Introduction
In China, bladder cancer is the urinary tract tumor
with the highest incidence (1).
Urothelial carcinoma is the main pathological type of bladder
cancer, and ~70% of patients are initially diagnosed with
non-muscle invasive bladder cancer (NMIBC), which has a recurrence
rate of 50–70%. Of the patients who experience recurrence, 10–20%
progress to muscle invasion of bladder cancer (MIBC) (2). Furthermore, ~30% of patients with an
initial diagnosis of MIBC have distant metastases and a high risk
of death (3). The pathogenesis of
bladder cancer is not clear, but the hypothesis that chronic
inflammation can promote tumor development has been recognized. It
is reported that 50% of bladder cancers are related to chronic
inflammation (4).
Interleukin 23 (IL-23) is an important
proinflammatory cytokine (5). Our
previous study found that IL-23 expression in bladder cancer tissue
was associated with tumor-infiltrating dendritic cells (DCs) and
that the IL-23 expression levels in cancer tissue were
significantly increased compared to that in adjacent tissues,
suggesting that IL-23 may be related to the progression of bladder
cancer (6). It has been reported
that in the human tumor microenvironment is mainly derived from
inflammatory DCs (7), and that
IL-23 is the key linker molecule between the tumor-induced
proinflammatory process and adaptive tumor immunosuppression. IL-23
and IL-12 belong to the same family of proinflammatory heterodimer
cytokines (8). The two cytokines
co-possess the p40 subunit, which form IL-12 with p35 through a
covalent bond, and form IL-23 with p19. IL-23 induces and amplifies
Th17 cells (9) and IL-12 induces
and amplifies Th1 cells (10).
IL-23 is an essential cytokine for the maintenance of the normal
number and function of Th17 cells and the secretion of IL-17 by
Th17 cells (11). In addition to
having similar functions with partial IL-12, IL-23 is also able to
promote the autoimmune response and occurrence of immune-related
inflammatory diseases through the IL-17 pathway. The balance
between IL-12 and IL-23 is very important in the process of
tumorigenesis. Mounting evidence suggests that IL-12 and IL-23 can
independently play their respective roles without relying on
interferon-γ (IFN-γ) and IL-17A, respectively (12). At present, there is little research
on whether IL-23 can play a direct role in tumor cells; the
relationship between IL-23 and bladder cancer especially remains
unclear.
In the present study, we further investigated the
relationship between IL-23 and bladder cancer through Oncomine and
TCGA database analysis, clinical specimen testing and in
vitro cell experiments.
Materials and methods
Patients and samples
The tissue specimens were obtained from 10 MIBC
patients who were treated at Daping Hospital of the Third Military
Medical University (Chongqing, China) from March 2012 to December
2014. The study subjects included 2 females and 8 males, with an
average age of 63 years (45–79 years). The pathological type of all
specimens was independently determined by two pathologists. Tumor
diagnosis and grading followed the International Union Against
Cancer TNM Classification criteria (13); the staging criteria followed the WHO
Classification of Tumors (14). The
exclusion criteria were: i) non-urothelial tumors; ii) patients
with an immune system disorder or receiving an immunosuppressive
agent; and iii) patients who had undergone preoperative
radiotherapy or immunotherapy. The tissue samples were rapidly
frozen in liquid nitrogen immediately after dissection and stored
in liquid nitrogen until further assessed by quantitative real-time
PCR. The study was approved by the Ethics Committee on Human
Experimentation of Daping Hospital of the Third Military Medical
University and informed written consent was obtained from all
patients. All collected samples were then eligible for experimental
purposes.
Oncomine database and The Cancer
Genome Atlas (TCGA) analysis
The expression level of the IL23A gene in
bladder urothelial carcinoma was assessed using the Oncomine
(www.oncomine.org) database (IL-23A is
indicated in Oncomine database and The Cancer Genome Atlas to
represent gene IL-23) (15).
Hence, we compared the clinical specimens of cancer and normal
bladder mucosa tissues (including paracancerous tissue). To reduce
the false discovery rate, we selected P<1×10−4 as the
threshold and analyzed the results of P-values, fold change and
cancer subtype. Correlation of IL23A expression with
clinicopathological parameters of bladder urothelial carcinoma was
analyzed using the TCGA BLCA-Bladder Urothelial Carcinoma (Nature
2014) data set (16).
Cells, reagents and transfection
The human bladder urothelial carcinoma cell lines
were purchased from the Type Culture Collection of the Chinese
Academy of Sciences (Shanghai, China). T24, SW-780, J82 and UM-UC-3
cell lines were treated with Dulbecco's modified Eagle's medium
(DMEM)/F12 medium containing 10% fetal bovine serum (FBS). At 48 h
after transfection, the cells were collected for qRT-PCR assays.
Recombinant human IL-23 (IL-23) was purchased from R&D Systems
(Minneapolis, MN, USA).
RNA isolation and quantitative
RT-PCR
Total RNA was isolated from cultured cells or tissue
using a Total RNA Isolation kit (Takara Biotechnology, Co., Ltd.,
Dalian, China), followed by reverse transcription and
SYBR-Green-based RT-PCR analysis (Takara Biotechnology). IL-23p19,
IL-23 receptor (IL-23R), IL-17 and IL-6 were assayed by qRT-PCR
using PrimeScript RT Reagent kit, SYBR-Green Real-Time PCR Master
Mix and Premix ExTaq (all from Takara Biotechnology) following the
manufacturer's protocols.
These primers purchased from Sangon Biotech Co.,
Ltd. (Shanghai, China) are listed in Table I. IL-23p19, IL-23R, IL-17 and IL-6
levels were normalized to the β-actin expression level. The
2−ΔΔCq method was conducted for analyzing gene
expression (17). At least three
independent experiments were conducted for each experimental
condition.
 | Table I.Primer sequences. |
Table I.
Primer sequences.
| Gene | Primer
sequences |
|---|
| IL-6 |
5′-AAATGCCAGCCTGCTGACGAAG-3′ |
|
|
3′-CATCGTACCCGTGAGTCTAACAACAA-5′ |
| IL-17A |
5′-AATACAACCGATCCACCTCAC-3′ |
|
|
3′-GACACTAGACCCTCCGTTACT-5′ |
| Il-23p19 |
5′-TGTGAATGACTTGGTCCCTGAA-3′ |
|
|
3′-GACACTAGACCCTCCGTTACT-5′ |
| Il-23R |
5′-GCAAACGCACTAGGCATGGAAG-3′ |
|
|
3′-CTCTATGTTCCGATGTTGTTTGGTT-5′ |
| β-actin |
5′-CTCTTCCAGCCTTCCTTCCT-3′ |
|
|
3′-GACATGCGGTTGTGTCACGA-5′ |
Colony formation assay
T24 cells were plated in each well of a 6-well plate
at a density of 500 cells/well and treated with IL-23 (0, 10, 20 or
40 ng/ml). When the plates appeared visibly cloned, the clones were
fixed with methanol and stained by crystal violet for 20 min. The
number of clones was over 50 cells in wells and was counted using a
light microscope (Eclipse E800M; Nikon, Tokyo, Japan). At least
three independent experiments were performed.
CCK-8 method for cell viability
T24 cells that were pre-treated with different
concentrations of IL-23 (0, 10, 20 or 40 ng/ml) for 48 h were
seeded at a density of 2×103 cells/well in independent
96-well plates and the media were changed to media without IL-23.
No vehicle control was used. The cell proliferation was then
detected using Cell Counting Kit-8 assay (CCK-8; Dojindo Molecular
Technologies, Beijing, China) following the manufacturer's
protocol; 2×103 cells were plated in 96-well
microplates, followed by the addition of 10 µl of CCK-8 (Dojindo
Molecular Technologies) solution to each well and incubation of the
samples for 1 h before measuring the absorbance at 450 nm in a
microplate autoreader El309 (Bio-Tek Instruments, Inc., Woonoski,
VT, USA). Experiments were performed in triplicate. No vehicle
control was used.
Immunofluorescence
The cells were divided into the control group, the
IL-23 10-ng/ml group, the IL-23 20-ng/ml group and the IL-23
40-ng/ml group, in triplicate in each group. The culture coverslips
with a diameter of 18 mm were placed in 24-well plates and
1×104 cells were inoculated in each well. When the cell
density was increased to 40%, it was replaced with the serum-free
DMEM/F12 and then starved for 2 h to make the cells' growth cycle
consistent. After incubation for 48 h at 37°C in a 5%
CO2 cell incubator, the cells were washed three times
with phosphate-buffered saline (PBS) (0.1 mol/l), fixed in 4%
paraformaldehyde for 15 min, washed three times with PBS solution,
placed in 0.03% Triton X-100 for 20 min to improve cell membrane
permeability, flushed with PBS 3 times, and then sealed with
immunofluorescence blocking fluid for 20 min. Anti-Ki-67 (rabbit
polyclonal to Ki-67, 1:100; cat. no. ab15580; Abcam, Cambridge, MA,
USA) and Anti-Histone H3 (phospho-histone S28) (HTA28, 1:100; cat.
no. ab10543; Abcam) were added dropwise and were uniformly covered
on the slide and then placed in a refrigerator at 4°C overnight.
After washing with PBS solution 3 times (5 min/time), goat
anti-rabbit IgG secondary antibody (cat. no. W10816; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) was added dropwise and
incubated in a wet box at 37°C for 1 h and washed 3 times (5
min/time) in PBS solution. DAPI staining was added dropwise and
incubated at 37°C for 3 min. After washing 3 times (5 min/time) in
PBS solution, a 10 µl anti-fluorescent decaying tablet was added
dropwise to coat cells, followed by sealing with clean coverslips
and images were acquired with Leica Application Suite (Version
4.2.0; Leica Microsystems, Oberkochen, Germany) under a Leica DM
4000B microscope.
Wound-healing and Transwell
assays
Before wound-healing or Transwell assays were
conducted, the cells of each group were treated with IL-23 as
aforementioned. The wound-healing assay was performed by scratching
the single cell layer with tips. The images of the scratch area
were recorded at three random spots at 0 and 24 h. A standard size
field was used to measure the migrating distance of the wound edge
for each image. An inverted microscope (Carl Zeiss Axiovert 25C;
Carl Zeiss, Göttingen, Germany) was used to examine the scratch
wounds. The mean migrating distances of the three spots were
calculated according to the scale plate. Wound-healing assays were
performed in triplicate and all data were statistically
processed.
The Transwell assay was used to test cell migration
and invasion abilities. For the migration assay, T24 cells were
placed into the upper chamber of each insert (Corning Inc.,
Corning, NY, USA). For the invasion assay, the cells were placed
into the upper chamber of inserts coated with 45 µg of diluted
Matrigel (2 µg/µl); 600 µl medium (containing 10% FBS) was added to
the lower chambers. After 24 h of incubation, a cotton tip was used
to wipe the upper surface of the membrane and cells that were
attached to the lower surface were stained for 20 min with crystal
violet and then rinsed in PBS prior to light microscopic inspection
(Eclipse E800M; Nikon). Invasion values were obtained by counting
eight fields per membrane, which represent the average of three
independent experiments.
Western blot analysis
Protein was collected from 2×106 cells
after IL-23 treatment for 48 h with lysis buffer containing
protease and phosphatase inhibitors (Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany). Cell lysates were centrifuged at 12,000 × g at
4°C for 15 min. Concentration was measured using BCA methods and 30
ng was loaded per lane and run to separate on 10% sodium dodecyl
sulfate-polyacrylamide gels. The proteins were transferred onto
polyvinylidene fluoride (PVDF) membranes (Merck Millipore,
Billerica, MA, USA) after 90 min of electrophoresis. The membranes
were blocked with 5% non-fat milk in 0.1% TBS-Tween-20 at room
temperature for 1 h and were then incubated with anti-GAPDH
(1:1,000 dilution; cat. no. ab8245; Abcam) at 4°C overnight,
followed by incubation with LI-COR IRDye 680-labeled secondary
antibodies (1:10,000 dilution; cat. no. KFC200; Rockland
Immunochemicals, Gilbertsville, PA, USA) for 1 h at room
temperature. Signals were detected with an Odyssey Infrared Imaging
system (LI-COR Biosciences, Lincoln, NE, USA) and quantified using
the FluorChem 8900 system (Alpha Innotech, San Leandro, CA,
USA).
Statistical analysis
All data were analyzed with SPSS 19.0 software (IBM
Corp., Armonk, NY, USA). Data were expressed as the mean values ±
standard deviation (SD). Comparisons between two groups were
carried out using Student's t-test. Comparisons of multiple groups
were analyzed using one way analysis of variance (ANOVA) followed
by the Student-Newman-Keuls (SNK) post hoc test. The Chi-square
test was performed to determine the association between the
IL23A expression and the clinicopathological parameters of
bladder cancer. Differences were considered statistically
significant at P<0.05.
Results
Expression of IL-23, IL-23R, IL-6 and
IL-17 in bladder urothelial carcinoma tissues
To study the effect of IL-23 on bladder urothelial
carcinoma, we first detected the expression levels of IL-23 mRNA
and IL-23R mRNA in human MIBC and adjacent tissues, and found that
both expression levels were significantly increased in cancer
tissues, which was consistent with our previously reported
immunization results (Fig. 1A-D).
In recent years, a number of studies about breast, prostate and
gastric cancer revealed that the number of Th17 cells was
significantly increased in the tumor microenvironment and its
secreted IL-17 promoted the development of tumors through the
induction of chronic inflammation (18). The initial differentiation of Th17
cells is dependent on IL-6, but IL-23 is a key cytokine in the
process of survival, proliferation and secretion of IL-17 (11). Therefore, we also detected the
expression of IL-6 and IL-17, and determined that IL-6 mRNA and
IL-17 mRNA were also significantly increased in MIBC tissues,
indicating that IL-23, IL-6, IL-17 and IL-23R were increased in the
bladder tumor microenvironment (Fig.
1D-G).
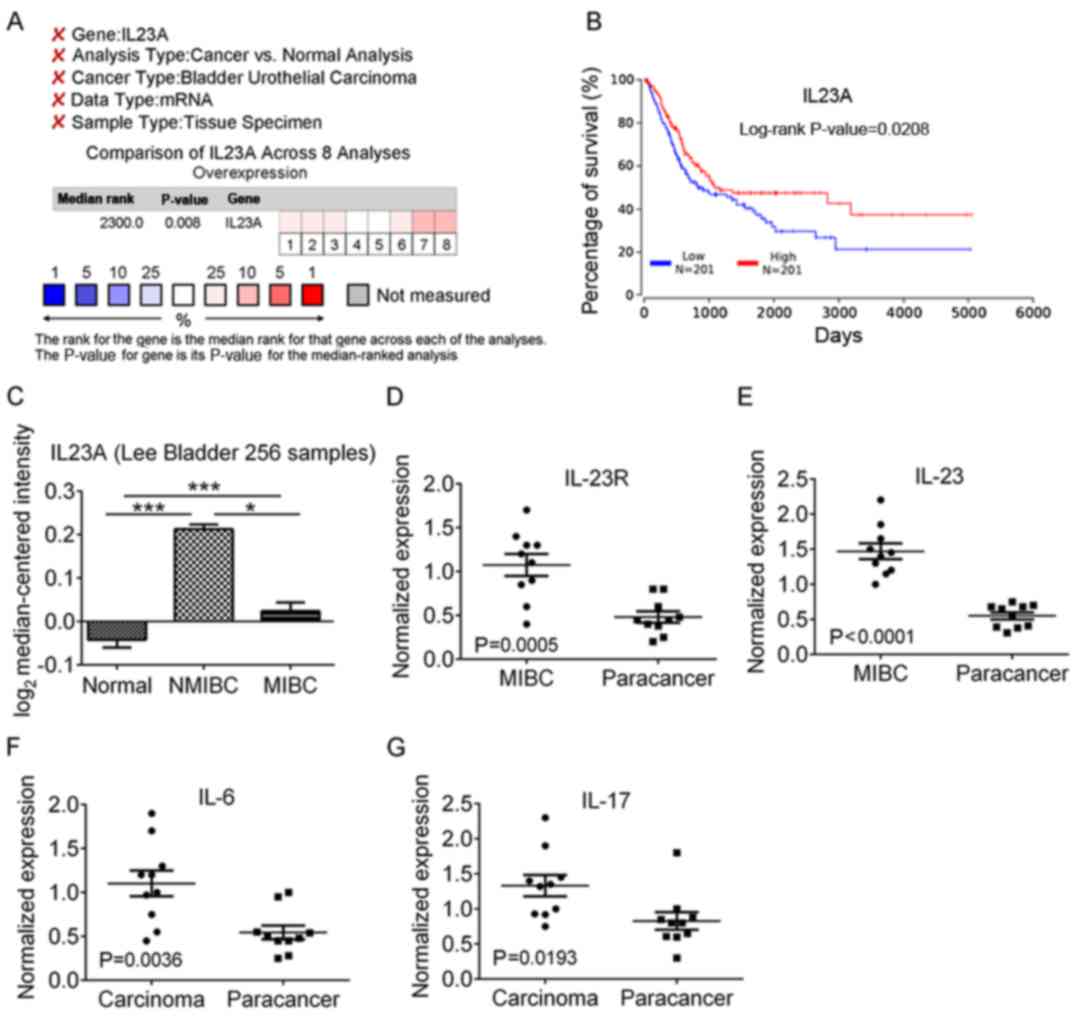 | Figure 1.mRNA expression analysis of IL23A and
related inflammatory cytokine. (A) Eight data sets of bladder
urothelial carcinoma were obtained by searching in the Oncomine
database with the key words including IL23A, Cancer vs. Normal
Analysis, Bladder Urothelial Carcinoma, mRNA and Tissue Specimen
and over-expression analysis revealed that the expression of IL23A
in cancer tissue was significantly increased in comparison with
normal bladder urothelial tissue. (B) TCGA BLCA-Bladder Urothelial
Carcinoma (Nature 2014) (16) Data
Center Kaplan-Meier plots revaled overall survival in bladder
urothelial carcinoma. In red: Patients with expression above the
median; in blue, patients with expression below the median. (C) The
expression level of IL23A was significantly higher in NMIBC and
MIBC than that in normal bladder tissue, and the expression of
IL23A in MIBC was significantly lower than that in NMIBC. (D-G) The
expression levels of IL-23R, IL-23, IL-6 and IL-17 mRNA in the MIBC
were significantly higher than those in the adjacent tissues. Lee,
represents the Lee Bladder dataset in Oncomine database (19); NMIBC, non-muscle invasive bladder
urothelial carcinoma; MIBC, muscle invasive bladder urothelial
carcinoma. *P<0.05, ***P<0.001. |
Using Oncomine Cancer Microarray
Database and The Cancer Genome Atlas (TCGA) to analyze the
expression of IL23A in bladder urothelial carcinoma
To further validate the accuracy of the results, we
used the Oncomine database to carry out bioinformatics analysis of
IL23A gene expression in bladder urothelial carcinoma, and used
normal bladder mucosa tissues and/or adjacent tissues as a control.
To reduce the error, we set the filter keywords before the
meta-analysis (Fig. 1A), with
threshold parameters for gene expression analysis at
P<1×10−4, fold change >2, and median gene rank in
the top 10%. We found IL23A gene expression in tissue samples of
424 patients in 8 data sets of 4 research projects, and the
overexpression analysis results indicated P=0.008 (Fig. 1A). The two most significant data
sets were infiltrating bladder urothelial carcinoma vs. normal
(P=4.79×10−4) (19) and
infiltrating bladder urothelial carcinoma vs. normal
(P=1.78×10−7) (20). We
selected data with the largest sample, which included 256 cases, to
analyze the expression difference, and the results revealed that
IL23A in NMIBC (also known as superficial tumor) and MIBC was
significantly increased and was higher in NMIBC than in MIBC
(Fig. 1C).
To further analyze the role of IL23A in bladder
cancer, we used TCGA data to analyze the relationship between IL23A
expression and clinicopathological characteristics (Table II) (21), since we were unable to obtain all of
the Oncomine data. The results indicated that a high expression
level of IL23A was correlated with a low clinical stage and good
overall survival, while a low expression level of IL23A was
correlated with a high clinical stage and poor overall survival.
However, the expression level of IL23A was not significantly
correlated with age, sex, tumor stage, grade or metastasis.
| Table II.Clinical association between the
IL23A expression and the clinicopathological variables in bladder
cancer patients. |
Table II.
Clinical association between the
IL23A expression and the clinicopathological variables in bladder
cancer patients.
|
|
| IL23A |
|
|---|
|
|
|
|
|
|---|
| Variable | No. of
patients | High
expression | Low expression | χ2 test
P-value |
|---|
| Age (years) |
|
|
| 0.112 |
|
>60 | 300 | 65 | 235 |
|
|
≤60 | 107 | 32 | 75 |
|
| Sex |
|
|
| 0.597 |
|
Male | 300 | 69 | 231 |
|
|
Female | 107 | 28 | 79 |
|
| Smoking status |
|
|
| 0.596 |
|
Smoking | 394 | 95 | 299 |
|
|
Non-smoking | 14 | 2 | 12 |
|
| T
classification |
|
|
| 0.483 |
|
T1+2 | 128 | 34 | 94 |
|
|
T3+4 | 41 | 8 | 33 |
|
| Lymph node
stage |
|
|
| 0.83 |
|
N0+1 | 282 | 70 | 212 |
|
|
N2+3 | 83 | 19 | 64 |
|
| M
classification |
|
|
| 0.185 |
|
M0 | 195 | 52 | 143 |
|
|
M1 | 209 | 43 | 166 |
|
| Metastasis |
|
|
| 0.794 |
| No | 147 | 31 | 116 |
|
|
Yes | 55 | 10 | 45 |
|
| Clinical stage |
|
|
| 0.017 |
|
I–II | 132 | 41 | 91 |
|
|
III–IV | 273 | 54 | 219 |
|
Low concentration of IL-23 promotes
T24 cell proliferation, migration, and invasion, and high
concentration of IL-23 inhibits T24 cell proliferation, migration
and invasion
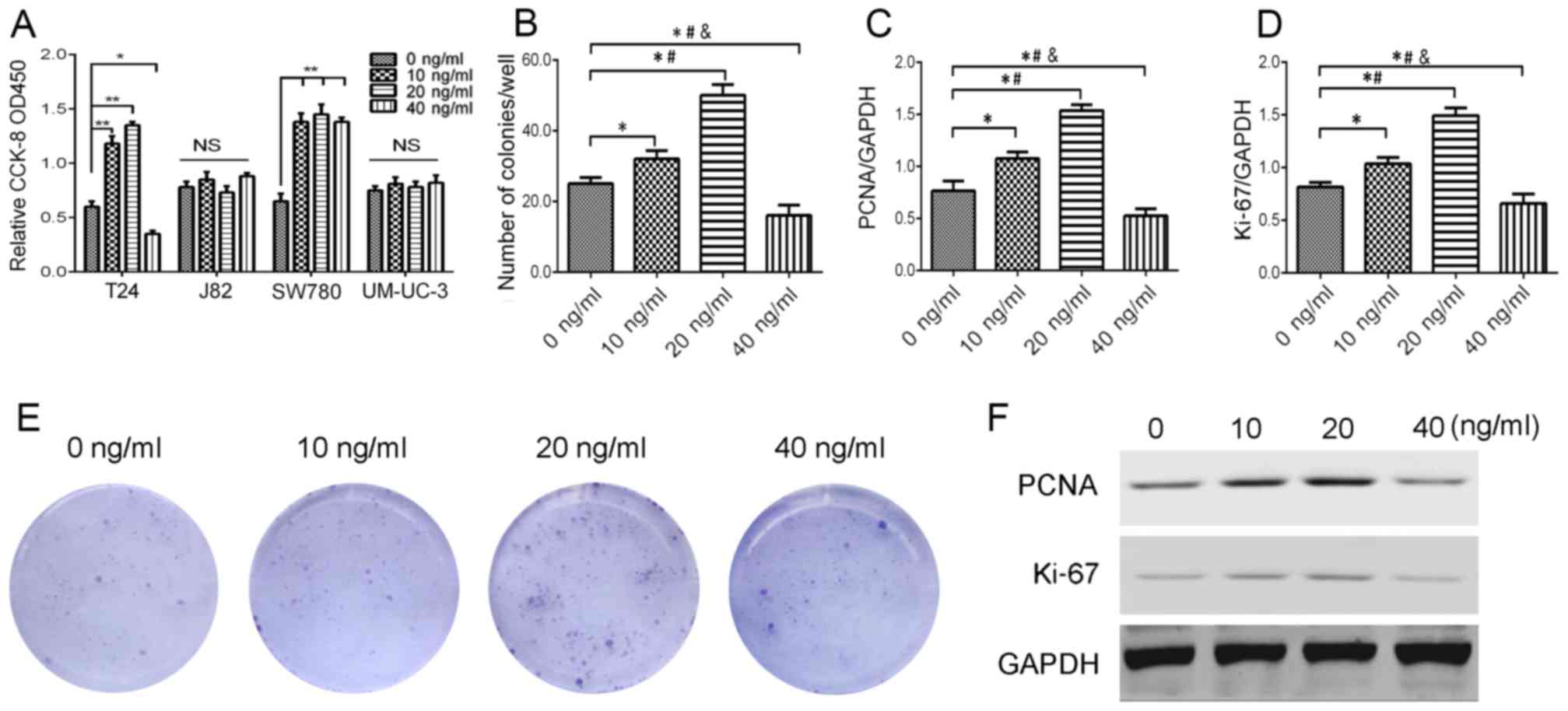
After IL-23 treatment (0, 10, 20 and 40 ng/ml) on
bladder urothelial carcinoma for 48 h, a CCK-8 assay revealed that
a low concentration of IL-23 (20 ng/ml) significantly promoted the
proliferation of T24 cells (P<0.05), while IL-23 (40 ng/ml)
inhibited its proliferation (P<0.05); 10–20 ng/ml IL-23 promoted
the proliferation of SW780 cells, which had no concentration
dependence; IL-23 had no significant proliferative effect on J82
cells and UM-UC-3 cells (Fig. 2A).
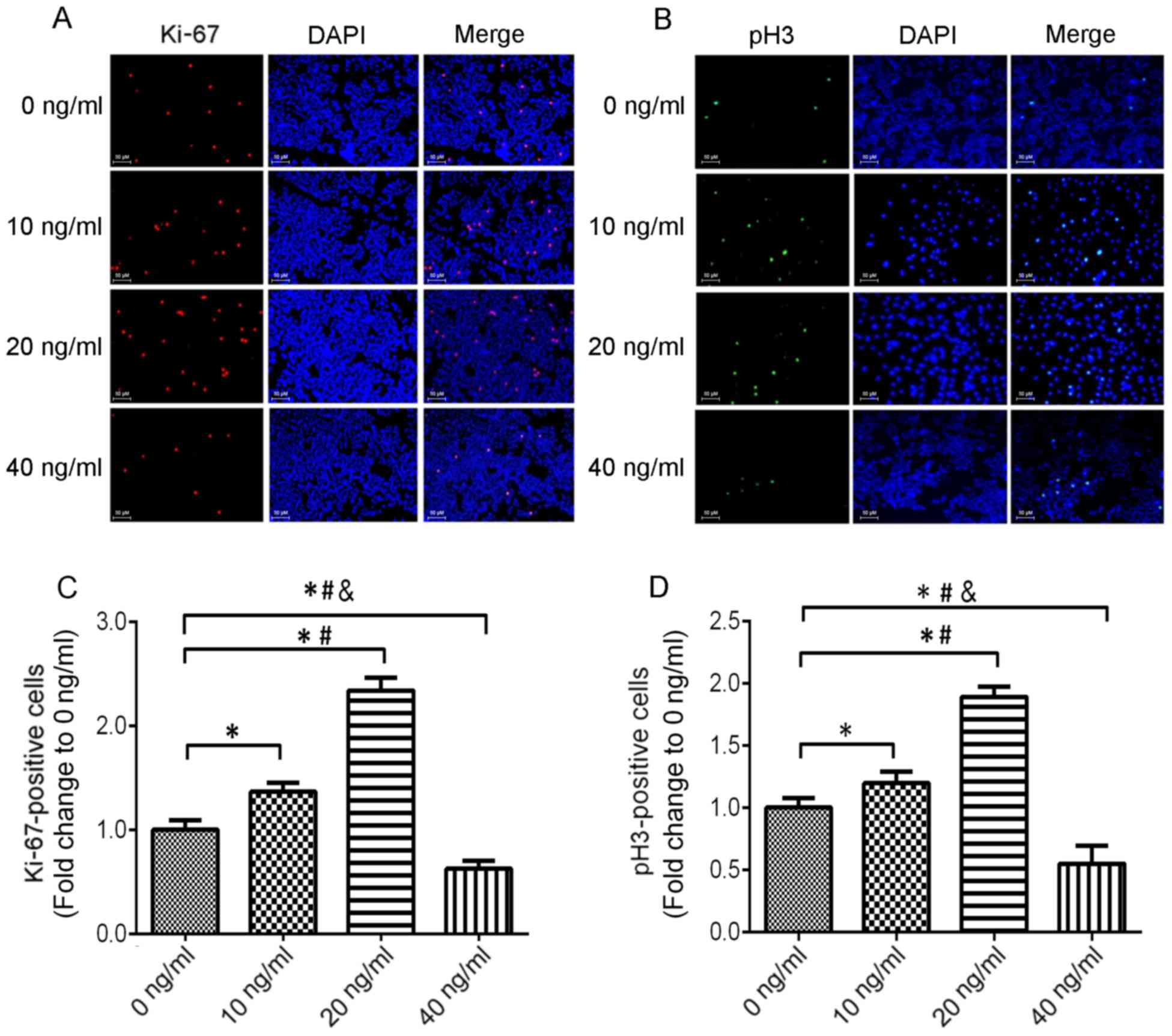
Colony formation assay (Fig. 2B and
E) and immunofluorescence (Fig.
3) were used to detect the number of Ki-67 and pH3 positive
cells to verify the proliferation effect of IL-23 on T24 cells,
which produced similar results with the CCK-8 assay. Concurrently,
we observed the effect of different concentrations of IL-23 on
Ki-67 and PCNA protein expression in T24 cells, revealing that 10
and 20 ng/ml of IL-23 promoted the upregulation of Ki-67 and PCNA
protein, while a high concentration of IL-23 (40 ng/ml) played an
inhibitory role (Fig. 2C, D and F).
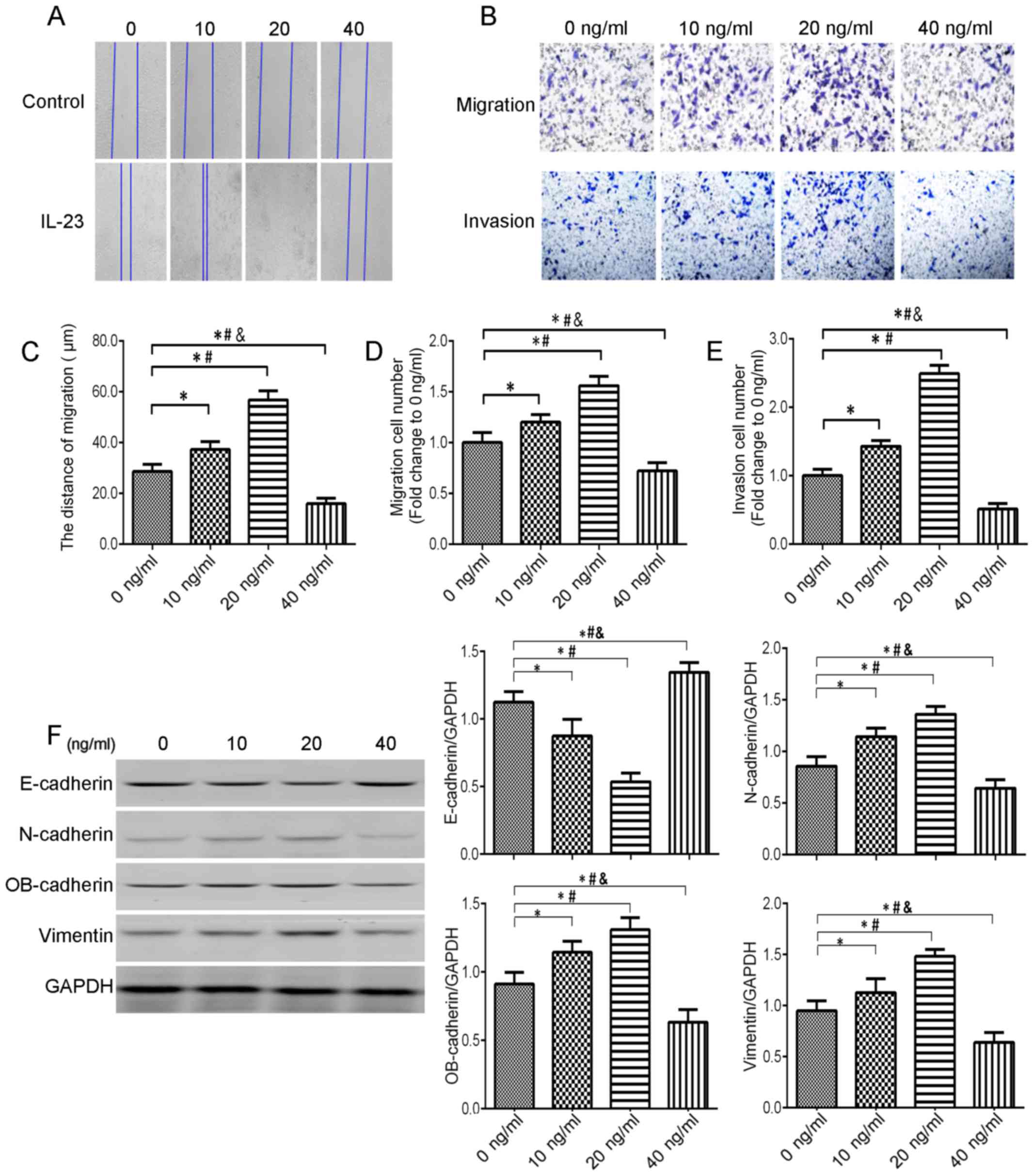
The relationship between IL-23 and migration and invasion of
bladder urothelial carcinoma cells has been studied in
vitro. In a wound-healing assay, low concentrations of IL-23
(10 and 20 ng/ml) promoted T24 cell migration (P<0.05) and a
high concentration of IL-23 inhibited T24 cell migration (P
<0.05) (Fig. 4A and C). A
Transwell assay further demonstrated that low concentrations of
IL-23 (10 and 20 ng/ml) promoted T24 migration and invasion,
whereas a high concentration of IL-23 (40 ng/ml) played an
inhibitory role (Fig. 4B, D and
E).
Low concentration of IL-23 upregulates
the expression of EMT-related proteins in T24 cells, but high
concentration of IL-23 downregulates the expression of EMT-related
proteins in T24 cells
Epithelial-mesenchymal transition (EMT) refers to
the transformation of epithelial cells into interstitial cells
after stimulation by external factors. In this process, epithelial
cells lose polarity and the adhesion connection between cells,
increase the migration and athletic ability of cells, and obtain
the interstitial phenotype (22).
It is characterized by spindle-shaped morphology, decreased cell
adhesion, and increased migration and motor capacity, which is
closely related to tumor metastasis. It is also characterized by
downregulation of the expression of E-cadherin, and upregulation of
the expression of N-cadherin, vimentin and OB-cadherin
(cadherin-11) (23). Studies have
reported that inflammatory factors can activate the EMT pathway. To
determine whether IL-23 can promote T24 cell EMT, we detected the
expression of EMT-related proteins in T24 cells under different
concentrations of IL-23. The expression of E-cadherin was
downregulated after treatment with IL-23 (10 and 20 ng/ml) for 48
h, and the expression of N-cadherin, OB-cadherin and vimentin was
upregulated; however, IL-23 (40 ng/ml) increased the expression of
E-cadherin, and decreased the expression of N-cadherin, OB-cadherin
and vimentin (Fig. 4F).
Discussion
Chronic inflammation is an important factor in
promoting tumor development. Due to the heterogeneity of the tumor
microenvironment, the role of chronic inflammation in different
tumor tissues is still widely controversial (24). In recent years, the role of the
IL-12 cytokine family in chronic inflammation and tumor progression
has been widely reported. The IL-12/STAT4 signaling pathway
activates antitumor immune response, which mainly secretes IFN-γ
(8). The IL-23/STAT3 signaling
pathway activates the tumor-associated inflammatory response, which
mainly secretes IL-17 and promotes tumor cell immune escape
(11). It was revealed that
exogenously overexpressed IL-23 inhibited tumorigenesis and
development, whereas endogenously low levels of IL-23 promoted
tumorigenesis and progression, while anti-IL-23 monoclonal antibody
can inhibit tumor growth and metastasis (25). IL-23 appears to play a double
opposing role in tumor development and progression, but there are
few studies on the direct effect of IL-23 on tumor cells.
In the present study, we reported that the level of
IL-23 expression in human bladder urothelial carcinoma was
significantly higher than that in adjacent tissues as determined by
immunohistochemistry. We further demonstrated these results by
detecting mRNA expression levels and using the Oncomine database.
IL-23 was significantly increased in most tumor tissues from
different organs and was not expressed in the same specimen of
normal tissues, which suggested that IL-23 was tumor-specifically
upregulated and may not be the result of mutations in susceptible
genes (26). The effect of IL-23 on
tumorigenesis was first demonstrated in IL-23p19-deficient mouse
models (27), which showed a
significant inhibitory effect on 7,12-dimethylbenz[a]anthracene
(DMBA)/12-O-tetradecanoyl-phorbol acetate (TPA)-induced
dermal papilloma. Another study also demonstrated that
IL-23p19-deficient mice were resistant to DMBA/TPA-induced skin
papilloma and methylcholanthrene (MCA)-induced murine fibrosarcoma
(12). IL-23, IL-17A and IL-6 in
peripheral blood of patients with bladder cancer were significantly
higher than those in normal controls, and Th17 cells were
significantly increased in bladder cancer tissues (28). It has also been reported that IL-23
can enhance the motility of tumor cells and activate the NF-κB/p65
signaling pathway to upregulate the level of MMP9 in tumor cells
(29). In addition, the expression
of IL-23 in patients with primary hepatocellular carcinoma was
strongly correlated with IL-17A and MMP9. Similarly, IL-23 promoted
the proliferation of human oral squamous cell carcinoma in patients
with positive IL-23R by activating the NF-κB/p65 signaling pathway
(30). This study also indicated
that different concentrations of IL-23 play different roles in T24
cells. A relatively low concentration of IL-23 promoted T24 cell
proliferation, migration, invasion and EMT transformation, while a
relatively high concentration of IL-23 played the opposite roles.
The TCGA data analysis also revealed that low expression levels of
IL-23 were associated with higher clinical staging and poor
clinical outcomes, while relatively high levels of IL-23 had the
opposite effects.
In contrast to the results of this study, some
results indicated that IL-23 only has a role in inhibiting tumor
development (25). For example,
mouse tumor cell lines expressing IL-23 can inhibit tumor growth
in vivo (31); the use of
adenovirus expressing IL-23 in established tumor models exhibited
antitumor effects (32). It has
also been reported that IL-23 can significantly enhance its
antitumor effect only after adoptive transfer of peptide vaccine or
antigen-specific T cells (33). The
drawback of the aforementioned in vivo experiment is that it
cannot accurately explain whether IL-23 acts directly on tumor
cells and how to regulate its biological processes. Some studies
are consistent with our findings. It has been reported that
endogenously low concentrations of IL-23 promote a tumor
microenvironment characterized by immunosuppression and promote
tumor progression, whereas anti-IL-23 monoclonal antibody inhibits
tumor growth and metastasis (34);
exogenous overexpression of IL-23 has been reported to manifest
antitumor effects (32). However,
studies about the direct role IL-23 on tumor cells are rare, and
the results of different studies have been contradictory, which may
be related to tumor cell heterogeneity, IL-23 dose and different
tumor microenvironments.
The evidence concerning IL-23 in the promotion of
tumorigenesis appears to be contradictory, but it is important to
note that it is difficult to assess whether the IL-23 used in the
study is fully compliant with the physiologic dose, and these
studies may not actually reflect the tumor regulation role of the
host endogenous IL-23 in the natural state. Furthermore, the
experimental results of different types of tumors are also
different, and it is worth noting whether IL-23R is expressed.
Studies on IL-23R-positive human lung cancer cell lines have also
reported that low concentrations of IL-23 promote tumor cell
proliferation while high concentrations of IL-23 have the opposite
effect (35). Hence, we should
carefully evaluate the study results that suggest that IL-23 can
inhibit tumorigenesis.
It is clear that the expression level of IL-23 in
the tumor microenvironment determines the cancer-promoting or
tumor-suppressing characteristics of IL-23. Functional IL-23R is
composed of IL-23R and IL-12Rβ1, the IL-23R downstream signaling
molecule STAT3 promotes the secretion of IL-17-based immune
response, while IL-12Rβ1 downstream signaling molecule STAT4
promotes Th1-type immune response. IL-12 and IL-23 belong to IL-12
family of cytokines, STAT4 is a key transcription factor of the
IL-12-specific signaling pathway, and STAT3 is a key transcription
factor of the IL-23-related signaling pathway. We found the
following phenomenon through this study: Low concentrations of
IL-23 promoted the progression of bladder tumors, while high
concentrations of IL-23 played an inhibitory role. The possible
mechanisms are as follows: When IL-23 concentration is low, it is
integrated with IL-23r which has stronger affinity, activates the
STAT3 signaling pathway and promotes tumor progression.
Transcription factor STAT3 activation has been reported in multiple
tumor studies and its activation is associated with cell
proliferation, survival, pro-angiogenesis and immune escape
(27,36,37).
When the concentration of IL-23 is high, it not only binds to
IL-23r, but also to IL-12Rβ1, activating the STAT4 signaling
pathway and exerting a similar effect as IL-12. The STAT4 signaling
pathway promotes the secretion of γ-interferon by activating NK
cells and Th1-type immune responses, and activates cytotoxic T
lymphocytes and promotes their proliferation, thereby exerting
antitumor immune effects (8,25,36).
From this point of view, the antitumor effect of high
concentrations of IL-23 on the IL-12Rβ1/STAT4 signaling pathway is
worthy of further study.
In general, the mechanism of IL-23 in regulating
tumorigenesis and tumor immunity remains unclear. Related studies
are still controversial. We believe that IL-23 plays a dual role in
the progression of bladder cancer. In a future study, the
properties of IL-23 in the microenvironment of bladder tumor, the
mechanism of its secretion and the functional mechanism of IL-23 in
tumor cells should be further elucidated.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
PW and ZS conceived and designed the study. PW
participated in every step of the specific experiment, and was also
the writer of this manuscript. ZS provided the overall idea of the
study and controlled the quality of all the work throughout the
entire process. YZ and JZ performed the experiments, analyzed the
results, conducted statistical analysis of data and also wrote the
manuscript. JJ provided practical advice and guidance, and proposed
many feasible solutions. FJ reviewed, edited the manuscript and
provided the assistance with clinical case selection and
statistical analyses. All authors read and approved the manuscript
and agree to be accountable for all aspects of the research in
ensuring that the accuracy or integrity of any part of the work are
appropriately investigated and resolved.
Ethics approval and consent to
participate
All experimental protocols were approved by the
Institutional Review Board of the Deparment of Laboratory Animal
Science of Daping Hospital (Chongqing, China).
Patient consent for publication
Not applicable.
Competing interests
The authors state that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
IL
|
interleukin
|
|
NMIBC
|
non-muscle invasive bladder urothelial
carcinoma
|
|
MIBC
|
muscle invasive bladder urothelial
carcinoma
|
|
EMT
|
epithelial mesenchymal transition
|
|
TCGA
|
The Cancer Genome Atlas
|
|
IL-23R
|
IL-23 receptor
|
|
DCs
|
dendritic cells
|
|
IFN-γ
|
interferon-γ
|
|
pH3
|
phospho-histone H3
|
References
|
1
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China,
2015. CA Cancer J Clin. 66:115–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Rodriguez Martinez RH, Rueda Buisan O and
Ibarz L: Bladder cancer: Present and future. Med Clin. 149:449–455.
2017.
|
|
3
|
Kaufman DS, Shipley WU and Feldman AS:
Bladder cancer. Lancet. 374:239–249. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Thompson DB, Siref LE, Feloney MP, Hauke
RJ and Agrawal DK: Immunological basis in the pathogenesis and
treatment of bladder cancer. Expert Rev Clin Immunol. 11:265–279.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Croxford AL, Mair F and Becher B: IL-23:
One cytokine in control of autoimmunity. Eur J Immunol.
42:2263–2273. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wang P, Yang B, Zhou B, Zhang J, Li S,
Jiang J, Sun Z and Jin F: Distribution and expression profiles of
dendritic cell subpopulations in human bladder cancer. Int J Clin
Exp Pathol. 9:7180–7187. 2016.
|
|
7
|
Segura E, Touzot M, Bohineust A, Cappuccio
A, Chiocchia G, Hosmalin A, Dalod M, Soumelis V and Amigorena S:
Human inflammatory dendritic cells induce Th17 cell
differentiation. Immunity. 38:336–348. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Croxford AL, Kulig P and Becher B:
IL-12-and IL-23 in health and disease. Cytokine Growth Factor Rev.
25:415–421. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Gagliani N, Hu B, Huber S, Elinav E and
Flavell RA: The fire within: Microbes inflame tumors. Cell.
157:776–783. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tugues S, Burkhard SH, Ohs I, Vrohlings M,
Nussbaum K, Berg Vom J, Kulig P and Becher B: New insights into
IL-12-mediated tumor suppression. Cell Death Differ. 22:237–246.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang K and Karin M: The IL-23 to IL-17
cascade inflammation-related cancers. Clin Exp Rheumatol. 33 4
Suppl 92:S87–S90. 2015.PubMed/NCBI
|
|
12
|
Teng MW, Andrews DM, McLaughlin N, von
Scheidt B, Ngiow SF, Möller A, Hill GR, Iwakura Y, Oft M and Smyth
MJ: IL-23 suppresses innate immune response independently of IL-17A
during carcinogenesis and metastasis. Proc Natl Acad Sci USA.
107:8328–8333. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sobin L, Gospodarowicz M and Wittekind C:
TNM classification of malignant tumors., in U1CC 1nternational
Union Against Cancer. Wiley-Blackwell. 262–265. 2009.
|
|
14
|
Kindelan Alvarez J, Campos Hernández JP,
López Beltrán A and Requena Tapia MJ: The 2004 WHO classification
of bladder tumors: A summary and commentary. Actas Urol Esp.
31:978–988. 2007.(In Spanish). PubMed/NCBI
|
|
15
|
Rhodes DR, Yu J, Shanker K, Deshpande N,
Varambally R, Ghosh D, Barrette T, Pandey A and Chinnaiyan AM:
ONCOMINE: A cancer microarray database and integrated data-mining
platform. Neoplasia. 6:1–6. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Cancer Genome Atlas Research Network:
Comprehensive molecular characterization of urothelial bladder
carcinoma. Nature. 507:315–522. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2−ΔΔCT method. Methods. 25:402–408. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Qian X, Gu L, Ning H, Zhang Y, Hsueh EC,
Fu M, Hu X, Wei L, Hoft DF and Liu J: Increased Th17 cells in the
tumor microenvironment is mediated by IL-23 via tumor-secreted
prostaglandin E2. J Immunol. 190:5894–5902. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lee JS, Leem SH, Lee SY, Kim SC, Park ES,
Kim SB, Kim SK, Kim YJ, Kim WJ and Chu IS: Expression signature of
E2F1 and its associated genes predict superficial to invasive
progression of bladder tumors. J Clin Oncol. 28:2660–2667. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sanchez-Carbayo M, Socci ND, Lozano J,
Saint F and Cordon-Cardo C: Defining molecular profiles of poor
outcome in patients with invasive bladder cancer using
oligonucleotide microarrays. J Clin Oncol. 24:778–789. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Cancer Genome Atlas Research, Network:
Comprehensive genomic characterization defines human glioblastoma
genes and core pathways. Nature. 455:1061–1068. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Singh M, Yelle N, Venugopal C and Singh
SK: EMT: Mechanisms and therapeutic implications. Pharmacol Ther.
182:80–94. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Dominguez C, David JM and Palena C:
Epithelial-mesenchymal transition and inflammation at the site of
the primary tumor. Semin Cancer Biol. 47:177–184. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kanda Y, Osaki M and Okada F:
Chemopreventive strategies for inflammation-related carcinogenesis:
Current status and future direction. Int J Mol Sci. 18:pii:
E8672017. View Article : Google Scholar
|
|
25
|
Ngiow SF, Teng MW and Smyth MJ: A balance
of interleukin-12 and −23 in cancer. Trends Immunol. 34:548–555.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Langowski JL, Kastelein RA and Oft M:
Swords into plowshares: IL-23 repurposes tumor immune surveillance.
Trends Immunol. 28:207–212. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Langowski JL, Zhang X, Wu L, Mattson JD,
Chen T, Smith K, Basham B, McClanahan T, Kastelein RA and Oft M:
IL-23 promotes tumour incidence and growth. Nature. 442:461–465.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chugh S, Anand V, Swaroop L, Sharma M,
Seth A and Sharma A: Involvement of Th17 cells in patients of
urothelial carcinoma of bladder. Hum Immunol. 74:1258–1262. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li J, Lau G, Chen L, Yuan YF, Huang J, Luk
JM, Xie D and Guan XY: Interleukin 23 promotes hepatocellular
carcinoma metastasis via NF-kappa B induced matrix
metalloproteinase 9 expression. PLoS One. 7:e462642012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Fukuda M, Ehara M, Suzuki S, Ohmori Y and
Sakashita H: IL-23 promotes growth and proliferation in human
squamous cell carcinoma of the oral cavity. Int J Oncol.
36:1355–1365. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hu P, Hu HD, Chen M, Peng ML, Tang L, Tang
KF, Matsui M, Belladonna ML, Yoshimoto T, Zhang DZ, et al:
Expression of interleukins-23 and 27 leads to successful gene
therapy of hepatocellular carcinoma. Mol Immunol. 46:1654–1662.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Reay J, Gambotto A and Robbins PD: The
antitumor effects of adenoviral-mediated, intratumoral delivery of
interleukin 23 require endogenous IL-12. Cancer Gene Ther.
19:135–143. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Jin HT, Youn JI, Choi SY, Seo SH, Park SH,
Song MY, Yang SH and Sung YC: Adenovirus-mediated gene transfer of
interleukin-23 shows prophylactic but not therapeutic antitumor
effects. Cancer Gene Ther. 15:693–702. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
von Scheidt B, Leung PS, Yong MC, Zhang Y,
Towne JE, Smyth MJ and Teng MW: Combined anti-CD40 and anti-IL-23
monoclonal antibody therapy effectively suppresses tumor growth and
metastases. Cancer Res. 74:2412–2421. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Li J, Zhang L, Zhang J, Wei Y, Li K, Huang
L, Zhang S, Gao B, Wang X and Lin P: Interleukin 23 regulates
proliferation of lung cancer cells in a concentration-dependent way
in association with the interleukin-23 receptor. Carcinogenesis.
34:658–666. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kortylewski M, Xin H, Kujawski M, Lee H,
Liu Y, Harris T, Drake C, Pardoll D and Yu H: Regulation of the
IL-23 and IL-12 balance by Stat3 signaling in the tumor
microenvironment. Cancer Cell. 15:114–123. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Stewart CA and Trinchieri G: Reinforcing
suppression using regulators: A new link between STAT3, IL-23, and
Tregs in tumor immunosuppression. Cancer Cell. 15:81–83. 2009.
View Article : Google Scholar : PubMed/NCBI
|