61).
Regulation of iron homeostasis
The maintenance of iron homeostasis is crucial.
Hepcidin, a 25-amino acid peptide hormone synthesized and released
by the liver, serves a key role in the regulation of iron storage,
distribution and consumption. It regulates FPN production by
directly binding to FPN and promoting its breakdown (62). An excessive increase in iron levels
induces the production of hepcidin, which destroys FPN in
intestinal epithelial cells, thus reducing plasma iron levels. By
contrast, under iron-deficient conditions, hepcidin levels decrease
to maintain FPN expression, facilitating the release of iron into
the plasma (63). As
aforementioned, DMT1 and TfR1 are essential proteins found in cells
that help regulate intracellular iron concentration, thereby
serving a vital role in iron homeostasis. Elevated iron levels can
lead to the formation of hydroxyl radicals through the Fenton
reaction, particularly when combined with hydrogen peroxide. This
process induces the oxidation of PUFAs in cell membranes,
substantially accelerating lipid peroxidation and ultimately
causing cell damage or death (64).
In the brain, the BBB and BCSFB serve crucial roles
in maintaining the stability of physical and chemical elements in
the brain tissue environment. The BBB is the most significant
barrier in the brain that prevents ~98% of small molecule reagents
from passing when treating CNS-related disease (65). Moreover, these barriers regulate the
transport of iron from the bloodstream to the brain parenchyma,
helping to maintain brain iron levels largely independent of
systemic iron levels and providing protection against systemic iron
toxicity (37,38). In addition, iron ion equilibrium is
maintained by three antioxidant mechanisms, namely GSH, selenium
and Coenzyme Q (CoQ) systems.
GSH system
GSH is an antioxidant tripeptide composed of
glutamic acid, cysteine and glycine. The enzyme glutamate cysteine
ligase continuously catalyzes the production of GSH from glutamic
acid and cysteine. However, the limited availability of cysteine
within cells can decrease GSH production. To counteract this
limitation, system xc− serves as a transport
mechanism during GSH production. Inhibition of systemic
xc-depletes intracellular cysteine, leads to a decrease
in glutathione concentration and triggers oxidative stress, and
increases the sensitivity of cells to ferroptosis (34). GSH serves a vital role in the system
xc−/GSH/GPX4-dependent antioxidant defense
pathway during ferroptosis, and it is essential for the maintenance
of normal GPX4 activity (66,67).
GPX4 has three subtypes: Mitochondrial, cellular solute and
nuclear; it remains unclear which of these subtypes is mainly
involved in the regulation of anti-ferroptotic effects (18).
Selenium system
Selenium, an essential trace element, regulates
cellular redox processes during oxidative stress. In
selenocysteine-containing proteins, such as GPX4, selenium serves a
crucial role in the enhancement of their antioxidant function
during ferroptosis. Selenium deficiency results in cellular death
that depends on lipid peroxidation (68). A previous study reported that sodium
selenite can reduce heme-driven ferroptosis triggered by
homocysteine (a metabolite of methionine) or intracerebral
hemorrhage. and enhanced the transcriptional upregulation of
selenoprotein genes (including GPX4, Selenop and Txnrd1) and GPX3.
However, it remains unclear whether selenoproteins other than GPX4
are also involved in the control of ferroptosis (69).
CoQ system
CoQ, commonly known as ubiquinone, is an
endogenously generated and ubiquitous oxy-benzoquinone molecule. By
promoting the movement of electrons from complexes I and II to
complex III, CoQ10 substantially contributes to the electron
transport chain. In addition, the lipophilic antioxidant CoQ can be
transformed into panthenol, a reduced form that is considered to be
involved in the recovery of other antioxidants (such as ascorbate
and tocopherol), preventing cells from undergoing ferroptosis
without the need for GSH (70).
Apoptosis-inducing factor mitochondria-associated
2/ferroptosis-suppressor protein 1 (FSP1) regulates ferroptosis by
synthesizing CoQH2 (Fig.
5) (71).
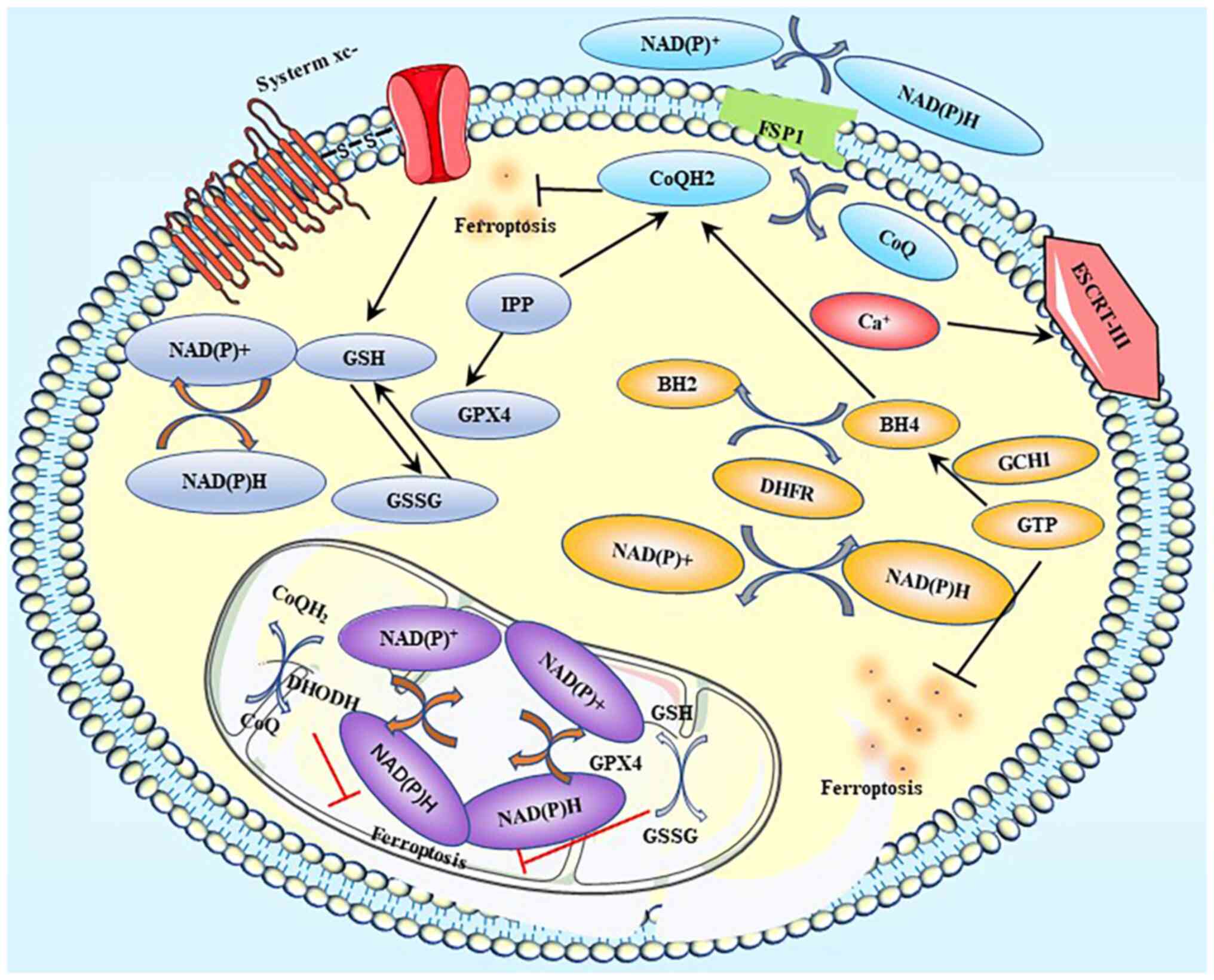 | Figure 5.Ferroptosis-related defense systems.
Label A represents the GPX4-GSH axis; label B represents the
FSP1-CoQ10-NAD(P)H pathway; label C represents the DHODH-mediated
ferroptosis defense; label D represents the GCH1-BH4-DHFR axis; and
label E represents the ESCRT III-mediated plasma membrane repair
system. GPX, glutathione peroxidase; GSH, glutathione; GSSG,
oxidized glutathione; FSP1, ferroptosis suppressor protein 1; CoQ,
coenzyme Q; CoQH2, Coenzyme QH2; DHODH,
dihydroorotate dehydrogenase; GCH1, GTP cyclohydrolase 1; BH4,
tetrahydrobiopterin; BH2, dihydrobiopterin; DHFR, dihydrofolate
reductase; ESCRT, endosomal sorting complex required for transport;
IPP, Isopentenyl pyrophosphate. |
There are at least five defensive mechanisms,
including the GPX4-GSH axis, FSP1-CoQ10-NAD(P)H pathway,
DHODH-mediated ferroptosis defense, GCH1-BH4-DHFR axis, and ESCRT
III-mediated plasma membrane repair system, which are involved in
the reduction or alleviation of the damage caused by ferroptosis
(49) (Fig. 5).
Glioma
Ferroptosis, a form of programmed cell death driven
by iron-dependent lipid peroxidation, has an unclear role in the
death of glioma cells. In addition to depleting GSH and cysteine,
phenylarsine oxide (PAB) has been reported to cause abnormal
elevations in the levels of intracellular ferrous ion,
H2O2 and lipid peroxides, reducing the
survival of glioma cells both in vitro and in vivo.
Deferoxamine, an iron chelator, has been reported to reduce
PAB-induced lipid peroxidation and glioma cell death in
vitro, whereas ferric ammonium citrate could reverse these
effects. Ferrostatin-1- and GSH-induced suppression of lipid
peroxidation inhibited glioblastoma (U87 and U251) cell death
induced by PAB. Treatment with PAB resulted in intact cell
membranes, fewer mitochondria, denser mitochondrial membranes,
normal nuclear size and no chromatin condensation. PAB increased
intracellular iron levels by activating Tf receptors. Excessive
iron supply triggered the activation of NADPH oxidase 4 (NOX4),
leading to excessive H2O2 and lipid peroxide
production. Additionally, PAB increased the production of
H2O2 and lipid peroxides by causing
intracellular GSH depletion via the p53-mediated xCT pathway. Thus,
PAB may induce glioblastoma multiforme cell ferroptosis,
potentially serving as a glioma treatment (72).
Overexpression of the nuclear factor
(erythroid-derived)-like 2 (NFE2L2) or suppression of the
Kelch-like ECH-associated protein 1 (Keap1) has been shown to
promote glioblastoma invasiveness and hence accelerate glioma
growth (72). The Keap1/NFE2L2
signaling pathway can modulate ferroptosis activation (72). High mobility group box 1 protein has
been reported to induce ferroptosis in mesangial cells by
stimulating the Keap1/NFE2L2 signaling pathway (73). By downregulating NFE2L2 and
inhibiting the nuclear receptor subfamily 2 group F member 6/KEAP1
signaling pathway, apatinib has been demonstrated to induce
ferroptosis in glioma cells. Ibuprofen stimulates ferroptosis by
inhibiting Keap1/NFE2L2 signaling. Moreover, NFE2L2 overexpression
or Keap1 knockdown has been reported to expedite the proliferation
and oncogenic transformation of glioblastoma U87 cells (74). Thus, the Keap1/NFE2L2 pathway may
inhibit ferroptosis (75,76).
Similar to NFE2L2, xCT is positively regulated by
ATF4 and serves a pivotal role in reducing ferroptosis-induced ROS
production in glioblastoma multiforme cells. xCT can be induced by
activating ATF4. Continuous targeting to promote ATF4-dependent
processes activates xCT and thus enhances the growth of malignant
gliomas (72). In addition, ATF4
has been reported to promote tumor angiogenesis in gliomas and
affect the vascular architecture in an xCT-dependent manner. In a
manner that is dependent on xCT, pseudolaric acid B also causes
ferroptosis (77).
Ubiquitin thioesterase OTUB1 is abundantly expressed
in gliomas. A novel axis of OTUB1/SLC7A11 contributing to the
stemness of glioblastoma U373, U87 and U251 cell lines has been
reported in a recent study (78).
OTUB1 was demonstrated to stabilize SLC7A11 by directly interacting
with it, while in the absence of OTUB1, ferroptosis was triggered
by SLC7A11 expression.
In addition to limiting proliferation via vitamin C
deficiency and ACSL4 inhibition, ferroptosis can be achieved by
activating the transcription factors BACH1 or NOX4 to increase
oxidative stress, restrict autophagy and trigger cell death. Drugs,
such as 2-nitroimidazoles, temozolomide and artemisinin (and its
derivatives) have been developed based on these principles.
Downregulation of GPX4 and subsequent accumulation of lipid ROS are
the key mechanisms through which dihydroartemisinin triggers
ferroptosis. Ferrostatin-1, a specific inhibitor of ferroptosis,
was reported to reverse all these changes (79). Additionally, temozolomide has been
reported to induce ferroptosis by increasing DMT1 expression in the
TJ905 glioblastoma cell line. Thus, DMT1 may be used as a
therapeutic target for glioblastoma (80–82).
A study examining expression data from The Cancer
Genome Atlas reported that a higher tumor grade and poor prognosis
in patients with glioma were associated with the upregulation of
coatomer subunit ζ-1 (COPZ1). Moreover, ferritin phagocytosis,
which has been linked with the development of cancer and
degenerative diseases, has been associated with nuclear receptor
coactivator 4 (NCOA4) expression (83). Knockdown of COPZ1 can activate
NCOA4, leading to the degradation of ferritin. The Fenton reaction,
which is triggered by large concentrations of divalent iron,
increases ROS production, and lipid peroxidation caused by ROS
eventually leads to ferroptosis. Studies have indicated that
depletion of COPZ1 induces ferroptosis in glioma cells by
increasing NCOA4 and ATG7 levels. Thus, the COPZ1/NCOA4/FTH1 axis
may be a novel therapeutic target in the treatment of glioma
(84).
Anxiety and depression
The development of psychological disorders is a
complex process that often involves the accumulation of multiple
emotional changes instead of a single emotional shift. An increase
in the prevalence of anxiety and depression is typically
accompanied by an increase in suicide rates, which is a major
concern in contemporary society. Depression and anxiety, the two
most prevalent psychological diseases, are responsible for the
morbidity and mortality of millions of individuals worldwide
(85). Numerous biological,
psychological and social environmental variables (86–88)
contribute to the pathophysiology of depression and anxiety, such
as personal experiences, workplace culture and significant life
events. Psychological problems can exert diverse effects on
physical health. Depression and anxiety disorders have been
reported to co-occur with cardiovascular disease, trauma and
numerous forms of cancer, such as lung cancer, breast cancer and
malignant brain tumors (89).
Cancer-related chronic pain and its management may lead to mental
disorders, and depression and anxiety in particular can alter the
synthesis of inflammatory cytokines and chemokines, the metabolism
of neurotransmitters and the function of the neuroendocrine system
(90).
Glioma is a fatal neurological disease associated
with sexual dysfunction, cognitive impairment and possibly death.
It is the most widespread and hazardous form of central nervous
system tumor, account for 40–60% of all primary central nervous
system tumors (91,92). Glioma is often accompanied by
psychological disorders, such as anxiety and depression. In one
study, the prevalence of anxiety disorders and depression in
patients with glioma determined using different diagnostic scales,
such as Hospital Anxiety and Depression Scale and Quality of Life
Core Questionnaire were 36.6–37.4 and 28.4–32.6% (93), respectively. Negative emotions in
response to the tumor and the interplay between the immune, neural
and psychological systems may lead to anxiety and depression in
patients with glioma, and anxiety and depression have been reported
to shorten the survival time in such patients (89,94,95).
However, the precise cause remains unknown, and anxiety and
depression continue to be important problems for patients with
glioma.
A total of 190 patients with glioma treated with
resection were included in a previous study. The hospital anxiety
and depression scale (HADS) and the Zung self-rating anxiety scale
(SAS) were used to assess anxiety, whereas the HADS and the Zung
self-rating depression scale (SDS) were used to assess depression.
All patients were monitored for 36 months or until death. According
to the survival data, OS was calculated, and The HADS for anxiety,
SAS and SDS were associated with a shorter OS, while the HADS for
depression was not (93). Changes
in the levels of receptor-interacting protein kinase 3,
phosphorylated mixed lineage kinase domain-like protein, ferritin
light chain and lipid peroxidation related to ferroptosis were
demonstrated using western blotting and biochemical assays. The
study reported a relationship between ferroptosis and depression,
thereby identifying a possible therapeutic target for depression
(93).
The presence of lipid peroxidation in psychiatric
diseases has been demonstrated through clinical research. High
lipid peroxidation rates were reported to increase the probability
of treatment-resistant depression (96). Recently, edaravone was shown to
ameliorate depression- and anxiety-like behaviors, oxidative stress
and neuroinflammation in mouse models of depression induced by
chronic social defeat stress (97).
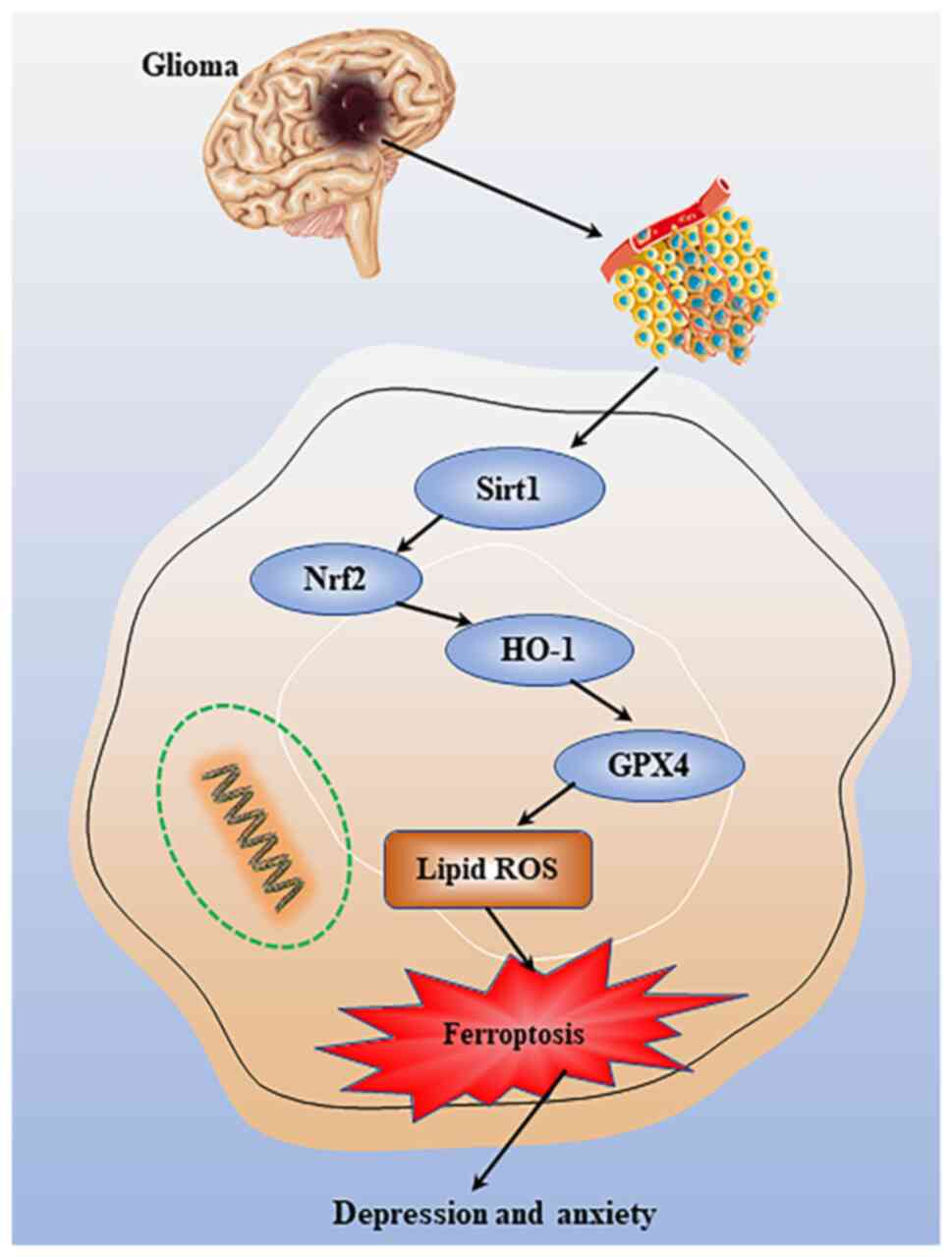
GPX4-mediated ferroptosis may be modulated through
underlying molecular mechanisms involving the sirtuin 1/Nrf2/heme
oxygenase-1 pathway (Fig. 6). This
previous study suggested that abnormal GPX4 expression is a
potential mechanism underlying depression (97). Therefore, GPX4-mediated ferroptosis
may be a promising target for treating major depressive disorder.
Xiao Yao San, a traditional Chinese medicine, was reported to exert
therapeutic effects by increasing the expression of GPX4 and other
ferroptosis-related molecules in the hippocampus of depressed rats
(97). A previous study revealed a
downregulation of ERK levels in the brains of depressed individuals
with suicidal tendencies (98).
Alterations in the expression of PEBP1 may influence the ERK
pathway, potentially contributing to the development of depression
(98).
Conclusion
Ferroptosis occurs due to an imbalance between the
body's antioxidant and oxidative mechanisms. Ferroptosis has been
linked to numerous diseases and systemic conditions, including
certain types of cancer, cardiovascular and digestive disorders.
Moreover, ferroptosis serves a crucial role in the occurrence and
development of gliomas, as well as the metabolic process of anxiety
and depression, which can be caused by cancer. Ferroptosis-related
biomarkers and long non-coding RNAs, such as AP003555.1 and
AC000584.1, have been reported to be useful in predicting prognosis
in patients with gliomas (99).
Anxiety and depression can affect the prognosis of these patients,
shortening their survival time. A recent study proposed that the
WHO classification of gliomas is an independent risk factor for
anxiety (93). However, no
experimental study has elucidated the mechanism underlying glioma
ferroptosis. More thorough research on ferroptosis is needed to
assess its advantages and disadvantages, to enhance patient
survival and quality of life, and to improve long-term clinical
outcomes for patients with gliomas. The present review summarizes
the research progress on ferroptosis in gliomas and its mechanistic
relevance to anxiety and depression. For patients with gliomas, the
focus should not only be on treatment methods, but also on their
quality of life after being diagnosed, with the state of their
mental health being the most concerning and a matter of societal
interest. Patients with neuroglioma have a high incidence of
anxiety and depression, markedly impacting their quality of life.
Therefore, the evaluation of the mechanisms behind anxiety and
depression in patients with glioma can provide a theoretical basis
for the improvement of patient outcomes.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
Not applicable.
Authors' contributions
YB wrote the majority of the manuscript. LM was
involved in topic selection and wrote a draft of the review. ZY, WL
and MJ edited the manuscript. All authors have read and approved
the final manuscript. Data authentication is not applicable.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Schaff LR and Mellinghoff IK: Glioblastoma
and other primary brain malignancies in adults: A review. JAMA.
329:574–587. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
McNamara C, Mankad K, Thust S, Dixon L,
Limback-Stanic C, D'Arco F, Jacques TS and Löbel U: 2021 WHO
classification of tumours of the central nervous system: A review
for the neuroradiologist. Neuroradiology. 64:1919–1950. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Szklener K, Mazurek M, Wieteska M,
Waclawska M, Bilski M and Mandziuk S: New directions in the therapy
of glioblastoma. Cancers (Basel). 14:53772022. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Salvador GA: Iron in neuronal function and
dysfunction. Biofactors. 36:103–110. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chen S and Zhang Z, Zhang B, Huang Q, Liu
Y, Qiu Y, Qiu Y, Long X, Wu M and Zhang Z: CircCDK14 promotes tumor
progression and resists ferroptosis in glioma by regulating PDGFRA.
Int J Biol Sci. 18:841–857. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Jiao JT, Jiang C, Huang J, Dai MC, Wang C,
Cheng C and Shao JF: Metabolic syndrome factors and risk of
postoperative depression in high-grade glioma patients in a
1.5-year prospective study. Med Oncol. 31:2342014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tiller JW: Depression and anxiety. Med J
Aust. 199((S6)): S28–S31. 2013.PubMed/NCBI
|
|
8
|
Middeldorp CM, Cath DC, Van Dyck R and
Boomsma DI: The co-morbidity of anxiety and depression in the
perspective of genetic epidemiology. A review of twin and family
studies. Psychol Med. 35:611–624. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Maes M, Kubera M, Obuchowiczwa E, Goehler
L and Brzeszcz J: Depression's multiple comorbidities explained by
(neuro)inflammatory and oxidative & nitrosative stress
pathways. Neuro Endocrinol Lett. 32:7–24. 2011.PubMed/NCBI
|
|
10
|
Bunevicius A, Deltuva VP and Tamasauskas
A: Association of pre-operative depressive and anxiety symptoms
with five-year survival of glioma and meningioma patients: A
prospective cohort study. Oncotarget. 8:57543–57551. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gathinji M, McGirt MJ, Attenello FJ,
Chaichana KL, Than K, Olivi A, Weingart JD, Brem H and
Quinones-Hinojosa A: Association of preoperative depression and
survival after resection of malignant brain astrocytoma. Surg
Neurol. 71:299–303; discussion 303. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Conrad M, Lorenz SM and Proneth B:
Targeting ferroptosis: New hope for as-yet-incurable diseases.
Trends Mol Med. 27:113–122. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liu T, Zhu C, Chen X, Guan G, Zou C, Shen
S, Wu J, Wang Y, Lin Z, Chen L, et al: Ferroptosis, as the most
enriched programmed cell death process in glioma, induces
immunosuppression and immunotherapy resistance. Neuro Oncol.
24:1113–1125. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wang Z, Li R, Hou N, Zhang J, Wang T, Fan
P, Ji C, Zhang B, Liu L, Wang Y, et al: PRMT5 reduces immunotherapy
efficacy in triple-negative breast cancer by methylating KEAP1 and
inhibiting ferroptosis. J Immunother Cancer. 11:e0068902023.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yan H, Talty R and Johnson CH: Targeting
ferroptosis to treat colorectal cancer. Trends Cell Biol.
33:185–188. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ramadori P, Gallage S and Heikenwalder MF:
Unique tumour microenvironment: When ferroptosis activation boosts
ICI of liver cancer. Gut. 72:1639–1641. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Han Z, Wang H, Long J, Qiu Y and Xing XL:
Establishing a prognostic model of ferroptosis- and immune-related
signatures in kidney cancer: A study based on TCGA and ICGC
databases. Front Oncol. 12:9313832022. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chen X, Li J, Kang R, Klionsky DJ and Tang
D: Ferroptosis: Machinery and regulation. Autophagy. 17:2054–2081.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Dixon SJ, Lemberg KM, Lamprecht MR, Skouta
R, Zaitsev EM, Gleason CE, Patel DN, Bauer AJ, Cantley AM, Yang WS,
et al: Ferroptosis: An iron-dependent form of nonapoptotic cell
death. Cell. 149:1060–1072. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Degterev A, Huang Z, Boyce M, Li Y, Jagtap
P, Mizushima N, Cuny GD, Mitchison TJ, Moskowitz MA and Yuan J:
Chemical inhibitor of nonapoptotic cell death with therapeutic
potential for ischemic brain injury. Nat Chem Biol. 1:112–119.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liu Y, Shoji-Kawata S, Sumpter RM Jr, Wei
Y, Ginet V, Zhang L, Posner B, Tran KA, Green DR, Xavier RJ, et al:
Autosis is a Na+, K+-ATPase-regulated form of cell death triggered
by autophagy-inducing peptides, starvation, and hypoxia-ischemia.
Proc Natl Acad Sci USA. 110:20364–20371. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yang WS, SriRamaratnam R, Welsch ME,
Shimada K, Skouta R, Viswanathan VS, Cheah JH, Clemons PA, Shamji
AF, Clish CB, et al: Regulation of ferroptotic cancer cell death by
GPX4. Cell. 156:317–331. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Xie Y, Hou W, Song X, Yu Y, Huang J, Sun
X, Kang R and Tang D: Ferroptosis: Process and function. Cell Death
Differ. 23:369–379. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Gunshin H, Mackenzie B, Berger UV, Gunshin
Y, Romero MF, Boron WF, Nussberger S, Gollan JL and Hediger MA:
Cloning and characterization of a mammalian proton-coupled
metal-ion transporter. Nature. 388:482–488. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lane DJ and Richardson DR: Chaperone turns
gatekeeper: PCBP2 and DMT1 form an iron-transport pipeline. Biochem
J. 462:e1–e3. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hurrell R and Egli I: Iron bioavailability
and dietary reference values. Am J Clin Nutr. 91:1461S–1467S. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ma S, Henson ES, Chen Y and Gibson SB:
Ferroptosis is induced following siramesine and lapatinib treatment
of breast cancer cells. Cell Death Dis. 7:e23072016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Gao M, Monian P, Quadri N, Ramasamy R and
Jiang X: Glutaminolysis and transferrin regulate ferroptosis. Mol
Cell. 59:298–308. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yanatori I, Richardson DR, Imada K and
Kishi F: Iron export through the transporter ferroportin 1 is
modulated by the iron chaperone PCBP2. J Biol Chem.
291:17303–17318. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lane DJ, Bae DH, Merlot AM, Sahni S and
Richardson DR: Duodenal cytochrome b (DCYTB) in iron metabolism: An
update on function and regulation. Nutrients. 7:2274–2296. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Qiu A, Jansen M, Sakaris A, Min SH,
Chattopadhyay S, Tsai E, Sandoval C, Zhao R, Akabas MH and Goldman
ID: Identification of an intestinal folate transporter and the
molecular basis for hereditary folate malabsorption. Cell.
127:917–928. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
White C, Yuan X, Schmidt PJ, Bresciani E,
Samuel TK, Campagna D, Hall C, Bishop K, Calicchio ML, Lapierre A,
et al: HRG1 is essential for heme transport from the phagolysosome
of macrophages during erythrophagocytosis. Cell Metab. 17:261–270.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Nishito Y and Kambe T: Absorption
mechanisms of iron, copper, and zinc: An overview. J Nutr Sci
Vitaminol (Tokyo). 64:1–7. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Cao JY and Dixon SJ: Mechanisms of
ferroptosis. Cell Mol Life Sci. 73:2195–2209. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhao J, Wang Y, Tao L and Chen L: Iron
transporters and ferroptosis in malignant brain tumors. Front
Oncol. 12:8618342022. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Hu C, Tao L, Cao X and Chen L: The solute
carrier transporters and the brain: Physiological and
pharmacological implications. Asian J Pharm Sci. 15:131–144. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Rouault TA and Cooperman S: Brain iron
metabolism. Semin Pediatr Neurol. 13:142–148. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Engelhardt B and Sorokin L: The
blood-brain and the blood-cerebrospinal fluid barriers: Function
and dysfunction. Semin Immunopathol. 31:497–511. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Skjorringe T, Moller LB and Moos T:
Impairment of interrelated iron- and copper homeostatic mechanisms
in brain contributes to the pathogenesis of neurodegenerative
disorders. Front Pharmacol. 3:1692012. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Li GJ, Choi BS, Wang X, Liu J, Waalkes MP
and Zheng W: Molecular mechanism of distorted iron regulation in
the blood-CSF barrier and regional blood-brain barrier following in
vivo subchronic manganese exposure. Neurotoxicology. 27:737–744.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zheng W and Monnot AD: Regulation of brain
iron and copper homeostasis by brain barrier systems: Implication
in neurodegenerative diseases. Pharmacol Ther. 133:177–188. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Qian ZM and Ke Y: Brain iron transport.
Biol Rev Camb Philos Soc. 94:1672–1684. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Burdo JR, Menzies SL, Simpson IA, Garrick
LM, Garrick MD, Dolan KG, Haile DJ, Beard JL and Connor JR:
Distribution of divalent metal transporter 1 and metal transport
protein 1 in the normal and Belgrade rat. J Neurosci Res.
66:1198–1207. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Carlson ES, Tkac I, Magid R, O'Connor MB,
Andrews NC, Schallert T, Gunshin H, Georgieff MK and Petryk A: Iron
is essential for neuron development and memory function in mouse
hippocampus. J Nutr. 139:672–679. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Wong BX, Tsatsanis A, Lim LQ, Adlard PA,
Bush AI and Duce JA: β-Amyloid precursor protein does not possess
ferroxidase activity but does stabilize the cell surface ferrous
iron exporter ferroportin. PLoS One. 9:e1141742014. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Miyajima H: Aceruloplasminemia.
Neuropathology. 35:83–90. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Gaasch JA, Lockman PR, Geldenhuys WJ,
Allen DD and Van der Schyf CJ: Brain iron toxicity: Differential
responses of astrocytes, neurons, and endothelial cells. Neurochem
Res. 32:1196–1208. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Huang R, Dong R, Wang N, He Y, Zhu P, Wang
C, Lan B, Gao Y and Sun L: Adaptive changes allow targeting of
ferroptosis for glioma treatment. Cell Mol Neurobiol. 42:2055–2074.
2022. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Du H, Ren X, Bai J, Yang W, Gao Y and Yan
S: Research progress of ferroptosis in adiposity-based chronic
disease (ABCD). Oxid Med Cell Longev. 2022:10526992022. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Shintoku R, Takigawa Y, Yamada K, Kubota
C, Yoshimoto Y, Takeuchi T, Koshiishi I and Torii S:
Lipoxygenase-mediated generation of lipid peroxides enhances
ferroptosis induced by erastin and RSL3. Cancer Sci. 108:2187–2194.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Yang WS, Kim KJ, Gaschler MM, Patel M,
Shchepinov MS and Stockwell BR: Peroxidation of polyunsaturated
fatty acids by lipoxygenases drives ferroptosis. Proc Natl Acad Sci
USA. 113:E4966–E4975. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Luo Y, Tian G, Fang X, Bai S, Yuan G and
Pan Y: Ferroptosis and its potential role in glioma: From molecular
mechanisms to therapeutic opportunities. Antioxidants (Basel).
11:21232022. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Zhou Y, Fang C, Xu H, Yuan L, Liu Y, Wang
X, Zhang A, Shao A and Zhou D: Ferroptosis in glioma treatment:
Current situation, prospects and drug applications. Front Oncol.
12:9898962022. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Yuan H, Li X, Zhang X, Kang R and Tang D:
Identification of ACSL4 as a biomarker and contributor of
ferroptosis. Biochem Biophys Res Commun. 478:1338–1343. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Chu B, Kon N, Chen D, Li T, Liu T, Jiang
L, Song S, Tavana O and Gu W: ALOX12 is required for p53-mediated
tumour suppression through a distinct ferroptosis pathway. Nat Cell
Biol. 21:579–591. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Tuo QZ, Lei P, Jackman KA, Li XL, Xiong H,
Li XL, Liuyang ZY, Roisman L, Zhang ST, Ayton S, et al:
Tau-mediated iron export prevents ferroptotic damage after ischemic
stroke. Mol Psychiatry. 22:1520–1530. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Shin D, Lee J, You JH, Kim D and Roh JL:
Dihydrolipoamide dehydrogenase regulates cystine
deprivation-induced ferroptosis in head and neck cancer. Redox
Biol. 30:1014182020. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Detivaud L, Island ML, Jouanolle AM,
Ropert M, Bardou-Jacquet E, Le Lan C, Mosser A, Leroyer P, Deugnier
Y, David V, et al: Ferroportin diseases: Functional studies, a link
between genetic and clinical phenotype. Hum Mutat. 34:1529–1536.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Luo M, Wu L, Zhang K, Wang H, Zhang T,
Gutierrez L, O'Connell D, Zhang P, Li Y, Gao T, et al: miR-137
regulates ferroptosis by targeting glutamine transporter SLC1A5 in
melanoma. Cell Death Differ. 25:1457–1472. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Dai C, Chen X, Li J, Comish P, Kang R and
Tang D: Transcription factors in ferroptotic cell death. Cancer
Gene Ther. 27:645–656. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Anandhan A, Dodson M, Schmidlin CJ, Liu P
and Zhang DD: Breakdown of an ironclad defense system: The critical
role of NRF2 in mediating ferroptosis. Cell Chem Biol. 27:436–447.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Liu XB, Yang F and Haile DJ: Functional
consequences of ferroportin 1 mutations. Blood Cells Mol Dis.
35:33–46. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Taniguchi R, Kato HE, Font J, Deshpande
CN, Wada M, Ito K, Ishitani R, Jormakka M and Nureki O: Outward-
and inward-facing structures of a putative bacterial
transition-metal transporter with homology to ferroportin. Nat
Commun. 6:85452015. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Galaris D, Barbouti A and Pantopoulos K:
Iron homeostasis and oxidative stress: An intimate relationship.
Biochim Biophys Acta Mol Cell Res. 1866:1185352019. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Pardridge WM: Drug transport across the
blood-brain barrier. J Cereb Blood Flow Metab. 32:1959–1972. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Wang H, An P, Xie E, Wu Q, Fang X, Gao H,
Zhang Z, Li Y, Wang X, Zhang J, et al: Characterization of
ferroptosis in murine models of hemochromatosis. Hepatology.
66:449–465. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Badgley MA, Kremer DM, Maurer HC,
DelGiorno KE, Lee HJ, Purohit V, Sagalovskiy IR, Ma A, Kapilian J,
Firl CEM, et al: Cysteine depletion induces pancreatic tumor
ferroptosis in mice. Science. 368:85–89. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Ingold I, Berndt C, Schmitt S, Doll S,
Poschmann G, Buday K, Roveri A, Peng X, Porto Freitas F, Seibt T,
et al: Selenium utilization by GPX4 is required to prevent
hydroperoxide-induced ferroptosis. Cell. 172:409–422. e212018.
View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Alim I, Caulfield JT, Chen Y, Swarup V,
Geschwind DH, Ivanova E, Seravalli J, Ai Y, Sansing LH, Ste Marie
EJ, et al: Selenium drives a transcriptional adaptive program to
block ferroptosis and treat stroke. Cell. 177:1262–1279. e252019.
View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Doll S, Freitas FP, Shah R, Aldrovandi M,
da Silva MC, Ingold I, Goya Grocin A, Xavier da Silva TN, Panzilius
E, Scheel CH, et al: FSP1 is a glutathione-independent ferroptosis
suppressor. Nature. 575:6936–698. 2019. View Article : Google Scholar
|
|
71
|
Bersuker K, Hendricks JM, Li Z, Magtanong
L, Ford B, Tang PH, Roberts MA, Tong B, Maimone TJ, Zoncu R, et al:
The CoQ oxidoreductase FSP1 acts parallel to GPX4 to inhibit
ferroptosis. Nature. 575:688–692. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Wang Z, Ding Y, Wang X, Lu S, Wang C, He
C, Wang L, Piao M, Chi G, Luo Y and Ge P: Pseudolaric acid B
triggers ferroptosis in glioma cells via activation of Nox4 and
inhibition of xCT. Cancer Lett. 428:21–33. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Wu Y, Zhao Y, Yang HZ, Wang YJ and Chen Y:
HMGB1 regulates ferroptosis through Nrf2 pathway in mesangial cells
in response to high glucose. Biosci Rep. 41:BSR202029242021.
View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Fan Z, Wirth AK, Chen D, Wruck CJ, Rauh M,
Buchfelder M and Savaskan N: Nrf2-Keap1 pathway promotes cell
proliferation and diminishes ferroptosis. Oncogenesis. 6:e3712017.
View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Roh JL, Kim EH, Jang H and Shin D: Nrf2
inhibition reverses the resistance of cisplatin-resistant head and
neck cancer cells to artesunate-induced ferroptosis. Redox Biol.
11:254–262. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Sun X, Ou Z, Chen R, Niu X, Chen D, Kang R
and Tang D: Activation of the p62-Keap1-NRF2 pathway protects
against ferroptosis in hepatocellular carcinoma cells. Hepatology.
63:173–184. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Chen D, Fan Z, Rauh M, Buchfelder M,
Eyupoglu IY and Savaskan N: ATF4 promotes angiogenesis and neuronal
cell death and confers ferroptosis in a xCT-dependent manner.
Oncogene. 36:5593–5608. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Zhao X, Zhou M, Yang Y and Luo M: The
ubiquitin hydrolase OTUB1 promotes glioma cell stemness via
suppressing ferroptosis through stabilizing SLC7A11 protein.
Bioengineered. 12:12636–12645. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Yi R, Wang H, Deng C, Wang X, Yao L, Niu
W, Fei M and Zhaba W: Dihydroartemisinin initiates ferroptosis in
glioblastoma through GPX4 inhibition. Biosci Rep.
40:BSR201933142020. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Song Q, Peng S, Sun Z, Heng X and Zhu X:
Temozolomide drives ferroptosis via a DMT1-Dependent pathway in
glioblastoma cells. Yonsei Med J. 62:843–849. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Zhang Y, Tan H, Daniels JD, Zandkarimi F,
Liu H, Brown LM, Uchida K, O'Connor OA and Stockwell BR: Imidazole
ketone erastin induces ferroptosis and slows tumor growth in a
mouse lymphoma model. Cell Chem Biol. 26:623–633. e92019.
View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Eaton JK, Furst L, Ruberto RA, Moosmayer
D, Hilpmann A, Ryan MJ, Zimmermann K, Cai LL, Niehues M, Badock V,
et al: Selective covalent targeting of GPX4 using masked
nitrile-oxide electrophiles. Nat Chem Biol. 16:497–506. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Sukseree S, Schwarze UY, Gruber R, Gruber
F, Quiles Del Rey M, Mancias JD, Bartlett JD, Tschachler E and
Eckhart L: ATG7 is essential for secretion of iron from ameloblasts
and normal growth of murine incisors during aging. Autophagy.
16:1851–1857. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Zhang Y, Kong Y, Ma Y, Ni S, Wikerholmen
T, Xi K, Zhao F, Zhao Z, Wang J, Huang B, et al: Loss of COPZ1
induces NCOA4 mediated autophagy and ferroptosis in glioblastoma
cell lines. Oncogene. 40:1425–1439. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Santini ZI, Jose PE, York Cornwell E,
Koyanagi A, Nielsen L, Hinrichsen C, Meilstrup C, Madsen KR and
Koushede V: Social disconnectedness, perceived isolation, and
symptoms of depression and anxiety among older Americans (NSHAP): A
longitudinal mediation analysis. Lancet Public Health. 5:e62–e70.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Forero DA, Guio-Vega GP and
Gonzalez-Giraldo Y: A comprehensive regional analysis of
genome-wide expression profiles for major depressive disorder. J
Affect Disord. 218:86–92. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Tang W, Lu Y and Xu J: Post-traumatic
stress disorder, anxiety and depression symptoms among adolescent
earthquake victims: Comorbidity and associated sleep-disturbing
factors. Soc Psychiatry Psychiatr Epidemiol. 53:1241–1251. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Gialluisi A, Bonaccio M, Di Castelnuovo A,
Costanzo S, De Curtis A, Sarchiapone M, Cerletti C, Donati MB, de
Gaetano G and Iacoviello L; Moli-Sani Study Investigators, :
Lifestyle and biological factors influence the relationship between
mental health and low-grade inflammation. Brain Behav Immun.
85:4–13. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Treudler R, Zeynalova S, Riedel-Heller SG,
Zuelke AE, Roehr S, Hinz A, Glaesmer H, Kage P, Loeffler M and
Simon JC: Depression, anxiety and quality of life in subjects with
atopic eczema in a population-based cross-sectional study in
Germany. J Eur Acad Dermatol Venereol. 34:810–816. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Singer S, Roick J, Danker H, Kortmann RD,
Papsdorf K, Taubenheim S, Renovanz M, Jähne K and Meixensberger J:
Psychiatric co-morbidity, distress, and use of psycho-social
services in adult glioma patients-a prospective study. Acta
Neurochir (Wien). 160:1187–1194. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Louis DN, Perry A, Reifenberger G, von
Deimling A, Figarella-Branger D, Cavenee WK, Ohgaki H, Wiestler OD,
Kleihues P and Ellison DW: The 2016 World health organization
classification of tumors of the central nervous system: A summary.
Acta Neuropathol. 131:803–820. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Ostrom QT, Gittleman H, Fulop J, Liu M,
Blanda R, Kromer C, Wolinsky Y, Kruchko C and Barnholtz-Sloan JS:
CBTRUS statistical report: Primary brain and central nervous system
tumors diagnosed in the United States in 2008–2012. Neuro Oncol. 17
(Suppl 4):iv1–iv62. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Hao A, Huang J and Xu X: Anxiety and
depression in glioma patients: Prevalence, risk factors, and their
correlation with survival. Ir J Med Sci. 190:1155–1164. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Young K and Singh G: Biological mechanisms
of cancer-induced depression. Front Psychiatry. 9:2992018.
View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Satin JR, Linden W and Phillips MJ:
Depression as a predictor of disease progression and mortality in
cancer patients: A meta-analysis. Cancer. 115:5349–5361. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Sawangjit A, Oyanedel CN, Niethard N,
Salazar C, Born J and Inostroza M: The hippocampus is crucial for
forming non-hippocampal long-term memory during sleep. Nature.
564:109–113. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Dang R, Wang M, Li X, Wang H, Liu L, Wu Q,
Zhao J, Ji P, Zhong L, Licinio J and Xie P: Edaravone ameliorates
depressive and anxiety-like behaviors via Sirt1/Nrf2/HO-1/Gpx4
pathway. J Neuroinflammation. 19:412022. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Jiao H, Yang H, Yan Z and Chen J, Xu M,
Jiang Y, Liu Y, Xue Z, Ma Q, Li X and Chen J: Traditional Chinese
formula xiaoyaosan alleviates depressive-like behavior in CUMS Mice
by Regulating PEBP1-GPX4-Mediated Ferroptosis in the Hippocampus.
Neuropsychiatr Dis Treat. 17:1001–1019. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Erratum to: path-03. Ferroptosis-related
long non-coding rna signatures predict prognosis in patients with
glioma. Neuro Oncol. 24:20102022. View Article : Google Scholar : PubMed/NCBI
|