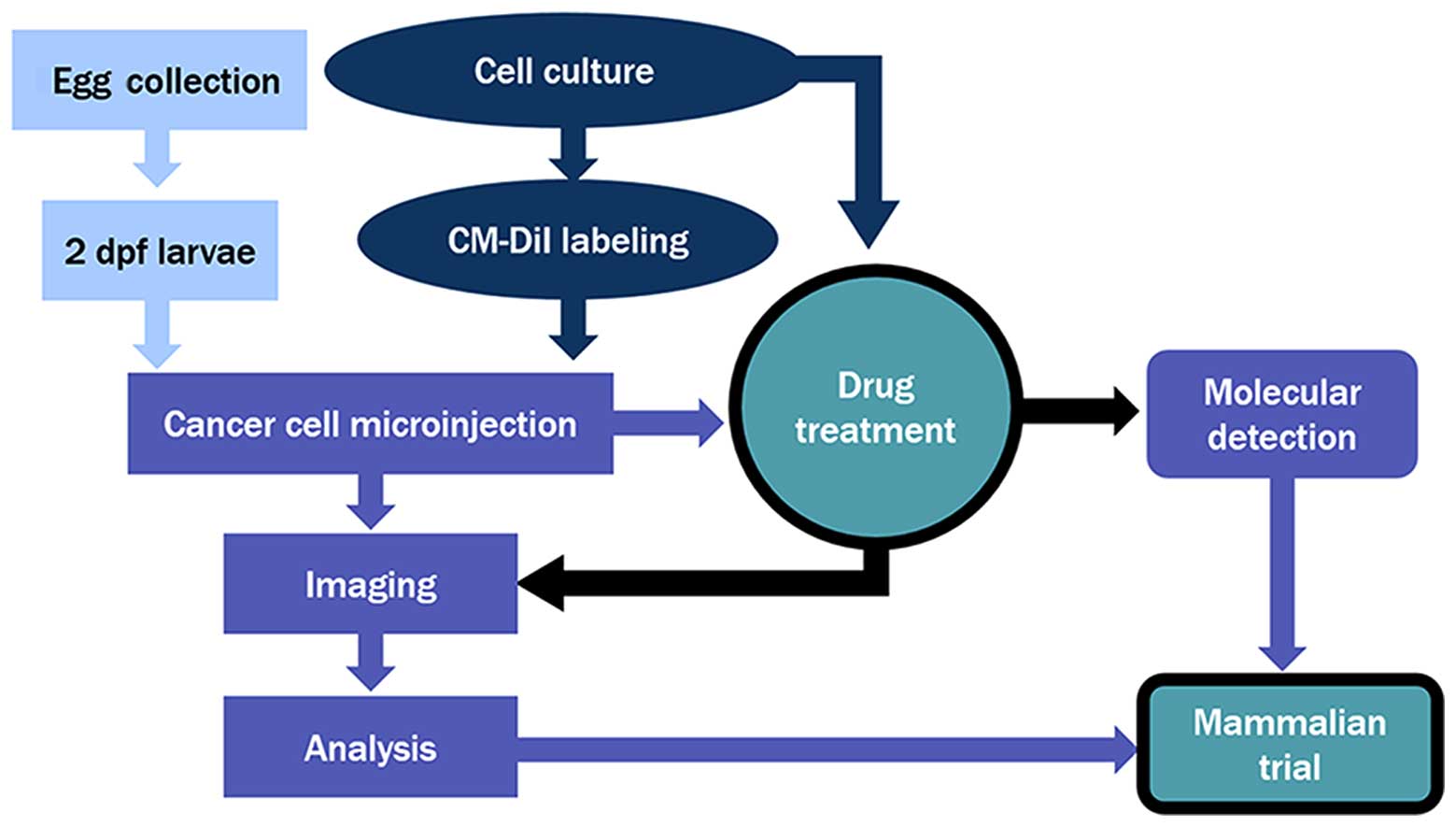
Introduction
Pancreatic cancer is one of the most aggressive
malignancies worldwide, with a median survival time of less than 6
months and with a 5-year overall survival rate less than 5%
(1). The annual death rate for
pancreatic cancer is almost equal to the incidence rates in both
developed and developing countries, suggesting the lack of an
effective screening method for targeted drug therapy (2–4). Since
traditional drug development processes are slow and expensive,
newer and more effective high-throughput animal models are greatly
needed to promote preclinical therapies for pancreatic cancer
(5). Zebrafish models have been
shown to be effective drug discovery tools due to the following
characteristics: high fecundity, small size, feasible manipulation,
easy to observe, rapid development and analogous structures to
other vertebrates (6). In addition,
the zebrafish at larval stages is transparent with an immature
immune system, making it an ideal model for cancer
xenotrans-plantation and in vivo fluorescent imaging. The
efficiency of drugs in inhibiting tumors can be observed at the
cellular level in vivo and assessed precisely at the whole
organ level (7,8). Xenotransplantation studies in
zebrafish are potentially valuable for investigating pancreatic
cancer biology, particularly for assaying high-throughput screening
drugs in vivo (9). In the
present study, we performed xenotransplantation of tumor cell lines
derived from pancreatic cancer in both larval and adult zebrafish
and demonstrated the advantages of the zebrafish as an in
vivo model by which to test potential drug therapies for
pancreatic cancer.
Activating mutations in the KRAS signaling pathway
have been observed in 90% of pancreatic cancer patient tissues,
indicating a major role for these mutations in pancreatic cancer
initiation and progression (10,11).
Aberrant activation of RAS-mediated proliferation and survival
signaling pathways in cells can be initiated by activating
mutations in KRAS. Targeting KRAS mutations may reduce cancer
mortality; nevertheless, effectively targeting of KRAS has proven
challenging for clinical treatments thus far (12). Inhibiting the downstream effector
MEK1/2, a dual-specific kinase that is required for ERK1/2
activation and that is sustained at a high level through the
Raf/MEK/ERK pathway driven by mutant KRAS, has been shown to be
effective in preclinical studies (13). Multiple small-molecule inhibitors of
MEK are now being studied in preclinical or clinical trials to
treat malignant neoplasms (14).
Identifying and characterizing druggable inhibitors of crucial
downstream effectors of KRAS have also been challenging, suggesting
that new animal models for drug screening in vivo are
required. The compound U0126 was identified as an inhibitor that
directly suppresses MEK1/2 activation (15,16).
Here, we used U0126 as an example and present a robust in
vivo system for evaluating the effectiveness of a suppressor
molecule.
First, we investigated whether xenografted Mia
PaCa-2 (human pancreatic cancer) cells could proliferate and spread
in vivo while retaining the biological features of carcinoma
in zebrafish. Then, we attempted to verify and quantify the
proliferation and migration of fluorescence-labeled pancreatic
cancer cells in vivo by imaging analysis. Additionally, we
studied the interaction between pancreatic cancer cell dispersal
and angiogenesis. Finally, we investigated whether U0126 suppressed
the proliferation of Mia PaCa-2 cells in the xenotransplanted
zebrafish larvae and compared those results with our in
vitro experiments. Our findings indicated that the zebrafish
can be utilized as a rapid and simple animal model that can be
visualized for cancer metastasis mechanistic research and
anti-pancreatic cancer molecular inhibitor screening.
Materials and methods
Cell culture and proliferation assay
The pancreatic cancer cell lines Mia PaCa-2 and
BxPC-3 were purchased from the American Type Culture Collection
(ATCC; Rockville, MD, USA) and grown in Dulbecco’s modified Eagle’s
medium (DMEM) and high-glucose RPMI-1640 (Invitrogen Life
Technologies, Carlsbad, CA, USA) supplemented with 10% fetal bovine
serum (FBS) and 1% penicillin-streptomycin at 37°C in 5%
CO2. A final concentration of 5 µM CM-DiI
(Invitrogen Life Technologies) was added to PBS-washed cells and
incubated for 15 min in a CO2 incubator at 37°C followed
by another 15-min incubation at 4°C. The cells were pelleted,
washed, and resuspended in DMEM before transplantation.
Mia PaCa-2 cells were seeded at 5,000 cells/well in
96-well plates and allowed to stabilize in DMEM complete media.
After 24 h, the cells were treated with 0, 2.5, 5, 10, 20, 40 or 80
µM U0126 in DMEM media supplemented with 0.5% FBS. The cells
were maintained for 12 h, incubated with 10%
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT;
Sigma-Aldrich, St. Louis, MO, USA) for 4 h, aspirated and dissolved
in DMSO to generate the formazan product. Absorbance was measured
at 560 nm with the reference wavelength at 700 nm.
Zebrafish and pancreatic cancer
transplantation
Larval and adult zebrafish were obtained, staged and
raised from natural spawnings of wild-type (AB) and
Tg(flk1:EGFP) zebrafish [visualization of vascular
endothelium (18)]. All the
zebrafish utilized in the subsequent experiments were maintained in
accordance with the Molecular Medicine Laboratory of Fudan
University guidelines. Zebrafish embryos were obtained using
standard mating conditions and were de-chorionized at 48 h
post-fertilization (hpf).
For transplantation of the larvae, the embryos were
anesthetized with 0.003% tricaine and positioned with their right
side up on a wet agarose pad. Cells were acquired from culture
dishes using a non-enzymatic cell-lifting solution and resuspended
in PBS at a concentration of 106 cells/50 µl and
maintained on ice before injection. Approximately 100–200 cancer
cells were injected into the perivitelline cavity of each zebrafish
larva using an Eppendorf FemtoJet injector equipped with a 0.75-mm
borosilicate glass needle. The injection parameters were as
follows: injection pressure, 20 hp; holding pressure; and injection
time, 30 sec. Immediately following transplantation, the injected
larvae were washed and selected under a fluorescence-stereo
microscope. The larvae with fluorescence-positive cells were
separated into individual containers and kept at 35.5°C.
For adult transplantation, the 6-mpf zebrafish were
irradiated with 25 Gy of γ-irradiation at 2 days before
transplantation and allowed to recover in standard fish water. On
the day of the transplant, recipients were anesthetized in 0.2%
tricaine. Using a Hamilton syringe, 10 µl of cell suspension
was transplanted into the cardiac chamber, and the fish was
immediately returned to fresh fish water. Recipients were kept off
of system water (to minimize infectious risk) for 1 week and fed
standard flake/brine shrimp meal.
All animal procedures were approved by the
Institutional Animal Care Committee, Fudan University, China.
Histological analysis
Adult fish were fixed in 4% paraformaldehyde
overnight. After the fish were processed and embedded in paraffin,
5-mm sections were cut either longitudinally or transversely.
Staining was performed with hematoxylin and eosin.
Morphological analyses and drug
treatment
Transplanted zebrafish were subsequently visually
documented on a daily basis. Live zebrafish embryos and adults were
anesthetized with tricaine before microscopic observation. Digital
micrographs were acquired using a DP70 CCD camera mounted on an
IX71 inverted fluorescence microscope (both from Olympus, Japan).
Two images were acquired in the same focal plane in bright field
and in transmitted light passing through the RFP or GFP filters and
then superimposed and processed using the Adobe Photoshop program.
In processing the fluorescent images, the RFP signal in the
zebrafish embryos changed dynamically. Image-Pro Plus 6.0 (Media
Cybernetics, Rockville, MD, USA) software was used to analyze the
imaging data after transplantation and to quantify the fluorescence
variation that represented the dynamic changes in the cell counts
and locations. Images that were more sophisticated were obtained
using light sheet microscopy (Zeiss, Germany).
For drug treatment, U0126 (Cell Signaling
Technology, Beverly, MA, USA) was dissolved in DMSO to 1,000
µM and added directly to the water at a final concentration
of 4 µM. Equivalent DMSO was used as a vehicle control. E3
medium plus drug was changed every 24 h.
Western blot analysis
Total proteins were separated by SDS-PAGE and
transferred to membranes. After blocking with milk, the membranes
were incubated in a milk solution containing phospho-p44/42 MAPK
(Erk1/2) rabbit mAb and p44/42 MAPK (Erk1/2) rabbit mAb (both from
Cell Signaling Technology). The bound antigen-antibody complex was
detected by a secondary antibody (Cell Signaling Technology) using
a chemiluminescence kit (Millipore, Germany). The same membrane was
used to determine β-actin levels, which were detected using an
anti-β-actin antibody (Sigma-Aldrich), as an internal loading
control.
KRAS mutation analysis
Mutation detection was performed by direct DNA
sequencing of kras exons 2 and 3 as described previously
(17).
Statistical analysis
The dynamic changes in the pancreatic cancer cells,
as represented by fluorescence variations counted by Image Pro Plus
6.0 analyses, were compared between the drug-treated and control
animals. Fisher’s exact test was performed in SAS 9.1 (SAS
Institute, Cary, NC, USA) to determine P-values. P-values <0.05
were considered to indicate statistically significant results.
Results
Xenografted human pancreatic cancer cells
were able to proliferate and migrate in zebrafish larvae and
adults
First, we conducted a comparison and analysis of two
human pancreatic cancer cell lines, Mia PaCa-2 and BxPC-3, to
select an appropriate source for xenografting. After the xenografts
were performed, we observed that the Mia PaCa-2 and BxPC-3 cells
proliferated and disseminated throughout the zebrafish bodies
intravitally at 6 h post-transplantation (hpt). We microinjected
CM-DiI-labeled Mia PaCa-2 cells into the perivitelline cavity of 48
hpf embryos and analyzed the larvae within 10 days by continuous
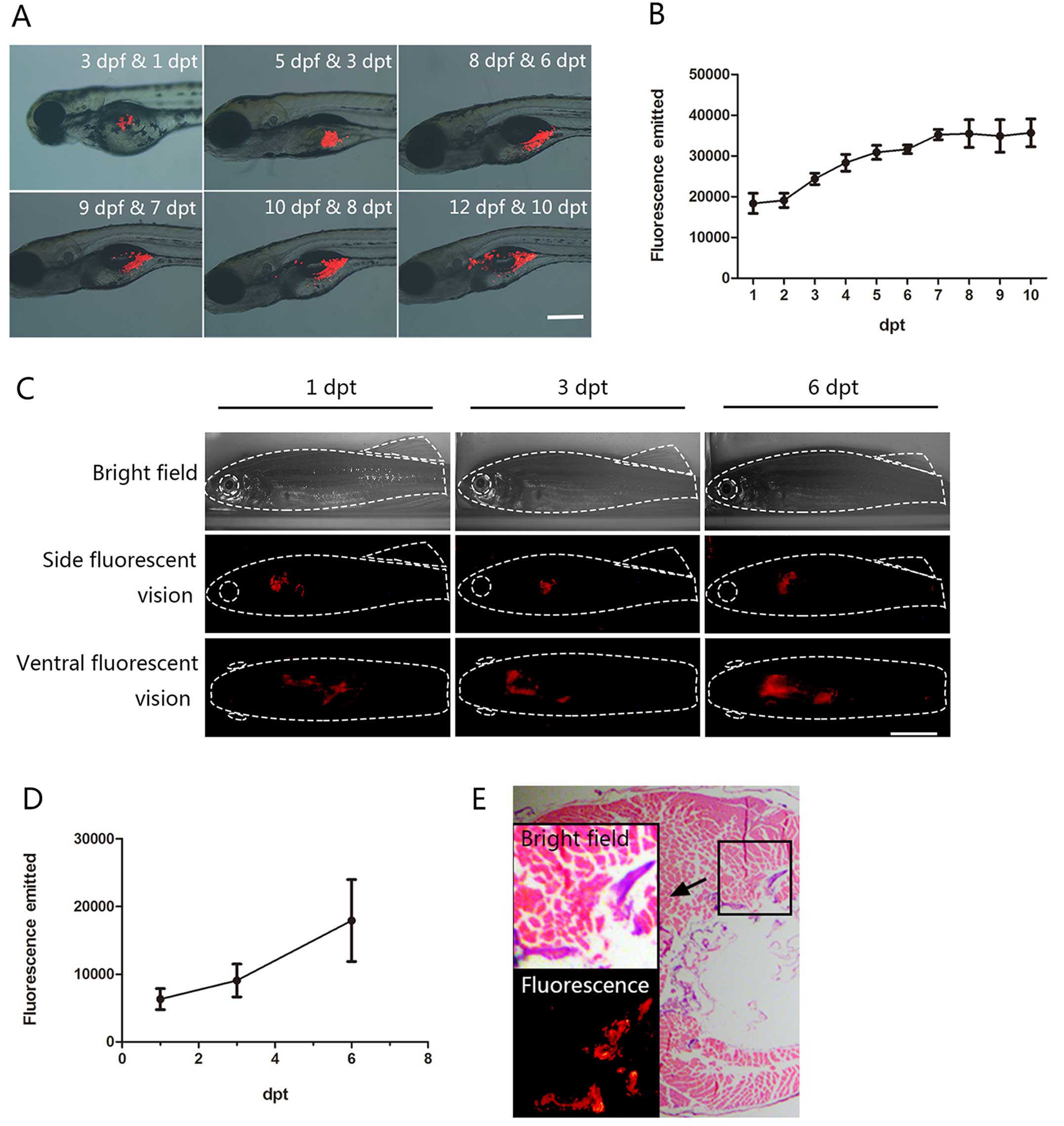
fluorescence microscopy. We noted that the majority of larvae (86%)
showed tumor cells within their bodies after a 12-h period and that
the cells were concentrated at 36 hpt (Fig. 1A). Moreover, we detected xenografted
cells that appeared to disseminate in the vitelline sac at 6 days
post-transplantation (dpt) and to spread into the body cavity and
pancreas until 10 dpt (Fig. 1A). In
addition, we confirmed CM-DiI-labeled Mia PaCa-2 cell proliferation
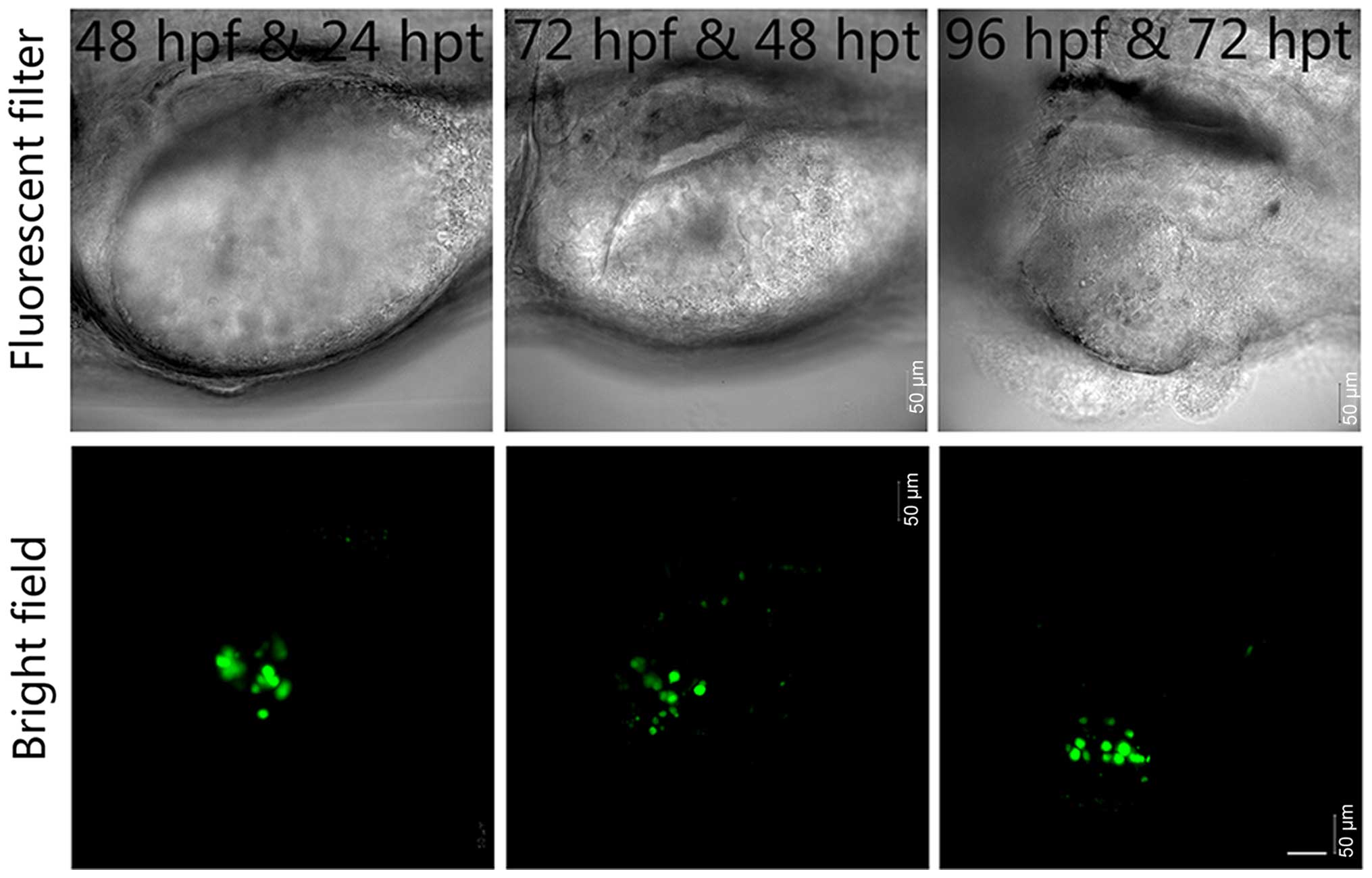
by recording and analyzing the images daily (Fig. 1B). In contrast, we found that
EGFP-labeled BxPC-3 cells barely proliferated and disseminated
outside the yolk sac region (Fig.
2). Therefore, we utilized Mia PaCa-2 cells as the grafted
cells in subsequent transplantation experiments.
To assess the broad distribution of the engrafted
cells in zebrafish tissues, we next investigated the morphological
characteristics of the CM-DiI-labeled Mia PaCa-2 cells after
xenografting in adult fish and performed histological analysis
using hematoxylin and eosin staining. We determined that the
fluorescently labeled Mia PaCa-2 cells were able to proliferate and
migrate to the pancreas within 7 days after intraperitoneal
xenotransplantation into the irradiated adult fish (Fig. 1C). Additionally, Mia PaCa-2 cell
proliferation in the adult fish was consistent with the
observations made in the juvenile fish (Fig. 1D).
To further verify the previous results, we prepared
serial tissue slices and then visualized the hematoxylin and
eosin-stained images under fluorescence. Histological analysis
revealed that the Mia PaCa-2 cells were distributed in the
digestive organs and abdominal wall (Fig. 1E). The fluorescence persisted in the
adjacent serial histological sections, demonstrating the existence
of human pancreatic cancer cells in accordance with the
histological analysis (Fig. 1E,
lower-left inset image). These results suggest that the Mia PaCa-2
human pancreatic cancer cell line is competent for use in zebrafish
xenografting. Furthermore, the proliferation of the xenografted
human pancreatic cancer cells was able to be visualized in the
living zebrafish.
Pancreatic cancer cells spread and
migrate throughout the vasculature in zebrafish larvae
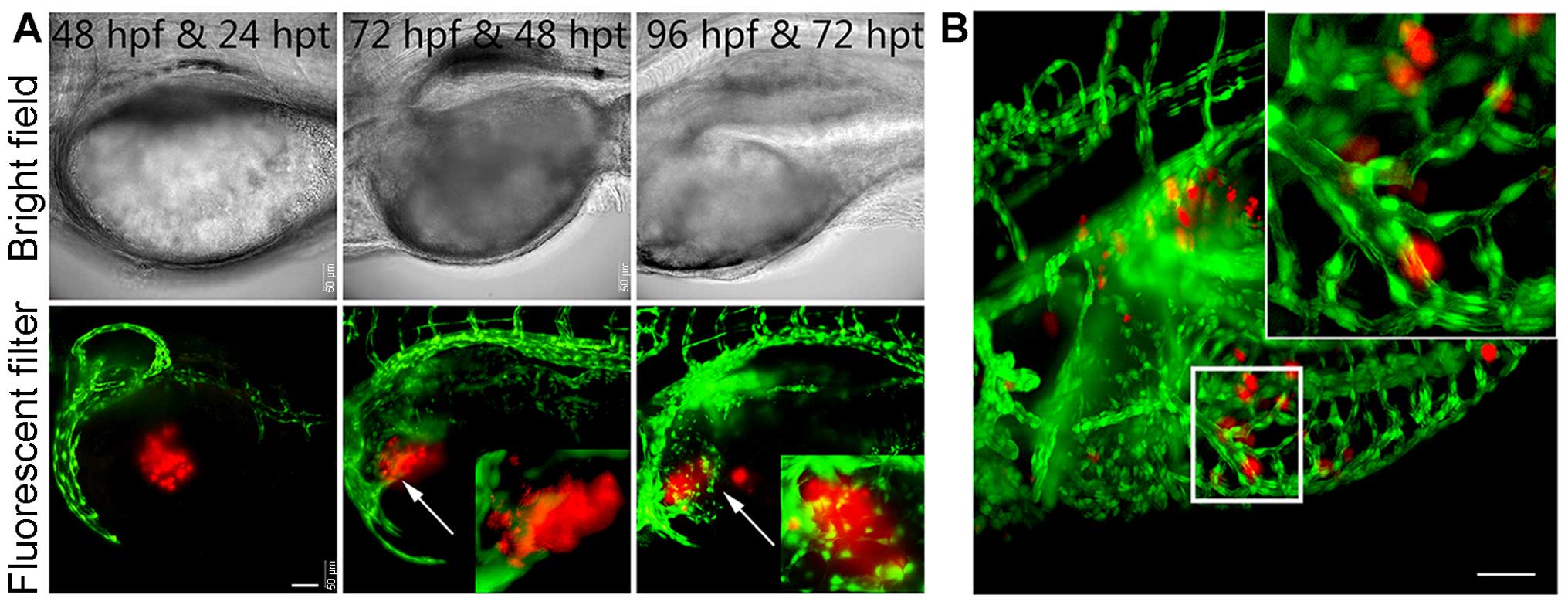
To study the initial tumor formation by the
transplanted human pancreatic cancer cells in vivo, we
investigated the morphological characteristics of these cells by
confocal laser scanning microscopy during the preliminary stage of
transplantation. Although transplanted Mia PaCa-2 cells can barely
form solid tumor masses that resemble the ones shown within human
patients and mammalian xenograft models, we identified several
CM-DiI-labeled Mia PaCa-2 cell clusters that were integrated into
the intestinal region (Fig. 3).
The later steps of invasive cancer cell progression
involve locating vessels, invading and inducing angiogenesis, which
facilitates the extravasation of the cells to form secondary tumors
(18,19). To investigate the process of
pancreatic cancer cell invasion of the vascular system of zebrafish
and induction of angiogenesis, we performed microinjections of
CM-DiI-labeled Mia PcCa-2 cells into the yolk and primordial cavity
of Tg(flk1:EGFP) transgenic zebrafish larvae at 2 dpf
(18). The human pancreatic cancer
cluster was initially distinct from the developing vasculature of
the larvae (Fig. 3A, left image).
Over time, however, the xenografted Mia PaCa-2 cells had a tendency
to migrate to the vasculature (Fig.
3A, middle image), and the developing zebrafish vasculature
extended toward and directly contacted the tumor clusters (Fig. 3A, middle and right images). These
results indicated that the human pancreatic cancer cells stimulated
angiogenesis and recruited blood vessels from the host larvae that
subsequently supported the continued growth of the tumor.
To study the effect of the developing vascular
distribution on the diffusion of the human pancreatic cancer cells,
we further observed the advanced aspects of xenotransplanted
zebrafish. Approximately 30% of the xenografted larvae exhibited
Mia PaCa-2 cell clusters that diffused within the invading blood
vessels and that spread throughout the vascu-lature (Fig. 3B). Therefore, all of these results
corroborated the findings that pancreatic cancer cells were able to
spread throughout the vasculature and stimulate angiogenesis in the
zebrafish.
Treatment with U0126 deregulates KRAS
activity in human pancreatic cancer cells in live zebrafish
To investigate whether larval zebrafish are suitable
for in vivo drug screening, we tested the effects of a
targeted inhibitor of KRAS signaling on xenotransplanted human
pancreatic cancer cells in our zebrafish model. The compound U0126
is able to directly suppress MEK1/2 activation, which is critical
for transmitting signals to ERK (15). This effector can be abnormally
sustained in an activated state by mutated KRAS signaling and is
thus an attractive pharmaceutical target for tumors harboring
aberrant MAPK pathway signaling (11,12,20).
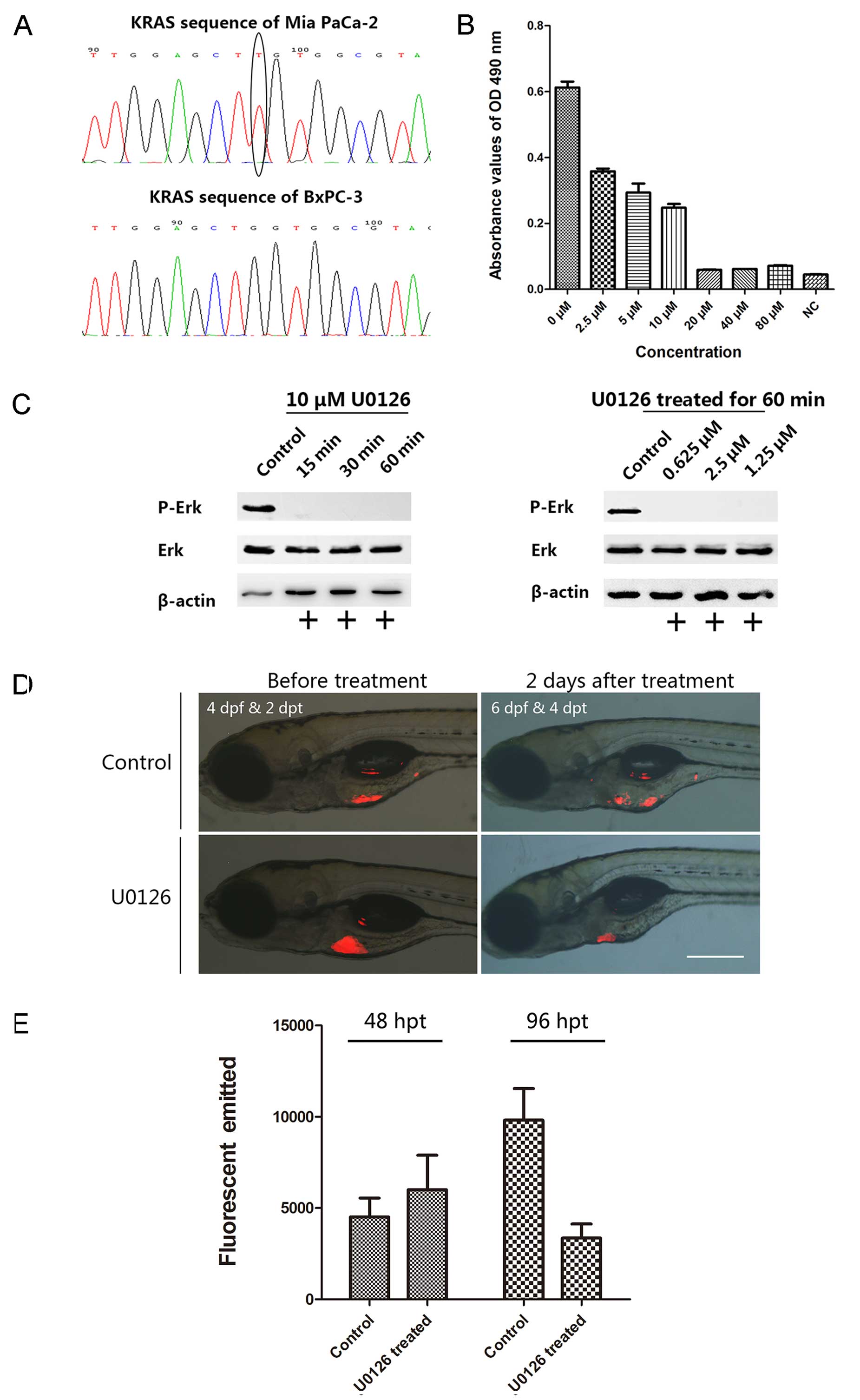
First, we performed kras gene sequencing in
the two cell lines to investigate the inhibitory effect of U0126 on
human pancreatic cancer cells with KRAS mutations. Our sequencing
results showed that BxPC-3 cells expressed the wild-type
kras gene, whereas Mia PaCa-2 cells had a missense mutation
at codon 12 (Fig. 4A). In addition,
we verified that U0126 exerted an inhibitory effect on Mia PaCa-2
cell proliferation in vitro. U0126 treatment lead to
dose-dependent suppression of cell proliferation at concentrations
ranging from 2.5 to 80 µM, and the maximum inhibitory
concentration was 20 µM (Fig.
4B).
To validate the U0126-induced inhibition of MEK1/2
phosphorylation to suppress mutated KRAS signaling, we tested the
time- and dosage-effect relations after treating Mia PaCa-2 cells
with U0126 and then investigated ERK phosphorylation. Western blot
analysis showed that MEK phosphorylation in Mia PaCa-2 cells was
inhibited by U0126 treatment after a short incubation period (15
min) and at low concentrations (0.625 µM) (Fig. 4C). MEK phosphorylation is critical
to the activation of the primary route of signal transmission in
the MAPK pathway, and MEK phosphorylation is inhibited by U0126
(21). These results suggest that
U0126 can suppress Mia PaCa-2 cell proliferation by inhibiting MEK
phosphorylation.
Next, we assessed the efficacy of U0126 to validate
our zebrafish xenograft model system. As demonstrated in the above
experiments, U0126 inhibited cell proliferation in vitro in
a dose-dependent manner, and the inhibitory concentration was used
as the basis of the IC50 value calculation. In our
xenograft model, U0126 produced a prompt effect that suppressed
cell proliferation (Fig. 4D). We
exposed xenotrans-planted larvae to U0126 for 48 h at a final
concentration of 4 µM in an aqueous medium. The
xenotransplanted larvae in the experimental group exhibited
29.28±9.2% cancer cell degeneration following U0126 treatment in
60.0% (36 in 60) of the larvae. In contrast, the corresponding
control group (untreated) showed 121.05±37.2% cancer cell
proliferation in 85.0% (51 in 60) of the larvae. Representative
images acquired before and after processing are shown in Fig. 4D. Moreover, statistical analysis
showed that U0126 significantly inhibited Mia PaCa-2 cell
proliferation compared with the in vivo results (P<0.05,
Fig. 4E). In addition, we detected
fewer metastases in the bodies of the larvae in the experimental
group; however, the mock-treatment group displayed frequent
metastasis. These assays indicated that U0126 can suppress the
proliferation and metastasis of pancreatic cancer cells by
inhibiting KRAS signaling downstream effector activation both in
vivo and in vitro, suggesting that this
xenotransplantation pancreatic cancer model utilizing larval
zebrafish is useful for facilitating in vivo drug
screening.
Discussion
The development of a proper animal model test will
provide an efficient platform for investigating novel drugs.
Utilizing xenograft models that recapitulate key processes of
cancer development may be extremely favorable for screening large
numbers of small-molecule compounds in an in vivo setting.
Presently, several targeted therapeutic strategies have been used
in pancreatic cancer preclinical studies (22). Pancreatic cancer genetic alterations
in the KRAS signaling pathway are involved in over 90% of human
pancreatic cancer cases. The components in this pathway are
promising targets for identifying novel therapies (23). The most common KRAS mutation in
human pancreatic cancer is a gain-of-function mutation at codon 12.
Moreover, pancreatic cancer cell growth has been shown to be
dependent on the activity of the mutated kras gene;
accordingly, silencing the kras gene has been shown to be
effective in controlling pancreatic cancer cell line proliferation
(24). However, the clinical
development of targeted agents for pancreatic cancer has not been
successful. Most known inhibitors have already been processed
preclinically and represent promising targets for therapeutic
intervention; however, these inhibitors lack concise and effective
in vivo studies (11). In
the present study, we harnessed the advantages of xenograft models
of pancreatic cancer in zebrafish for KRAS signaling pathway
inhibitor screening.
The zebrafish model offers a rapid and inexpensive
platform with which to investigate the proliferation and metastatic
potential of human cancer cells. Moreover, the larvae
experimentation allowed us to intuitively evaluate the mechanism by
which U0126 targets tumor cell signaling pathways. In the present
study, we described this zebrafish xenograft model, which provides
an excellent platform to study human pancreatic cancer cell
proliferation and migration and the relationship with angiogenesis
in vivo. The study indicated that Mia PaCa-2 cells with
kras codon 12 mutation showed stronger proliferative and
metastatic ability than BxPC-3 cells in vivo (Figs. 1, 2
and 4A). These result suggest that
xenotrans-plantation of cells with KRAS mutation was able to
simulate pancreatic cancer initiation and progression in
vivo.
In the present study, we utilized the KRAS signaling
inhibitor U0126 as an example to test the feasibility of this model
for high-throughput screening. Although the pharmacological
characteristics of U0126 indicated that this drug was unsuitable
for clinical development, the small-molecule compound was
ultimately demonstrated to be an efficient inhibitor when used in
multiple studies (13,25). We first described the inhibitory
role of U0126 in suppressing Mia PaCa-2 cell proliferation in
vitro due to the KRAS activating mutation in these cells. In
addition, we tested ERK phosphorylation following U0126 treatment
to verify that U0126 could target the downstream effectors of KRAS
signaling. Finally, we validated our findings that the
proliferation of transplanted Mia PaCa-2 cells could be inhibited
by exposure to U0126, which was consistent with the findings of
another study (26). The image
analysis combined with the statistical results also showed that
U0126 markedly suppressed cell growth in the majority of
transplanted individuals. All these results demonstrated that this
elementary and reliable xenotransplantation model in zebrafish is
effective for investigating anti-pancreatic cancer drugs.
Metastasis is the major cause of death in pancreatic
cancer patients and is not fully understood. An intriguing
observation in our study was the phase relationship between the
transfer of pancreatic cancer cells and the vascular development of
larvae. We dynamically detected that xenografted Mia PaCa-2 cells
migrated to the host vascular system, induced angiogenesis and
diffused through developing vessels in the Tg(flk1:EGFP)
larvae. Neovascularization could offer the necessary material for
tumor metabolism (27), although
these components are not yet clear. In contrast, xenotransplanted
Mia PaCa-2 cells may recruit new blood vessels; this recruitment is
one of the typical forms of hematogenous metastasis. Therefore, we
propose that the zebrafish xenograft model may allow us to
investigate hematogenous pancreatic cancer cell metastasis and
tumor-induced angiogenesis in vivo.
Transplantation of primary cancer cells from tissue
culture could offer a more realistic model than cell lines. All of
our analyses were performed using human pancreatic cancer cell
lines, which confirmed the use of CM-DiI labeling as an effi-cient
and feasible technique (28).
Primary cancer cells can also be tagged through this method and
processed as immortal cell lines. Therefore, this platform could
potentially be used for screening novel therapeutic agents that
block cancer invasion and metastasis similar to clinical studies.
The results demonstrated that this robust zebrafish
xenotransplantation model provides an extremely encouraging
solution for evaluating the proliferation potential of primary
cells from biopsies or following surgery, a brilliant advantage for
clinical studies. Furthermore, a variety of candidate MEK
inhibitors can be screened for their ability to suppress pancreatic
cancer by targeting multiple pathways (25). A typical screening using the larvae
xenograft model can be completed under suitable time and space
conditions (Fig. 5).
Taken together, our present study describes a
zebrafish xenograft model that can be used to evaluate the
inhibitory potential of KRAS pathway signaling inhibitors in human
pancreatic cancer cell lines in vivo. This zebrafish model
of xenotransplanted human pancreatic cancer cells showed a
significant ability to perform tumor characteristic analysis in
large cohorts of fish, allowing a robust statistical analysis in a
short time frame (~12 days) and providing a rapid means of
evaluating the effects of anticancer drugs in vivo. Based on
this reliable model, new therapeutic drugs that block pancreatic
cancer invasion and proliferation could potentially be developed by
targeting the KRAS signaling pathway.
Acknowledgments
We acknowledge Professor H.Y. Song for her kind help
in this study. The present study was supported in part by the
National Natural Science Foundation of China (no. 81402582, no.
21202021) and by Shanghai Science and Technology Development Funds
(no. 14YF1400600).
References
|
1
|
Hidalgo M: Pancreatic cancer. N Engl J
Med. 362:1605–1617. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Long J, Luo GP, Xiao ZW, Liu ZQ, Guo M,
Liu L, Liu C, Xu J, Gao YT, Zheng Y, et al: Cancer statistics:
Current diagnosis and treatment of pancreatic cancer in Shanghai,
China. Cancer Lett. 346:273–277. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Siegel R, Ma J, Zou Z and Jemal A: Cancer
statistics, 2014. CA Cancer J Clin. 64:9–29. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Singh HM, Ungerechts G and Tsimberidou AM:
Gene and cell therapy for pancreatic cancer. Expert Opin Biol Ther.
15:505–516. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
White R, Rose K and Zon L: Zebrafish
cancer: The state of the art and the path forward. Nat Rev Cancer.
13:624–636. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Geiger GA, Fu W and Kao GD:
Temozolomide-mediated radio-sensitization of human glioma cells in
a zebrafish embryonic system. Cancer Res. 68:3396–3404. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Berghmans S, Jette C, Langenau D, Hsu K,
Stewart R, Look T and Kanki JP: Making waves in cancer research:
New models in the zebrafish. Biotechniques. 39:227–237. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang B, Shimada Y, Kuroyanagi J,
Nishimura Y, Umemoto N, Nomoto T, Shintou T, Miyazaki T and Tanaka
T: Zebrafish xenotransplantation model for cancer stem-like cell
study and high-throughput screening of inhibitors. Tumour Biol.
35:11861–11869. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Jones S, Zhang X, Parsons DW, Lin JC,
Leary RJ, Angenendt P, Mankoo P, Carter H, Kamiyama H, Jimeno A, et
al: Core signaling pathways in human pancreatic cancers revealed by
global genomic analyses. Science. 321:1801–1806. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Eser S, Schnieke A, Schneider G and Saur
D: Oncogenic KRAS signalling in pancreatic cancer. Br J Cancer.
111:817–822. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zorde Khvalevsky E, Gabai R, Rachmut IH,
Horwitz E, Brunschwig Z, Orbach A, Shemi A, Golan T, Domb AJ, Yavin
E, et al: Mutant KRAS is a druggable target for pancreatic cancer.
Proc Natl Acad Sci USA. 110:20723–20728. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Montagut C and Settleman J: Targeting the
RAF-MEK-ERK pathway in cancer therapy. Cancer Lett. 283:125–134.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Frémin C and Meloche S: From basic
research to clinical development of MEK1/2 inhibitors for cancer
therapy. J Hematol Oncol. 3:82010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Davies SP, Reddy H, Caivano M and Cohen P:
Specificity and mechanism of action of some commonly used protein
kinase inhibitors. Biochem J. 351:95–105. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Favata MF, Horiuchi KY, Manos EJ, Daulerio
AJ, Stradley DA, Feeser WS, Van Dyk DE, Pitts WJ, Earl RA, Hobbs F,
et al: Identification of a novel inhibitor of mitogen-activated
protein kinase kinase. J Biol Chem. 273:18623–18632. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kim ST, Lim H, Jang KT, Lim T, Lee J, Choi
YL, Jang HL, Yi JH, Baek KK, Park SH, et al: Impact of KRAS
mutations on clinical outcomes in pancreatic cancer patients
treated with first-line gemcitabine-based chemotherapy. Mol Cancer
Ther. 10:1993–1999. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Stoletov K, Montel V, Lester RD, Gonias SL
and Klemke R: High-resolution imaging of the dynamic tumor cell
vascular interface in transparent zebrafish. Proc Natl Acad Sci
USA. 104:17406–17411. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Gupta GP and Massagué J: Cancer
metastasis: Building a framework. Cell. 127:679–695. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Morris JP IV, Wang SC and Hebrok M: KRAS,
Hedgehog, Wnt and the twisted developmental biology of pancreatic
ductal adenocarcinoma. Nat Rev Cancer. 10:683–695. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang X and Studzinski GP: Phosphorylation
of raf-1 by kinase suppressor of ras is inhibited by ̔MEK-specific̓
inhibitors PD 098059 and U0126 in differentiating HL60 cells. Exp
Cell Res. 268:294–300. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Vaccaro V, Melisi D, Bria E, Cuppone F,
Ciuffreda L, Pino MS, Gelibter A, Tortora G, Cognetti F and Milella
M: Emerging pathways and future targets for the molecular therapy
of pancreatic cancer. Expert Opin Ther Targets. 15:1183–1196. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
McCubrey JA, Steelman LS, Chappell WH,
Abrams SL, Wong EW, Chang F, Lehmann B, Terrian DM, Milella M,
Tafuri A, et al: Roles of the Raf/MEK/ERK pathway in cell growth,
malignant transformation and drug resistance. Biochim Biophys Acta.
1773:1263–1284. 2007. View Article : Google Scholar
|
|
24
|
Réjiba S, Wack S, Aprahamian M and Hajri
A: K-ras oncogene silencing strategy reduces tumor growth and
enhances gemcitabine chemotherapy efficacy for pancreatic cancer
treatment. Cancer Sci. 98:1128–1136. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
McCubrey JA, Steelman LS, Abrams SL,
Chappell WH, Russo S, Ove R, Milella M, Tafuri A, Lunghi P, Bonati
A, et al: Emerging MEK inhibitors. Expert Opin Emerg Drugs.
15:203–223. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Gysin S, Lee SH, Dean NM and McMahon M:
Pharmacologic inhibition of RAF→MEK→ERK signaling elicits
pancreatic cancer cell cycle arrest through induced expression of
p27Kip1. Cancer Res. 65:4870–4880. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Bai X, Zhi X, Zhang Q, Liang F, Chen W,
Liang C, Hu Q, Sun X, Zhuang Z and Liang T: Inhibition of protein
phosphatase 2A sensitizes pancreatic cancer to chemotherapy by
increasing drug perfusion via HIF-1α-VEGF mediated angiogenesis.
Cancer Lett. 355:281–287. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ji F, Duan HG, Zheng CQ and Li J:
Comparison of chloromethyl-dialkylcarbocyanine and green
fluorescent protein for labeling human umbilical mesenchymal stem
cells. Biotechnol Lett. 37:437–447. 2015. View Article : Google Scholar
|