Introduction
In recent years, the clinical use of surgery,
radiation therapy and chemotherapy has reduced the recurrence rates
of cancer. However, cellular resistance to chemotherapeutic drugs
remains a major obstacle in the successful treatment of cancer
(1,2). The efficacy of chemotherapy is limited
due to acquired resistance from previous treatment. Consequently,
research strategies to circumvent such resistance in cancer cells
have become a current focus for the development of novel
combination chemotherapy. Both intrinsic and acquired drug
resistance can produce multiple changes in various cellular
pathways, leading to a decrease in the cytotoxicity, and, thus, a
reduction in the efficacy of antineoplastic drugs (3). Therefore, cancer patients that receive
chemotherapy can become increasingly insensitive to
chemotherapeutic drugs.
One of the primary cellular mechanisms that produces
resistance to antineoplastic therapy involves the efflux of drugs
from the cancer cells by specific transmembrane transporters or
pumps (4). These transporter
proteins originate from the superfamily of ATP-binding cassette
(ABC) transporters that share common structural and functional
properties (5). Previous studies
have shown that the majority of the members of the C subfamily of
ABC transporters are multidrug resistance proteins (MRPs/ABCCs),
which are characterized by cross-resistance to several structurally
unrelated drugs (2,4,6,7).
A number of studies suggest that cancer cells that
express the ABCC subfamily transporter multidrug resistance protein
7 (MRP7/ABCC10) can develop resistance to various chemotherapeutic
drugs. For example, human salivary gland adenocarcinoma (SGA) cells
that overexpress MRP7 mRNA and the MRP7 protein display
significant resistance to vincristine (8). MRP7 expression has also been
immunohistochemically identified in tumor-bearing mice xenografted
with human SGA following treatment with vincristine (8). In addition, E217βG,
a competitive inhibitor of MRP7 transport, significantly decreased
docetaxel accumulation in human SGA cells (8).
The MRP7-overexpressing cells confer resistance to
several anticancer drugs including paclitaxel, vincristine and
vinblastine (9). Recent studies
also reported that MRP7-overexpressing cells confer resistance to
nucleoside analogues and epothilone B (10). Furthermore, our laboratory revealed
that cepharanthine, a biscoclaurine-derived alkaloid, reversed
MRP7-mediated paclitaxel resistance (11).
An important discovery about tyrosine kinase
inhibitors (TKIs) was that certain ‘small molecule’ drugs could
inhibit TK activity by competing with ATP for binding to the
intracellular catalytic domain of receptor TKs, which produced
inhibition of various downstream signaling cascades by
autophosphorylation (12). Notably,
imatinib, nilotinib and dasatinib are inhibitors of the TK
breakpoint cluster region-Abelson (BCR-Abl) and KIT, a class
III receptor TK (13–17). The BCR-Abl gene is associated
with a dysregulation of TK function, subsequently leading to a
malignant transformation in chronic myelogenous leukemia (CML)
(18,19). The recognition of the BCR-Abl
gene and its corresponding protein has led to the development of
small-molecule drugs designed to block the activation of
BCR-Abl TK through competitive binding at the ATP-binding
site (18).
In recent years, several experiments determined that
TKIs can reverse the resistance of cancer cells to antineoplastic
drugs through multiple mechanisms. We and others have reported that
some of the TKIs are potent modulators of ABC transporters,
including P-glycoprotein (P-gp) and breast cancer resistance
protein (BCRP/ABCG2) (20,21). Results from our laboratory suggested
that nilotinib significantly reverses P-gp- and BCRP-mediated MDR
(22). Our further study found that
imatinib and nilotinib can reverse MDR in cancer cells by
inhibiting the efflux activity of the MRP7/ABCC10 (23). In addition, we also reported that
lapatinib and erlotinib are potent reversal agents for
MRP7/ABCC10-mediated MDR (24).
Tandutinib (MLN518/CT53518) is a novel
quinazoline-based inhibitor of FMS-like tyrosine kinase 3 (FLT3, a
transmembrane receptor in the tyrosine kinase family),
platelet-derived growth factor receptor and KIT (25). In the present study, we evaluated
the possible interactions of tandutinib with MRP7/ABCC10, with the
aim to identify if tandutinib can reverse MRP7/ABCC10-mediated drug
resistance. Consequently, it is possible that tandutinib, in
combination with other antineoplastic drugs, may be useful in the
treatment of cancer that may express MDR proteins, including the
ABC transporters.
Materials and methods
Materials
Dulbecco's modified Eagle's medium (DMEM), bovine
serum and penicillin/streptomycin were purchased from HyClone
(Logan, UT, USA). Tandutinib was a product of Selleck Chemicals LLC
(Houston, TX, USA). Paclitaxel, fetal bovine serum (FBS), dimethyl
sulfoxide (DMSO) and
1-(4,5-dimethylthiazol-2-yl)-3,5-diphenylformazan (MTT), the
polyclonal goat antibody against MRP7 (C-19), glyceraldehyde
3-phosphate dehydrogenase (GAPDH), the secondary horseradish
peroxidase-labeled anti-goat and anti-mouse IgG were purchased from
Sigma-Aldrich Chemical Co. (St. Louis, MO, USA).
[3H]-paclitaxel (45 mCi/mmol) was purchased from Moravek
Biochemicals (Brea, CA, USA). Other routine laboratory reagents
were obtained from commercial sources of analytical grade.
Cell lines and cell culture
HEK293 cells and the MRP7 cDNA were generously
provided by Dr Gary Kruh (University of Illinois at Chicago). The
HEK293-MRP7-transfected cells and empty vector transfected
HEK293-pcDNA3.1 cells were established from HEK293 cells through
electroporation (26). Both cell
lines were grown as adherent monolayers in flasks with DMEM
supplemented with 10% FBS, 2 mM glutamine, 100 U/ml penicillin, and
100 mg/ml streptomycin under standard cell culturing conditions in
a humidified incubator containing 5% CO2 at 37°C.
MTT cytotoxicity assay
Prior to the antineoplastic drug sensitivity
analysis, we performed the MTT cytotoxicity assay of tandutinib on
HEK293-pcDNA3.1 cells and HEK293-MRP7-transfected cells, and the
procedure was the same as the following.
Drug sensitivity was analyzed using an MTT
colorimetric assay (20). Empty
vector-transfected HEK293-pcDNA3.1 cells and
HEK293-MRP7-transfected cells were seeded in 96-well plates in
triplicate at 5,000 cells/well. Following incubation in DMEM
supplemented with 10% FBS at 37°C for 24 h, various concentrations
of antineoplastic drugs were added and incubated with the cells
continuously for 72 h. For the combination group, a potential
inhibitor was added 1 h prior to the addition of an anticancer
drug.
Following drug incubation of 72 h, 20 μl MTT (4
mg/ml) was added to each well and the plate was further incubated
for 4 h, allowing viable cells to develop from the yellow-colored
MTT into dark-blue formazan crystals. Subsequently, the medium was
gently removed without agitating the adhesive monolayer of cells,
and 100 μl of DMSO was added into each well to dissolve the
formazan crystals. The plates were well shaken for 5 min, and an
Opsys microplate reader read the absorbance at 570 nm (Dynex
Technologies Inc, Chantilly, VA, USA). The degree of resistance was
calculated by dividing the IC50 for the MDR cells by
that of the parental cells, whereas the degree of MDR reversal was
calculated by dividing the IC50 of the cells with the
anticancer drug in the absence of inhibitor by that obtained in the
presence of the inhibitor. The concentrations required to inhibit
growth by 50% of the control cells were calculated from survival
curves using a modified Bliss method (27).
The antineoplastic drugs used in this study included
paclitaxel, vincristine and cisplatin at varying concentrations up
to a final concentration of 3, 3 and 100 μM, respectively.
Tandutinib was used at non-toxic concentrations of 5, 10 and 20 μM
and lapatinib at 3 μM to screen against paclitaxel. We subsequently
selected their concentrations to determine whether their reversal
effects were concentration-dependent to paclitaxel, vincristine and
cisplatin.
Preparation of cell lysates
The cell lines were cultured in DMEM containing 10%
FBS at 37°C in the presence of 5% CO2. Confluent
monolayer cells in T-25 flask were harvested and rinsed twice with
cold PBS. The cell extracts were prepared using the
Radioimmunoprecipitation assay buffer (1X PBS, 1% Nonidet P-40,
0.5% sodium deoxycholate, 0.1% SDS, 100 mM p-APMSF, 10 mM leupeptin
and 10 mM aprotinin) for 30 min on ice with occasional rocking
followed by centrifugation at 12,000 rpm at 4°C for 15 min. The
supernatant containing total cell lysates was collected and stored
at −80°C until use.
Immunoblotting
Equal amounts of total cell lysates (40 μg) were
resolved by 4–12% sodium dodecyl sulfate polyacrylamide gel
electrophoresis (SDS-PAGE) and electrophoretically transferred onto
nitrocellulose membranes (21). The
cell lysates were denatured in a 100°C water beaker for 5 min
before loading onto the 4–12% SDS-PAGE. The gel was run in the SDS
electrophoresis buffer (25 mM Tris base, 0.192 M glycine, 1% SDS)
at 170 V for 2 h. The transfer was performed in a transfer buffer
(25 mM Tris base, 0.192 M glycine, pH 8.3) at 80 V for 2 h. The
nitrocellulose membrane was then immersed in 5% skim milk to block
non-specific binding for 1 h at room temperature. The membrane was
then immunoblotted overnight with primary antibodies (polyclonal
MRP7 to GAPDH at 1:200 and polyclonal MRP7 at 1:400) at 4°C. The
following day, the membrane was washed three times with TBST buffer
(0.3% Tris, 0.8% NaCl, 0.02% KCl, 0.05% Tween-20) followed by a 2-h
incubation with secondary antibody against GAPDH (ab9483) at
1:2,000. The protein-antibody complex was measured using an
enhanced chemiluminescence detection system (Amersham Biosciences,
Piscataway, NJ, USA). The membrane was then exposed to the film for
development. The conventionally used loading control GAPDH was used
to detect equal loading in each lane in the samples prepared from
cell lysates.
Paclitaxel accumulation
Cells in 24-well plates were preincubated with or
without tandutinib or lapatinib for 1 h at 37°C, then incubated
with 0.1 μM [3H]-paclitaxel for 2 h in the presence or
absence of the inhibitors (tandutinib or lapatinib) at 37°C. After
washing 3 times with ice-cold PBS, the cells were trypsinized and
lysed in 10 mM lysis buffer (pH 7.4, containing 1% Triton X-100 and
0.2% SDS). Each sample was placed in scintillation fluid and
radioactivity was measured in a Packard TRI-Carb 1900CA liquid
scintillation analyzer from Packard Instrument Company, Inc.
(Downers Grove, IL, USA).
Paclitaxel efflux
The HEK293-pcDNA3.1 cells and HEK-transfected cells
were seeded in two T-75 flasks and incubated with DMEM supplemented
with 10% FBS at 37°C. After the cells were grown to 60–80%
confluency, each inhibitor (tandutinib or lapatinib) was added to
separate flasks and the cells were incubated for 1 h. The cells
were then trypsinized and two aliquots (730,000 cells) from each
cell line were suspended in the medium. Subsequently, cells were
suspended in the medium containing [3H]-paclitaxel at a
concentration of 0.1 μM with or without inhibitor for 1 h at 37°C.
The incubation medium was replaced by the medium containing only an
inhibitor without [3H]-paclitaxel. Aliquots (233,000
cells) were collected at various time points (0, 30, 60 and 120
min). The cells were then washed with ice-cold PBS and each sample
was placed in scintillation fluid to measure the radioactivity in a
Packard Tri-Carb 1900CA liquid scintillation counter from Packard
Instrument Inc.
Statistical analysis
All experiments were repeated at least three times
and the differences were determined by two-tailed Student's t-test.
When statistical differences between more than 2 groups were
analyzed, one-way ANOVA followed by Tukey's multiple comparison
test were performed, as indicated. Results are presented as the
means ± standard deviations (SD). The statistical significance of
differences was determined at P<0.05.
Results
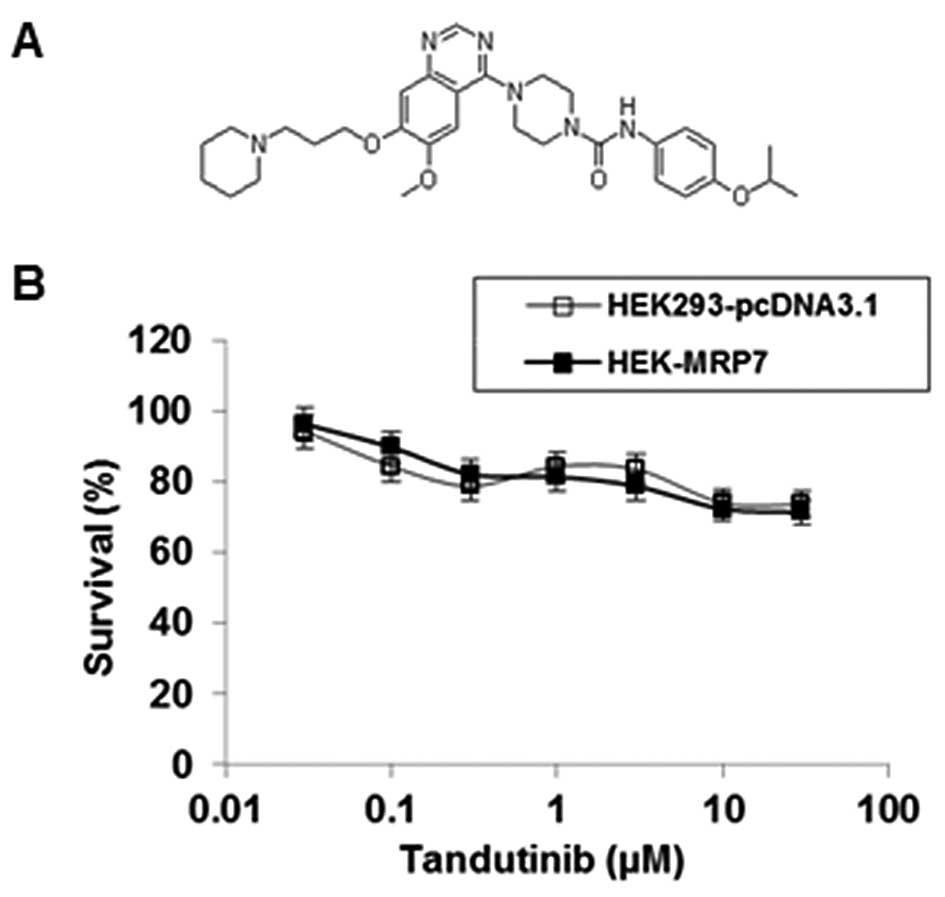
Sensitivity of tandutinib on
HEK293-pcDNA3.1 and HEK-MRP7-transfected cells
To investigate the effect of tandutinib on the MRP7
transporter, we first examined the sensitivity of HEK293-pcDNA3.1
and HEK-MRP7 cells to tandutinib. As shown in Fig. 1B, the IC50 values of
tandutinib on both HEK293-pcDNA3.1 and HEK-MRP7 cells were >30
μM. Notably, the results of the MTT assay showed that tandutinib
did not significantly inhibit the cell growth of these two cell
lines at concentrations up to 10 μM (Fig. 1B).
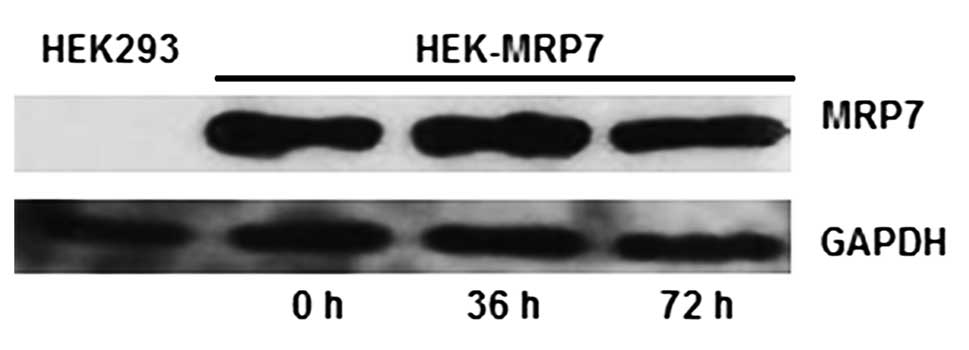
Effect of tandutinib on the expression of
MRP7 in HEK-MRP7 cells
Immunoblot analysis was performed to detect the
expression levels of MRP7 protein in the aforementioned lines. MRP7
protein (MW 171 kDa) was expressed in HEK-MRP7 cells, but not in
HEK293-pcDNA3.1 cells (Fig. 2).
GAPDH, with a molecular weight of 38 kDa, was detected in the
HEK293-pcDNA3.1 or HEK-MRP7 cell lines (Fig. 2).
To evaluate the effect of tandutinib on the
expression of MRP7, the HEK-MRP7 cells were incubated with 10 μM of
tandutinib for 36 and 72 h. The incubation of HEK-MRP7 cells with
tandutinib did not significantly alter the expression of the
protein levels of MRP7 at different time points (Fig. 2), which is similar to our results
for lapatinib (24). This finding
suggests that the reversal effect of tandutinib on MRP7-mediated
MDR is not due to the regulation of MRP7 expression.
Analysis of the drug sensitivity of
HEK293-pcDNA3.1 and MRP7-transfected HEK293 cells
To determine the drug resistance profile of MRP7,
the sensitivity of HEK-MRP7-transfected cells to specific
antineoplastic drugs was compared to that of the empty
vector-transfected cells, HEK293-pcDNA3.1. The HEK-MRP7 cells
exhibited a significantly higher level of resistance to paclitaxel
and vincristine (15.94- and 6.25-fold resistance compared to the
control cells, respectively) (Table
I). These results indicated that the HEK-MRP7 cell line was
able to confer resistance to various antineoplastic drugs, which is
consistent with our previous reports (11,23).
 | Table IEffect of tandutinib on reversing
MRP7-mediated resistance to paclitaxel, vincristine and
cisplatin. |
Table I
Effect of tandutinib on reversing
MRP7-mediated resistance to paclitaxel, vincristine and
cisplatin.
|
HEK293-pcDNA3.1 | HEK-MRP7 |
|---|
|
|
|
|---|
| Compounds | IC50 ±
SD (nM) (RF) | DMFc | IC50 ±
SD (nM) (RF) | DMFd |
|---|
| Paclitaxel | 3.52±0.32
(1.0) | | 56.04±4.09
(15.94) | |
| +Tandutinib 5
μM | 3.37±0.32
(0.96) | 1.04 | 11.72±0.74
(3.33) | 4.78d |
| +Tandutinib 10
μM | 3.42±0.35
(0.97) | 1.03 | 3.30±0.28
(0.94) | 17.0e |
| +Lapatinib 3
μM | 3.27±0.19
(0.93) | 1.08 | 4.29±0.36
(1.22) | 13.07e |
| Vincristine | 2.5±0.24
(1.0)b | | 15.6±1.48
(6.25) | |
| +Tandutinib 5
μM | 2.26±0.16
(0.91) | 1.10 | 3.22±0.31
(1.29) | 4.85d |
| +Tandutinib 10
μM | 1.76±0.15
(0.7) | 1.42 | 2.01±0.21
(0.81) | 7.75d |
| +Lapatinib 3
μM | 2.02±0.19
(0.81) | 1.23 | 1.91±0.19
(0.77) | 8.17d |
| Cisplatin | 3428±99 (1.0) | | 3724±615
(1.09) | |
| +Tandutinib 10
μM | 3515±407
(1.03) | 0.98 | 4335±615
(1.26) | 0.86 |
| +Lapatinib 3
μM | 3311±367
(0.97) | 1.04 | 3583±430
(1.05) | 1.04 |
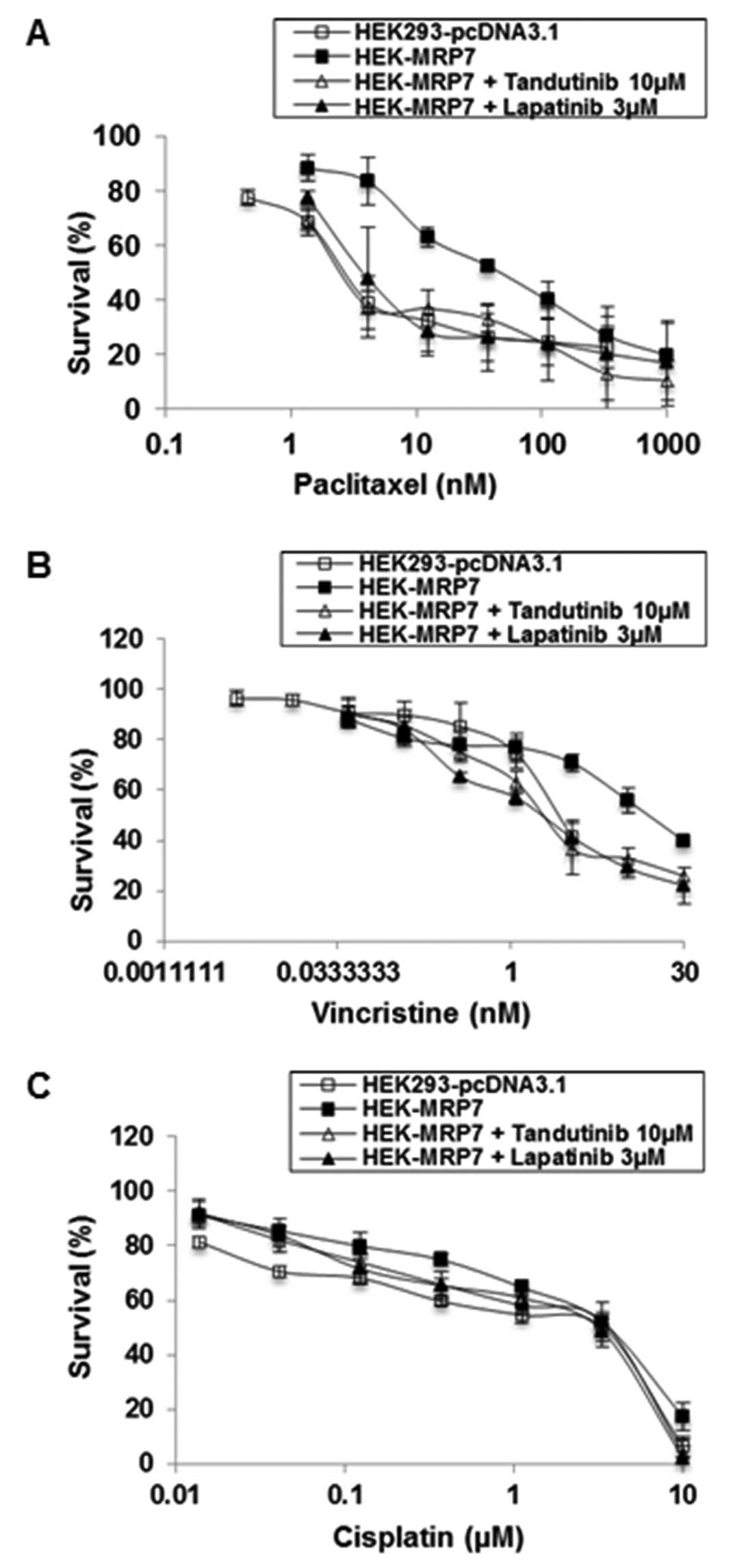
Effect of tandutinib on the sensitivity
of MRP7-transfected HEK293 cells to anticancer drugs
The preincubation of cells with tandutinib at 10 μM
or lapatinib as a positive control, at 3 μM, significantly reversed
the resistance of HEK-MRP7 cells to paclitaxel (Table I, Fig.
3A and B). Tandutinib and lapatinib produced a 17.0- and
13.07-fold reversal, respectively, of the resistance to paclitaxel.
The IC50 of paclitaxel in HEK-MRP7 cells co-cultured
with 10 μM of tandutinib was significantly decreased from
56.04±4.09 to 3.3±0.28 nM, which is comparable to that of
paclitaxel in HEK293-pcDNA3.1 cells (3.52±0.32 nM). These results
suggested that the resistance to paclitaxel was completely reversed
when tandutinib was co-incubated with paclitaxel in HEK-MRP7 cells.
When HEK293-pcDNA3.1 was co-incubated with tandutinib and
paclitaxel, tandutinib, at 5 and 10 μM concentrations, had no
significant effect on the sensitivity to paclitaxel. Compared with
this sensitivity in HEK293-pcDNA3.1 cells, the sensitivity
determined for the MRP7-transfected cells was much greater
(Table I, Fig. 3A).
In addition to paclitaxel, we also examined the
effect of tandutinib to sensitize cells to another anticancer drug,
vincristine. Similar to the findings with paclitaxel, tandutinib (5
and 10 μM) and lapatinib (3 μM) significantly reversed
MRP7-mediated vincristine resistance (4.85-, 7.75-fold for
tandutinib; and 8.17-fold for lapatinib) in a
concentration-dependent manner (Table
I and Fig. 3B). We also
examined the response of MRP7-transfected cells to a non MRP7
substrate anticancer drug, cisplatin. Our results indicated that
tandutinib (5 and 10 μM) did not significantly sensitize the
response of HEK293-pcDNA3.1 and HEK-MRP7 cells to cisplatin
(Table I and Fig. 3C). This indicates that the response
to these drugs is specific for MRP7, as cisplatin is not a
substrate for MRP7 and thus would not mediate cisplatin efflux.
In conclusion, tandutinib, similar to lapatinib,
significantly reversed MRP7-mediated resistance to paclitaxel and
vincristine, but not cisplatin (Table
I, Fig. 3).
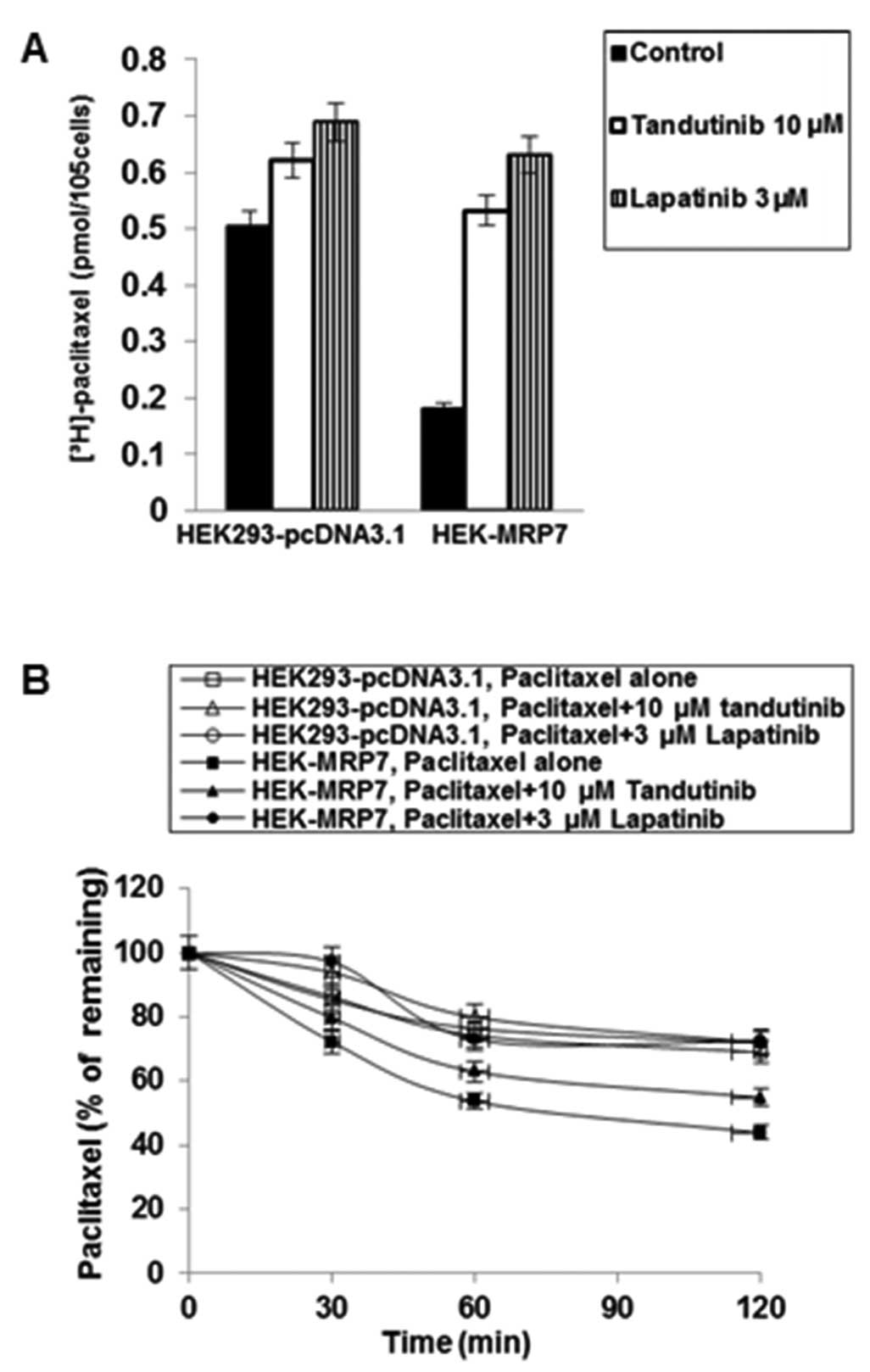
Effects of tandutinib on the
intracellular accumulation of [3H]-paclitaxel
To investigate the potential mechanism by which
tandutinib sensitizes MRP7-transfected cells to chemotherapeutic
drugs, we examined the effect of tandutinib on the accumulation of
[3H]-paclitaxel. Intracellular
[3H]-paclitaxel was measured in MRP7-transfected and
empty vector-transfected cells and the results are shown in
Fig. 4A. After a 2-h incubation,
the intracellular level of [3H]-paclitaxel in
MRP7-transfected cells was significantly lower than that in the
parental HEK293-pcDNA3.1 cells. Tandutinib at 10 μM significantly
increased the intracellular level of [3H]-paclitaxel
similar to the effect of lapatinib at 3 μM in HEK-MRP7 cells.
Effects of tandutinib on the efflux of
[3H]-paclitaxel
We previously determined that lapatinib and
erlotinib could also reverse MRP7-mediated drug resistance and
found that their effects are due to their interaction with the MRP7
protein (24). In the present
study, using lapatinib as a comparison agent, we sought to further
determine the mechanism by which tandutinib increases intracellular
accumulation of [3H]-paclitaxel in HEK-MRP7 cells.
It is possible that the increase of intracellular
paclitaxel produced by tandutinib is due to: i) a decrease in the
efflux of paclitaxel and/or, ii) an increase in the uptake of
paclitaxel. HEK-MRP7 cells and HEK293-pcDNA3.1 cells were incubated
with paclitaxel and a time course for intracellular paclitaxel
remaining was determined (Fig. 4B).
HEK-MRP7 cells released a significantly higher percentage of
accumulated paclitaxel than HEK293-pcDNA3.1 cells, and the amount
of paclitaxel that was effluxed increased with time. When the cells
were incubated with tandutinib at 10 μM or lapatinib at 3 μM, they
significantly blocked the intracellular [3H]-paclitaxel
efflux at different time points (0, 30, 60 and 120 min) from
HEK-MRP7 cells, but not from HEK293-pcDNA3.1 cells. The
accumulation of [3H]-paclitaxel at the 0 min time point
of efflux was set as 100%, at 30, 60 and 120 min of drug efflux
time points, the percentages of the accumulated
[3H]-paclitaxel that remained in HEK-MRP7 cells in the
absence of tandutinib or lapatinib were 72.18, 53.71 and 43.86%,
respectively. When HEK-MRP7 cells were incubated with tandutinib,
the percentages of intracellular paclitaxel remaining at 30, 60 and
120 min were increased to 79.66, 62.94 and 54.84%, respectively
(P<0.05 for the same time point comparison) (Fig. 4B). Lapatinib increased the
percentage of [3H]-paclitaxel accumulation at 30, 60 and
120 min to 97.08, 72.97 and 72.34%, respectively (P<0.05 for the
same time point comparison) (Fig.
4B). Lapatinib was slightly more potent than tandutinib, which
is consistent with the results in colorimetric growth assay and
[3H]-paclitaxel accumulation experiments.
Discussion
It is well established that MRP7, P-gp and MRP1 are
drug efflux pumps responsible for the transport of a variety of
antineoplastic drugs from the cells. Consequently, when these pumps
are present in the tumor cells concurrently, each of the pumps
contributes to the efflux of the drugs and decreases of
intracellular drug concentrations. This latter action ultimately
leads to lower drug levels that are no longer cytotoxic, leading to
the failure of cancer chemotherapy. For instance, nilotinib has
been identified as an inhibitor of P-gp and the BCRP efflux pumps
(22), and has also been identified
as a reversal agent for MRP7-mediated resistance to paclitaxel
(23). More recently, we found that
nilotinib was also able to revise ABC transporter-mediated MDR in
in vivo tumor xenograft mouse models (28). These findings suggested for the
first time that TKIs may be useful in treating cancer that has
become resistant to anticancer drugs as a result of ABC transporter
overexpression.
Tandutinib, known as a small-molecule inhibitor of
FLT3, is mainly used for acute myelogenous leukemia (AML) and is
currently in phase II clinical trials. Its phase I clinical results
with tandutinib in patients with AML or high-risk myelodysplastic
syndrome showed safety with appropriate pharmacokinetics and
pharmacodynamics (25). Previously,
Yang et al(29) demonstrated
that tandutinib was a substrate of P-gp and BCRP. P-gp and BCRP
played a role in oral absorption, systemic clearance and brain
penetration of tandutinib in the rodents. It has been reported that
cepharanthine and nilotinib significantly reversed P-gp-mediated
MDR in HEK-MRP7 cells (11) and
BCRP-overexpressing cell lines (22), respectively. Our study also found
that lapatinib and erlotinib can reverse MRP7-mediated MDR through
inhibition of the drug efflux function (24).
In the present study, we determined if tandutinib
has the ability to reverse MRP7-mediated drug resistance. We used
non-toxic concentrations of tandutinib (10 μM) and lapatinib (3
μM). The transfected HEK293-pcDNA3.1 and HEK-MRP7 cell lines used
in our experiments have been used in a previous study from our
laboratory (24). We used western
blot analysis to detect the expression of MRP7. The cell lines were
exposed to the same experimental conditions and procedures, and
were cultured with the same antineoplastic drugs for the same
incubation time.
We found that tandutinib (10 μM) significantly
sensitized MRP7-transfected HEK293 cells to paclitaxel as it
markedly decreased the IC50 of paclitaxel in
MRP7-transfected cells HEK-MRP7, compared to control cells
(Table I). In order to extend the
findings obtained with paclitaxel, we examined the effect of
specific TKIs on the response of MRP7-expressing cells to another
antineoplastic drug, vincristine, which is another substrate for
MRP7. Our results indicated that tandutinib (10 μM) completely
reversed MRP7-mediated vincristine resistance by a factor of
7.75-fold (Table I). Thus, similar
to lapatinib, tandutinib significantly attenuates MRP7-mediated
resistance to paclitaxel as well as to vincristine. As an
additional control, we examined the effect of tandutinib on the
response of MRP7-transfected cells to cisplatin, which is not a
substrate for MRP7. The results showed that tandutinib (10 μM) did
not significantly alter the response of cells to cisplatin
(Table I). In addition, our
previous study found that P-gp, MRP1 and BCRP were undetectable in
both control HEK293-pcDNA3.1 and transfected HEK-MRP7 cells through
western blot analysis (24). These
results indicated that the HEK-MRP7-transfected cell line
specifically expressed MRP7, but not P-gp, MRP1, or BCRP, and the
actions of tandutinib in reversing MRP7-mediated resistance are due
to a specific effect on the MRP7 pump.
Since the MTT assay results cannot be used as a
direct confirmation of MRP7-mediated drug transport, we determined
the effect of tandutinib on the accumulation and efflux of
[3H]-paclitaxel, a known chemotherapeutic substrate of
MRP7 transporter, in HEK293-pcDNA3.1 and HEK-MRP7 cells (10). In our experiments, tandutinib
significantly increased the intracellular concentration of
[3H]-paclitaxel, and decreased the intracellular
[3H]-paclitaxel efflux from the HEK-MRP7 cells but not
in the parental HEK293-pcDNA3.1 cells. The reversal effect of
tandutinib was similar to that of lapatinib. This suggests that
tandutinib modulates MRP7-mediated MDR by increasing intracellular
drug accumulation by inhibiting the drug efflux function of
MRP7.
In this study, we found that tandutinib
significantly decreased resistance to paclitaxel and vincristine in
MRP7-overexpressing cells (Table I,
Fig. 3) and did not significantly
inhibit or induce MRP7 expression (Fig.
2). Collectively, these findings tentatively suggest that
tandutinib is capable of reversing MRP7-mediated resistance by
inhibiting the function of MRP7.
Previous studies have demonstrated that the systemic
exposure achieved at the standard recommended dose of nilotinib is
similar to the concentration that reversed drug resistance in in
vitro cell models (30,31). Previous studies verified that
nilotinib is able to revise ABC transporter-mediated MDR both in
vitro and in vivo(22,23,28).
Thus, we expect it is also possible that tandutinib is a useful
chemosensitizing drug in the clinic for cancer patients if the
plasma tandutinib drug concentration can reach 5 μM. Further
studies in in vivo tumor xenograft studies are required to
evaluate the effects of specific FLT3 inhibitors such as tandutinib
on the resistance of cancer cells to antineoplastic drugs.
In conclusion, our findings show for the first time
that the small-molecule inhibitor of FLT3 tandutinib can
effectively reverse MRP7-mediated MDR. The mechanism of MDR
modulation by tandutinib is associated with an increase in
intracellular drug accumulation by inhibiting drug efflux from MDR
cells via MRP7. This suggests that tandutinib could be used to
augment the clinical response to conventional chemotherapeutic
agents that are substrates of MRP7. Therefore, tandutinib may be a
useful modifier of MRP7-mediated MDR in cancer patients.
Acknowledgements
The authors thank Dr Gary Kruh (University of
Illinois at Chicago) for providing HEK293 cells, MRP7 cDNA and also
for his support. We thank Mr. Atish Patel for the critical reading
and editing of the manuscript. This study was supported by funds
from NIH R15 No. 1R15CA143701 (Z.S.C.) and St. John's University
Seed grant No. 579-1110 (Z.S.C.).
Abbreviations:
|
MDR
|
multidrug resistance
|
|
ABC
|
ATP-binding cassette
|
|
MRP7
|
multidrug resistance protein 7
|
|
TKI
|
tyrosine kinase inhibitor
|
|
FLT3
|
FMS-like tyrosine kinase 3
|
References
|
1
|
Jemal A, Siegel R, Ward E, et al: Cancer
statistics, 2008. CA Cancer J Clin. 58:71–96. 2008. View Article : Google Scholar
|
|
2
|
Wu CP, Calcagno AM and Ambudkar SV:
Reversal of ABC drug transporter-mediated multidrug resistance in
cancer cells: evaluation of current strategies. Curr Mol Pharmacol.
1:93–105. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bradbury PA and Middleton MR: DNA repair
pathways in drug resistance in melanoma. Anticancer Drugs.
15:421–426. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Deeley RG, Westlake C and Cole SPC:
Transmembrane transport of endo-and xenobiotics by mammalian
ATP-binding cassette multidrug resistance proteins. Physiol Rev.
86:849–899. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Borges-Walmsley MI, McKeegan KS and
Walmsley AR: Structure and function of efflux pumps that confer
resistance to drugs. Biochem J. 376:313–338. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sodani K, Patel A, Kathawala RJ and Chen
ZS: Multidrug resistance associated proteins in multidrug
resistance. Chin J Cancer. 31:58–72. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chen ZS and Tiwari AK: Multidrug
resistance proteins (MRPs/ABCCs) in cancer chemotherapy and genetic
diseases. FEBS J. 278:3226–3245. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Naramoto H, Uematsu T, Uchihashi T, et al:
Multidrug resistance-associated protein 7 expression is involved in
cross-resistance to docetaxel in salivary gland adenocarcinoma cell
lines. Int J Oncol. 30:393–401. 2007.PubMed/NCBI
|
|
9
|
Hopper-Borge E, Chen ZS, Shchaveleva I,
Belinsky MG and Kruh GD: Analysis of the drug resistance profile of
multidrug resistance protein 7 (ABCC10): resistance to docetaxel.
Cancer Res. 64:4927–4930. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hopper-Borge E, Xu X, Shen T, et al: Human
multidrug resistance protein 7 (ABCC10) is a resistance factor for
nucleoside analogues and epithilone B. Cancer Res. 69:178–184.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhou Y, Hopper-Borge E, Shen T, et al:
Cepharanthine is a potent reversal agent for MRP7 (ABCC10)-mediated
multidrug resistance. Biochem Pharmacol. 77:993–1001. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Mendelsohn J and Baselga J: Status of
epidermal growth factor receptor antagonists in the biology and
treatment of cancer. J Clin Oncol. 21:2787–2799. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hirota S, Isozaki K, Moriama Y, et al:
Gain-of-function mutations of c-KIT in human gastrointestinal
stromal tumors. Science. 279:577–580. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Shah NP, Nicoll JM, Nagar B, et al:
Multiple BCR-ABL kinase domain mutations confer polyclonal
resistance to the tyrosine kinase inhibitor imatinib (STI571) in
chronic phase and blast crisis chronic myeloid leukemia. Cancer
Cell. 2:117–125. 2002. View Article : Google Scholar
|
|
15
|
Jordanides NE, Jorgensen HG, Holyoake TL
and Mountford JC: Functional ABCG2 is overexpressed on primary CML
CD34+cells and is inhibited by imatinib mesylate. Blood.
108:1370–1373. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Schittenhelm MM, Shiraga S, Schroeder A,
et al: Dasatinib (BMS-354825), a dual SRC/ABL kinase inhibitor,
inhibits the kinase activity of wild-type, juxtamembrane, and
activation loop mutant KIT isoforms associated with human
malignancies. Cancer Res. 66:473–481. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hiwase DK, Saunders V, Hewett D, et al:
Dasatinib cellular uptake and efflux in chronic myeloid leukemia
cells: therapeutic implications. Clin Cancer Res. 14:3881–3888.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Gora-Tybor J and Robak T: Targeted drugs
in chronic myeloid leukemia. Curr Med Chem. 15:3036–3051. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang H, Peng C, Hu Y, et al: The Blk
pathway functions as a tumor suppressor in chronic myeloid leukemia
stem cells. Nat Genet. 44:861–871. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Shi Z, Peng XX, Kim IW, et al: Erlotinib
(Tarceva, OSI-774) antagonizes ATP-binding cassette subfamily B
member 1 and ATP-binding cassette subfamily G member 2-mediated
drug resistance. Cancer Res. 67:11012–11020. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Dai C, Tiwari AK, Wu CP, et al: Lapatinib
(Tykerb, GW572016) reverses multidrug resistance in cancer cells by
inhibiting the activity of ATP-binding cassette subfamily B member
1 and G member 2. Cancer Res. 68:7905–7914. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tiwari AK, Sodani K, Wang SR, et al:
Nilotinib (AMN107, Tasigna) reverses multidrug resistance by
inhibiting the activity of the ABCB1/Pgp and ABCG2/BCRP/MXP
transporters. Biochem Pharmacol. 78:153–161. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Shen T, Kuang YH, Ouyang J, et al:
Imatinib and nilotinib reverse multidrug resistance in cancer cells
by inhibiting the efflux activity of the MRP7(ABCC10). PLoS One.
4:e75202009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kuang YH, Shen T, Sodani K, et al:
Lapatinib and erlotinib are potent reversal agents for MRP7
(ABCC10)-mediated multidrug resistance. Biochem Pharmacol.
79:154–161. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
DeAngelo DJ, Stone RM, Heaney ML, et al:
Phase 1 clinical results with tandutinib (MLN518), a novel FLT3
antagonist, in patients with acute myelogenous leukemia or
high-risk myelodysplastic syndrome: safety, pharmacokinetics, and
pharmacodynamics. Blood. 108:3674–3681. 2006. View Article : Google Scholar
|
|
26
|
Chen ZS, Hopper-Borge E, Belinsky MG,
Shchaveleva I, Kotova E and Kruh GD: Characterization of the
transport properties of human multidrug resistance protein 7 (MRP7,
ABCC10). Mol Pharmacol. 63:351–358. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Bliss CI: The calculation of the
dose-mortality curve. Ann Appl Biol. 22:134–167. 1935. View Article : Google Scholar
|
|
28
|
Tiwari AK, Sodani k, Dai CL, et al:
Nilotinib potentiates anticancer drug sensitivity in murine ABCB1-,
ABCG2-, and ABCC10-multidrug resistance xenograft models. Cancer
Lett. 328:307–317. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yang JJ, Milton MN, Yu S, et al:
P-glycoprotein and breast cancer resistance protein affect
disposition of tandutinib, a tyrosine kinase inhibitor. Drug Metab
Lett. 4:201–212. 2010.PubMed/NCBI
|
|
30
|
Kantarjian H, Giles F, Wunderle L, et al:
Nilotinib in imatinib-resistant CML and Philadelphia
chromosome-positive ALL. New Eng J Med. 354:2542–2551. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Tanaka C, Yin OQ, Sethuraman V, et al:
Clinical pharmacokinetics of the BCR-ABL tyrosine kinase inhibitor
nilotinib. Clin Pharmacol Ther. 872:197–203. 2010. View Article : Google Scholar : PubMed/NCBI
|