Introduction
Prostate cancer (PCa) is a heterogeneous,
multifaceted and biologically complex disease. Changes in the
cancer genome and epigenome have been extensively studied in recent
years using innovative high throughput methods. Projects aimed at
accelerating the expansion of knowledge concerning the genetic
landscape of cancer, including PCa, were launched with various
platforms based on next-generation sequencing (NGS) and microarrays
to discover molecular aberrations at the DNA, RNA, protein and
epigenetic levels. The main and first large-scale cancer genomic
project was The Cancer Genome Atlas (TCGA) (https://cancergenome.nih.gov/). A pilot study of TCGA
started approximately 10 years ago to discover major genetic
alterations in large cohorts of selected tumors, i.e. brain, lung,
ovarian cancer. Phase II of the project was expanded to over 30
human tumors including prostate adenocarcinoma (1). The comprehensive characterization of
333 primary prostate cancers in the TCGA Network revealed novel
molecular features (1). Methylation
of CpG islands in gene promoters is the main epigenetic mechanism
for gene expression silencing. Aberrant methylation pattern, e.g.,
increased methylation frequency of tumor-suppressor genes, is a
common molecular feature of the majority of human cancers including
prostate cancer (2). Genes that
protect cells from neoplastic transformation are not only known as
tumor suppressors, but often their products function as tumor cell
invasion factors and are involved in cell metabolism and DNA
repair. Failure of these functions leads to carcinogenesis
(3).
The necessity for precise prostate cancer
diagnostics and disease prognosis encourages the search for novel
biomarkers and basic scientific research. Microarray-based gene
signatures are used for cancer diagnostics, tumor classification
and prognosis, and prediction of response to therapies (4,5). There
are several types of signatures that have been evaluated for cancer
diagnostics: Based on gene expression-in the breast (5), colon (6) and lung cancers (7); based on methylation analysis-in colon
cancer (8); and based on miRNA
expression data (9). Profiling in
prostate cancer is still at the exploration stage.
The aim of the present study was to evaluate the
gene methylation profile using a prostate cancer cell line model
and The Human Prostate Cancer EpiTect Methyl II Signature PCR Array
designed to evaluate the following genes: APC, CAV1, CDH1,
CDKN2A, DKK3, DLC1, EDNRB, GPX3, GSTP1, MGMT, MSX1, PDLIM4, PTGS2,
RARB, RASSF1, SFRP1, SLC5A8, TIMP2, TNFRSF10D, ZNF185. For our
study, we chose prostate cancer cell lines PC3, PC3M, PC3MLN4 and
PC3MPro4 which shared the same origin but each of them demonstrated
different levels of metastatic capabilities in a mouse model of
human prostate cancer. They provided a system to associate the
level of expression with malignancy.
Oncogenesis is associated with abnormal regulation
of those genes, which are responsible for various functions in
cells, such as cell signaling, cytoskeletal architecture, cell-cell
contacts, cell motility, reorganization of the extracellular matrix
(ECM) and many other mechanisms. Changes in these processes are
important determinants of tumor invasion and metastasis. Molecular
mechanisms underlying these processes have been under evaluation in
the last few years: APC (10), CAV1 (11), CDKN2A (12), CDH1 (13), DLC1 (14), DKK3 (15), EDNRB (16), GPX3 (17), GSTP1 (18), MGMT (19), MSX1 (20) PDLIM4 (21), PTGS2 (22), RARB (23), RASSF1 (24), SFRP1 (25), SLC5A8 (26), TIMP2
(27), TNFRSF10D (28), ZNF185 (29).
For our study, we chose a prostate cancer metastasis
model (30,31) and wild-type normal skin fibroblasts
(32,33). After evaluation of methylation using
The Human Prostate Cancer EpiTect Methyl II Signature PCR Array, we
examined the expression status of the genes to confirm whether
methylation regulated them. Although several genes [for example
APC (34), GPX3
(35) PDLIM4 (21)] have been analyzed in the PC3 cell
line, this was the first study to use this qPCR method to describe
gene expression and methylation in PC3-derived cell lines (PC3M,
PC3MLN4 and PC3MPro4). Finally, gene methylation data in prostate
cancer patients derived from the TCGA project were evaluated.
Materials and methods
Cell line cultures
Prostate cancer cell lines, PC3, PC3M, PC3MLN4 and
PC3MPro4 (36), and reference human
WT fibroblast cell lines, VH10 and VH25 (32,33),
were kindly provided by Professor S. Huang and Dr A. Bialkowska,
respectively. Prostate cancer cell lines were cultured in cultured
dishes with a growth area of 100 mm2 in L-glutamine
RPMI-1640 medium (GE Healthcare Life Sciences, Marlborough, MA
USA). The fibroblast cell lines (VH10 and VH25) were cultured in
High Glucose DMEM medium (GE Healthcare Life Sciences). RPMI and
DMEM were supplemented with 10% fetal bovine serum (GE Healthcare
Life Sciences) and 1% antibiotic/antimycotic solution (GE
Healthcare Life Sciences): 100 U/ml of penicillin, 100 µg/ml of
streptomycin and 0.25 µg/ml of amphotericin B. The cells were
maintained at 37°C in a 5% CO2 atmosphere and a relative
humidity of 95%.
Methylation analysis of the cell
lines
Methylation analysis was performed using EpiTect
Methyl II PCR Array, Signature Panel (cat. no. EAHS-051Z; Qiagen,
Inc., Valencia, CA, USA) according to the manufacturer's protocol,
as follows. DNA from the PC3, PC3M, PC3MLN4 and VH10 cells was
isolated using QIAamp DNA FFPE Tissue Kit (Qiagen, Inc.) according
to the protocol, with additional incubation with RNase A. The
absence of RNA contamination was tested using agarose gel
electrophoresis. Subsequently, incubation with
methylation-sensitive (Ms), methylation-dependent (Md), and double
(Msd) restriction endonuclease was performed. After digestion,
quantitative PCR (qPCR) was performed using primer mixes
pre-dispensed into 96-wells to evaluate the methylation status of
the 20 (from 22) following genes: APC, CAV1, CDH1, CDKN2A, DKK3,
DLC1, EDNRB, GPX3, GSTP1, MGMT, MSX1, PDLIM4, PTGS2, RARB, RASSF1,
SFRP1, SLC5A8, TIMP2, TNFRSF10D and ZNF185. The
methylation status of selected gene promoters was analyzed using an
integrated Excel-based template (SA Bioscience, Qiagen). Raw
threshold cycle values of both digests along with mock digestion
values were normalized, and the percentage of un/methylated DNA was
automatically calculated using the MethylScreen™ technology
provided under license from Orion Genomics, LLC, St. Louis, MO,
USA). A heatmap was created using free on-line software-Morpheus
from Broad Institute (https://software.broadinstitute.org/morpheus/).
RNA isolation and cDNA synthesis
RNA isolation was performed in the cell cultures
reaching ~80% confluency in cultured dishes with a growth area of
100 mm2. Cells were trypsinized and centrifuged (300 ×
g, 5 min), and the cell pellet was suspended in 600 ml PBS 1X
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) and transferred
into 3 tubes in equal volumes (200 µl). Total RNA was isolated from
each cell line in triplicates with the High Pure RNA Isolation Kit
(Roche Diagnostics GmbH, Mannheim, Germany) according to the
manufacturer's protocol. RNA concentration was evaluated using
NanoDrop 1000 (Thermo Fisher Scientific, Inc., Waltham, MA, USA).
Approximately 1 µg of each RNA sample was used to synthesize
complementary DNA (cDNA) with the Transcriptor High Fidelity cDNA
Synthesis Kit (Roche Diagnostics GmbH) according to the
manufacturer's protocol with small modifications. All cDNAs were
synthesized using half of the recommended volume of the
anchored-oligo(dT)18 primer and the random hexamer
primer. Denaturation the template-primer mixture was carried out by
incubating the tube for 10 min at 65°C on a thermal cycler block,
followed by the addition of the remaining components of the reverse
transcriptase mix (RT-mix). The reverse transcription reaction was
carried out at 50°C for 30 min and 85°C for 5 min. The synthesized
cDNA was stored at −20°C until subsequent use.
cDNA purity control
The absence of genomic DNA in the cDNA samples was
tested by cDNA amplification with a set of primers localized in an
intron sequence of DNA: Forward primer (Int2F,
5′-ACATGTAATTATCATGTGAATTTATTACGA-3′) and reverse primer (Int2R,
5′-CTCAGAGCTTCAGTTATGGAGA-3′). The positive control was genomic DNA
routinely used in the laboratory in cDNA purity testing. Agarose
gel electrophoresis was performed. Lack of contamination with
genomic DNA was reflected by the lack of intron amplification
products for the cDNA samples.
Gene expression analysis
Quantitative reverse transcriptase real-time PCR
(RT-qPCR) was performed using two technologies: i) TaqMan
Probes-hydrolysis probes dual-labeled with a reporter fluorophore
and a dark quencher dye (LightCycler® 480 Probes Master,
The Universal ProbeLibrary Set, Human; both by Roche Diagnostics
GmbH) specific to target gene, and ii) a double-stranded DNA
binding dye (LightCycler 480 SYBR-Green; Roche Diagnostics GmbH).
Analyses were performed on LightCycler 2.0 and LightCycler 480
instruments (Roche Diagnostics GmbH). The PBGD (for the
probe-based assay) and hMRPL19 (for the SYBR-Green assay)
genes were used as a reference. Primers specific for the mRNA
sequences of the analyzed genes were designed using the Universal
ProbeLibrary Assay Design Center software accessible at www.universalprobelibrary.com. The
primers were designed to have intron-spanning sequences to avoid
false-positive signals from the possible residual genomic DNA.
Samples without reverse transcriptase for each cell line and
samples without RNA were used as negative controls. An amount of 2
µl of sample cDNA was added to each reaction with the PBGD
reference gene (Universal ProbeLibrary Human PBGD Gene Assay; Roche
Diagnostics GmbH). The Universal ProbeLibrary probe was
5′end-labeled with fluorescein (FAM) and 3′end-labeled with a dark
quencher dye. The UPL Reference Gene probe was labeled with
LightCycler® Yellow 555 at the 5′end and with a quencher
dye near the 3′end. Real-time PCR was performed in dual color. The
fluorescence signal was acquired in two detection channels: FAM
(530 nm) and LightCycler® Yellow 555 (610 nm). Real-time
PCR was conducted under the following conditions: one cycle at
95°C/10 min; 45 cycles of denaturation (95°C/10 sec), annealing
(60°C/30 sec) and extension (72°C/1 sec). The expression of the
second reference gene hMRPL19 was evaluated using SYBR-Green
and 1 µl of sample cDNA. PCR conditions consisted of: One cycle at
95°C/10 min; 45 cycles of denaturation (95°C/10 sec), annealing
(60°C/20 sec) and extension (72°C/5 sec); one cycle of melting
curve: 95°C/5 sec, 40°C/1 min, 97°C, according to a previous
publication (37). Relative gene
expression was calculated using the ΔΔCq method (38). Gene expression was randomly tested
in triplicates using the Universal ProbeLibrary Human GAPD Gene
Assay (Roche Diagnostics GmbH) which accounted for the third
control analysis. Any significant difference in the trends of high
or low expression of the targeted gene between PBGD and
hMRPL19 was observed.
MethHC database
The datasets for the analysis and visualization of
the methylation level of APC, DKK3, GPX3, GSTP1, MGMT, PTGS2,
RASSF1, TIMP2, TNFRSF10D and PDLIM4 in prostate
adenocarcinoma TCGA were obtained from the MethHC web base
(http://methhc.mbc.nctu.edu.tw/php/index.php) and
analyzed using tools available under the MethHC open access terms
(39).
Statistical analysis
RNA extraction and cDNA synthesis were performed in
three biological replicates for each cell line. Gene expression was
analyzed at least three times for each biological replicate, and
means were calculated from nine values. Promoter methylation status
was analyzed in three biological replicates for each cell line.
Differences in gene expression and methylation between analyzed
cell lines were determined using one-way analysis of variance
(ANOVA), followed by Tukey's HSD (honestly significant difference)
post hoc tests in the STATISTICA Software (StatSoft, Inc. Tulsa,
OK, USA). P≤0.05 and P≤0.01 were considered as statistically
significant. Statistically significant differences between the VH10
cell line and prostate cancer cell lines were marked on the figures
with asterisks. Data are presented as means with standard deviation
(SD).
Results
Methylation profile
The initial data to be established were the
methylation levels of genes important in the carcinogenesis process
in four cell lines, non-cancerous VH10 and three prostate cancer
cell lines, PC3, PC3M and PC3MLN4. The Human Prostate Cancer
EpiTect Methyl II Signature PCR Array was used to analyze the
methylation status of 20 out of 22 gene promoters. The kit is based
on DNA treatment with a methylation-sensitive and
methylation-dependent restriction enzyme followed by qPCR. It was
found that hypermethylation was the major mechanism of regulation
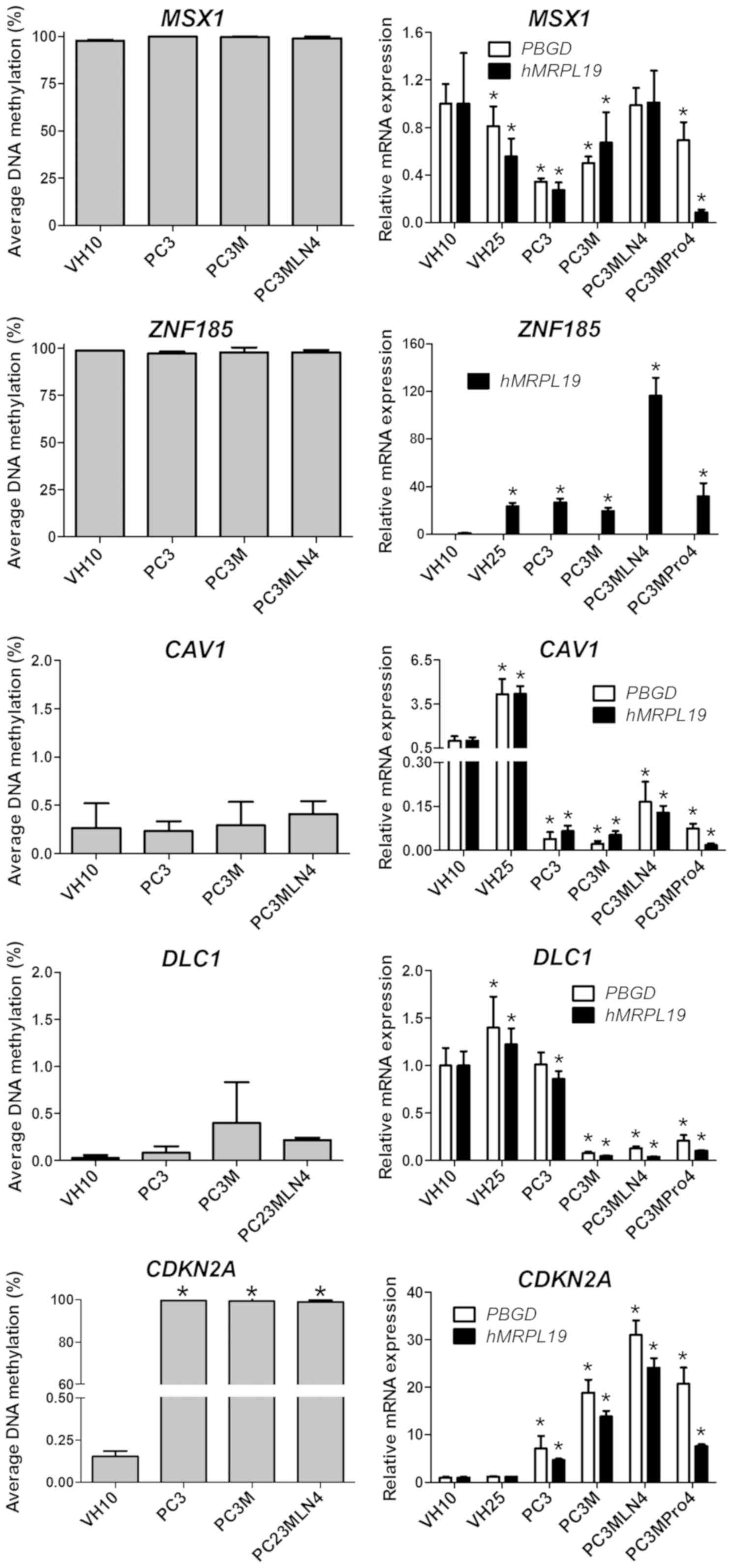
of expression of the analyzed genes (Fig. 1).
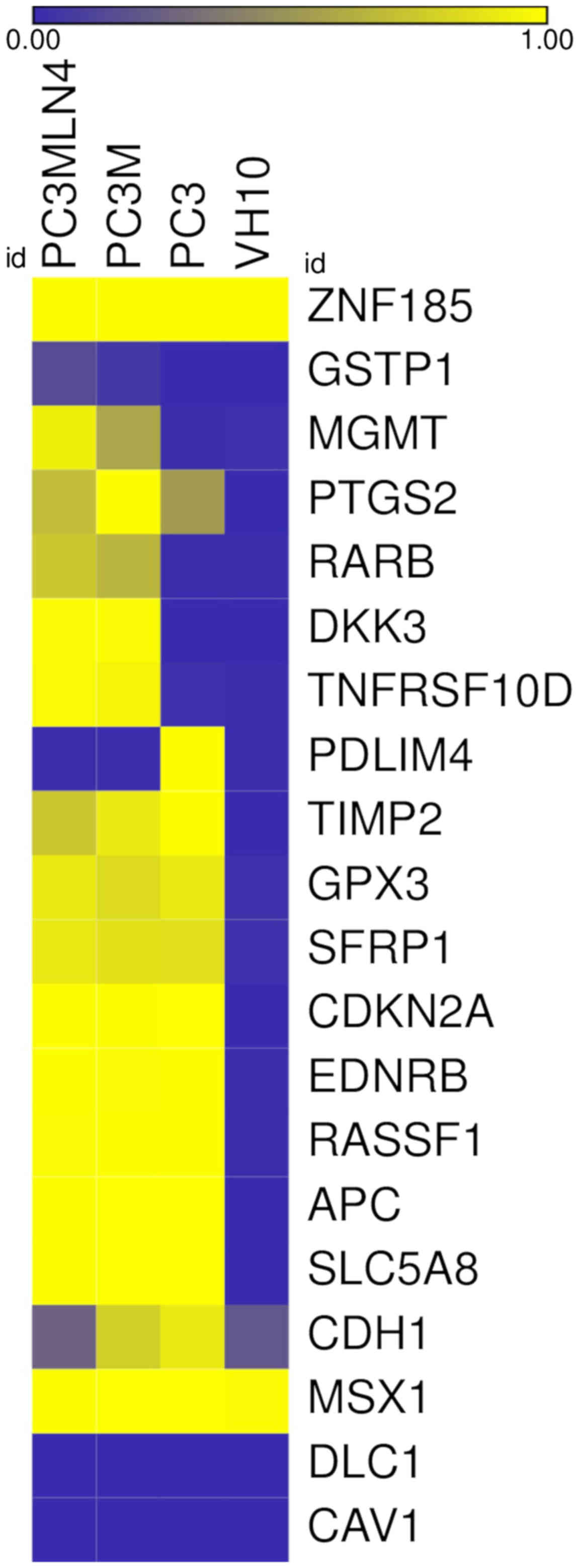 | Figure 1.Heatmap of CpG methylation in PC3,
PC3M, PC3MLN4 and VH10 cell lines. Each column represents an
average of cell line methylation data performed in triplicates;
each row represents a described gene. Methylation increases from
blue (non-methylated) to yellow (methylated). ZNF185, zinc
finger protein 185 with LIM domain; GSTP1, glutathione
S-transferase Pi 1; MGMT, O-6-methylguanine-DNA
methyltransferase; PTGS2, prostaglandin-endoperoxide
synthase 2; RARB, retinoic acid receptor β; DKK3,
Dickkopf WNT signaling pathway inhibitor 3; TNFRSF10D, TNF
receptor superfamily member 10d; PDLIM4, PDZ and LIM domain
4; TIMP2, TIMP metallopeptidase inhibitor 2; GPX3,
glutathione peroxidase 3; SFRP1, secreted frizzled related
protein 1; CDKN2A, cyclin dependent kinase inhibitor 2A;
EDNRB, endothelin receptor type B; RASSF1, Ras
association domain family member 1; APC, APC, WNT signaling
pathway regulator; SLC5A8, solute carrier family 5 member 8;
CDH1, cadherin 1; MSX1, Msh homeobox 1; DLC1,
DLC1 Rho GTPase activating protein; CAV1, caveolin 1. |
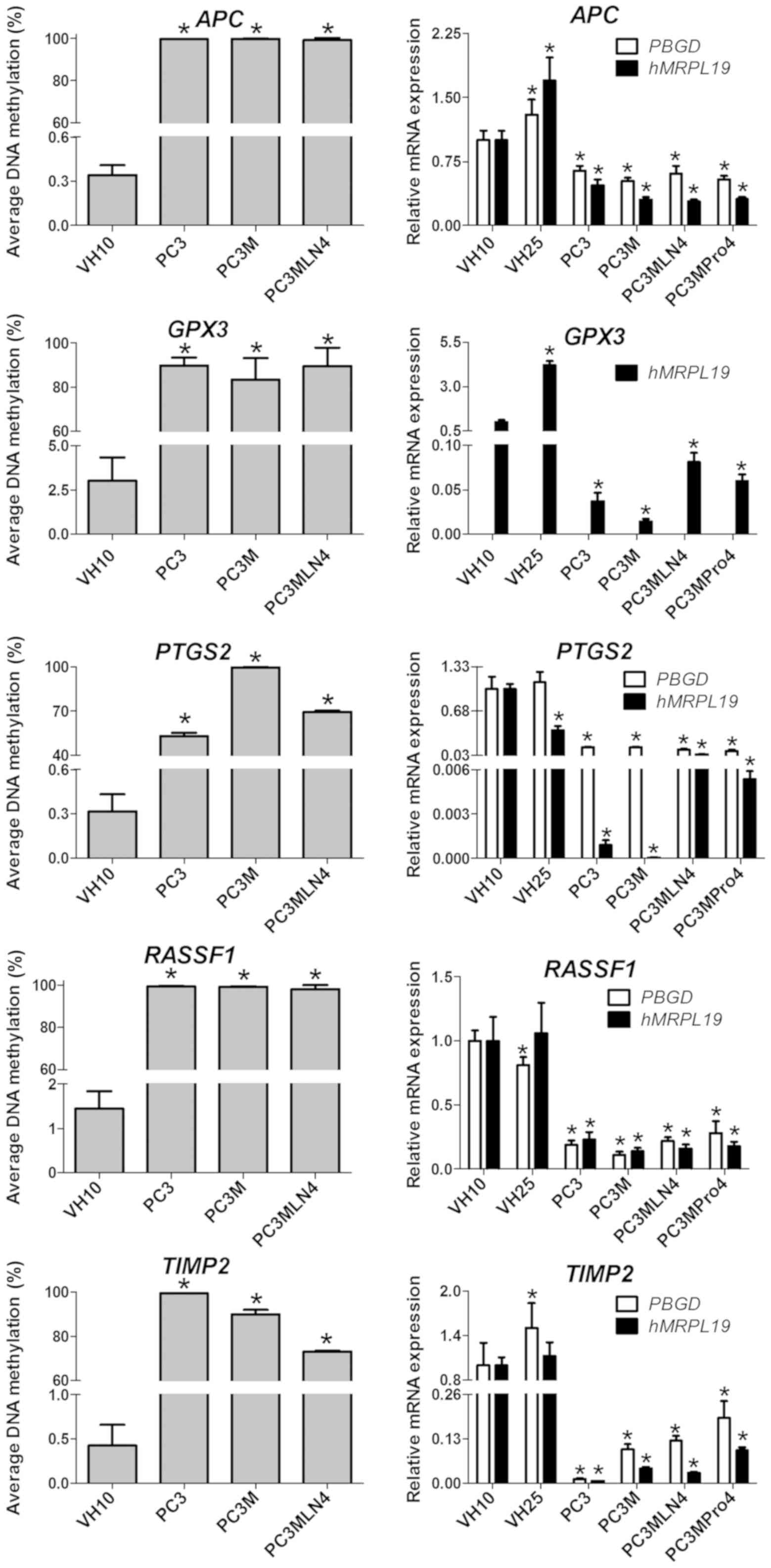
An increase in the methylation status in the
promoter regions of 9 genes in the prostate cancer cell lines
compared to that in the VH10 cells was observed: APC, CDKN2A,
EDNRB, RASSF1, SFRP1, SLC5A8, GPX3, PTGS2 and TIMP2
(Fig. 1). The methylation status
for the first 5 genes was higher and achieved statistical
significance (P<0.01) in all analyzed prostate cancer cell lines
(PC3, PC3M and PC3MLN4) compared to the VH10 cell line and was
close to 100%. GPX3 showed an increase in the methylation
level to 83% in PC3M and to 90% in PC3 and PC3MLN4 cell lines. The
methylation status of the PTGS2 gene increased to 53% in
PC3, to 69% in PC3MLN4 and to 99% in PC3M cells. The TIMP2
methylation gradually increased to 73% in PC3MLN4, to 90% in PC3M
and to 99% in PC3 cells (Fig. 1).
All these genes were hypomethylated in the VH10 cell line
(0–3%).
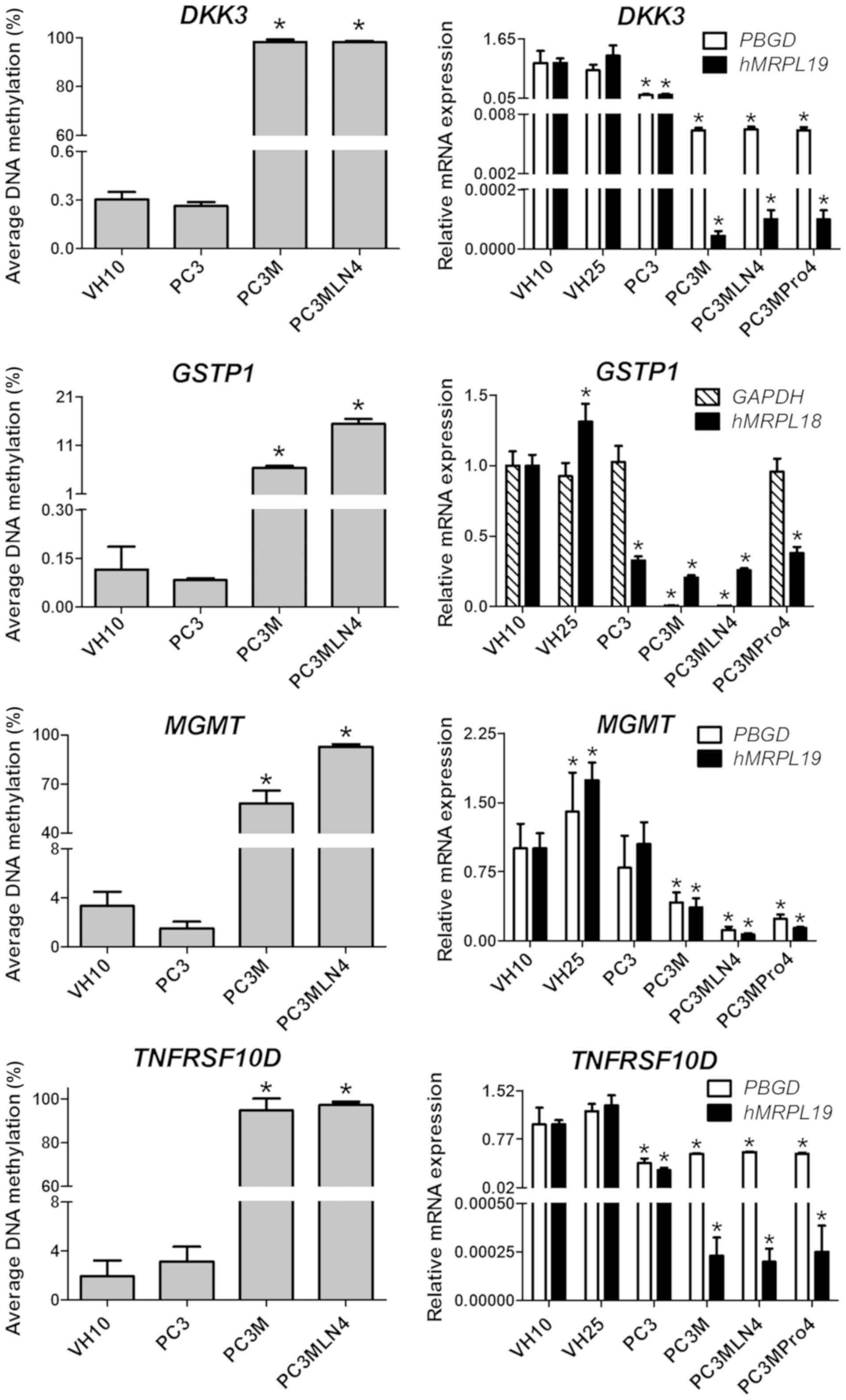
Moreover, 4 genes: DKK3, MGMT, TNFRSF10D,
RARB, were hypermethylated in PC3M and PC3MLN4 cells when
compared to the PC3 and VH10 cell lines (Fig. 1). The methylation level of these
genes in the VH10 and PC3 cells was in the range 0–3 %. The
methylation of DKK3 and TNFRSF10D in the PC3M and
PC3MLN4 cell lines increased to almost 100%. The MGMT gene
showed a methylation level of 58% in PC3M and 93% in the PC3MLN4
cells. The RARB methylation level in PC3M and PC3MLN4 cells
was 65 and 74%, respectively. For the GSTP1 gene, a much
smaller but statistically significant increase in the methylation
level in PC3M and PC3MLN4 compared to the PC3 and VH10 cell lines
(P≤0.01) was noted. The GSTP1 methylation level increased
from 0% in PC3 and VH10 to 6% in PC3M and 15% in the PC3MLN4 cell
line.
No differences were observed in the methylation
status of 4 genes: ZNF185, CAV1, MSX1 and DLC1.
ZNF185 and MSX1 were hypermethylated, while
CAV1 and DLC1 were hypomethylated in all analyzed
cell lines. The methylation level of the CDH1 gene exceeded
20% in all analyzed cell lines: VH10 (21%), PC3MLN4 (26%), PC3M
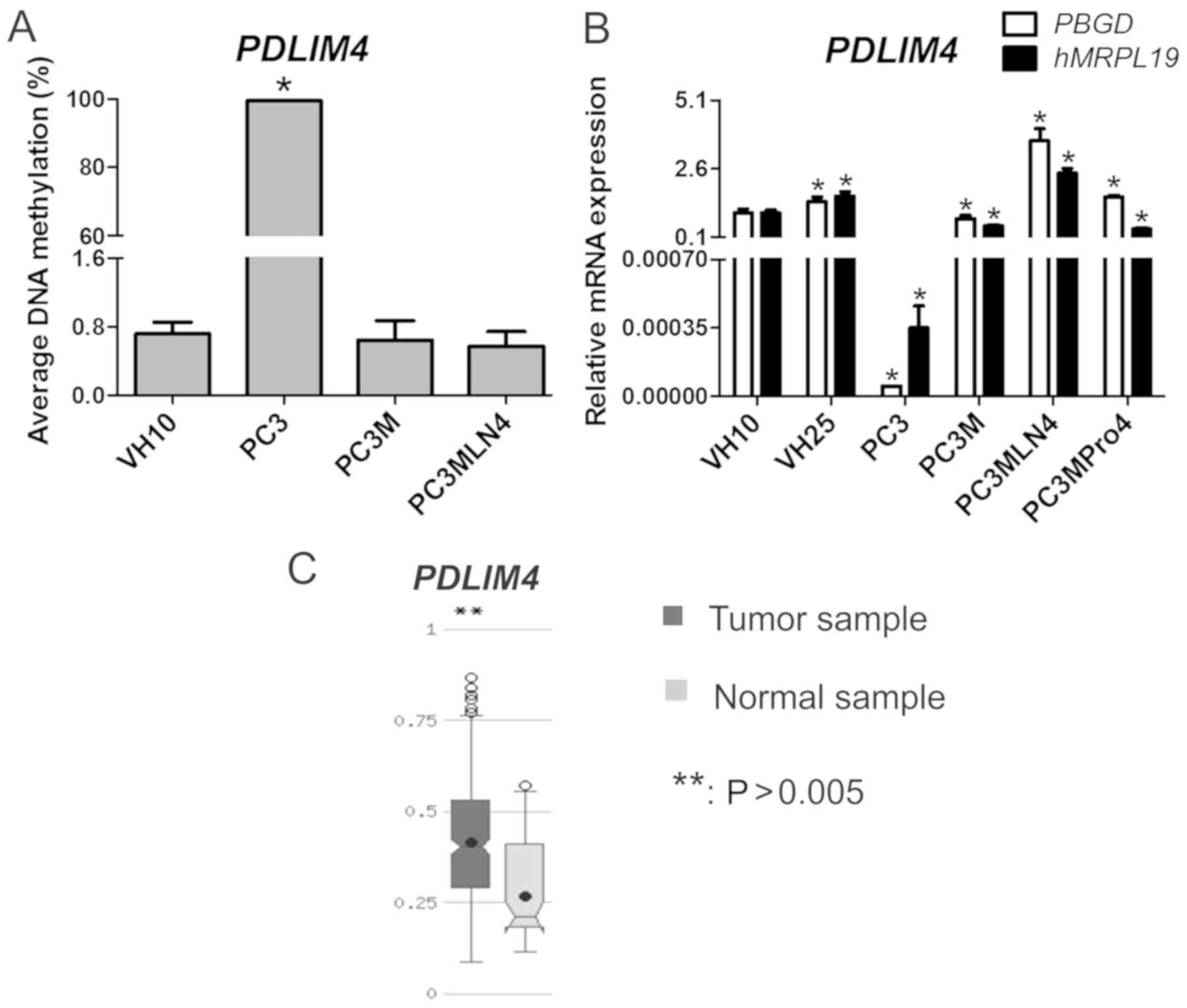
(77%) and PC3 (90%). Importantly, we found that the PDLIM4
methylation status was higher and achieved statistical significance
(P≤0.01) in PC3 (99%) compared to the VH10 cells, while in other
prostate cell lines, the gene was hypomethylated (1%).
Gene expression analysis and its
correlation with the methylation pattern
This stage was conducted to ascertain whether
methylation of the promoter CpG island regulates gene expression.
For this purpose, gene expression alterations were analyzed using
the qPCR method. Gene expression analysis included two further cell
lines: One prostate cancer cell line PC3MPro4 (with an increased
tumorigenic potential but a low incidence of metastasis) and
wild-type fibroblasts, VH25. Gene expression in cell lines with an
increasing tumorigenic and metastatic potential was analyzed
(PC3MLN4 produced a higher incidence of distant metastases).
APC, DKK3, GPX3, GSTP1, MGMT, PTGS2,
RASSF1, TIMP2 and TNFRSF10D gene hypermethylation downregulates
gene expression
Genes whose expression was downregulated by
hypermethylation were characterized. A high methylation level was
associated with a decrease in expression in 9 out of the 20
analyzed genes (APC, DKK3, GPX3, GSTP1, MGMT, PTGS2, RASSF1,
TIMP2 and TNFRSF10D). However, hypermethylation caused a
different degree of downregulation of these genes. The expression
of all genes in prostate cancer cell lines was compared to that in
the VH10 cell line.
In the case of 5 of the previously mentioned genes
hypermethylated in prostate cancer cell lines versus the
fibroblasts, a high methylation level was associated with
downregulation of their expression (Fig. 2). The majority of gene expression
results were separately normalized in respect to PBGD and
hMRPL19. In reactions normalized in respect to PBGD,
the most significant decrease in expression in prostate cancer cell
lines was observed for the PTGS2 gene (10-fold in PC3MLN4
and PC3MPro4, and 6-fold in PC3 and PC3M cells; P≤0.05).
TIMP2 expression decreased by gradually increasing factors:
5-fold in PC3MPro4, 8-fold in PC3MLN4, 10-fold in PC3M, and 80-fold
in PC3 cells. APC was downregulated 2-fold in prostate
cancer cell lines. The most significant decrease in RASSF1
expression was noted in PC3M (9-fold), and the lowest in PC3MPro4
(~3-fold). The GPX3 expression decreased from 12-fold in
PC3MLN4 to 100-fold in PC3M when compared to the VH10 cells.
However, this observation was based on the GPX3 expression
analysis using hMRPL19 as a single reference gene. The level
of the GPX3 expression analyzed with PBGD as the
reference gene and the TaqMan probe was undetectable (data not
shown).
In contrast, DKK3, GSTP1, MGMT and
TNFRSF10D were hypermethylated in the PC3M and PC3MLN4
cells, while the methylation level of these genes in the PC3 cell
line was similar to that in VH10, close to 0%. A decrease in the
expression of these genes was observed corresponding to the
hypermethylation of their promoters (Fig. 3.)
The DKK3 gene showed 10-fold lower expression
in the PC3 cell line. Other prostate cancer cell lines showed a
much greater decrease in the expression of the DKK3 gene
(100-fold). The MGMT gene was expressed at the lowest level
in PC3MLN4 cells (8-fold lower compared to VH10), while in PC3M and
PC3MPro4, the decrease was 2- and 4-fold, respectively.
TNFRSF10D was downregulated 2-fold in prostate cancer cell
lines. Interestingly, with hMRPL19 as the reference gene, a
3-fold decrease was noted only in PC3, while in other prostate
cancer cell lines, TNFRSF10D was expressed at a 4,000-fold
lower level compared to VH10. Finally, no statistically significant
differences in the expression level of GSTP1 normalized in
respect to PBGD were found, thus GAPDH was tested
instead. The expression of GSTP1 was downregulated 300-fold
in PC3M and PC3MLN4, while in other cell lines it was comparable to
the controls. With hMRPL19 as the reference gene, a similar
decrease was noted in all analyzed prostate cancer cell lines (3-
to 4-fold). The expression of APC, DKK3, PTGS2, RASSF1,
TIMP2 after normalization in respect to hMRPL19 was also
found to be decreased, similarly as when normalized in respect to
PBGD.
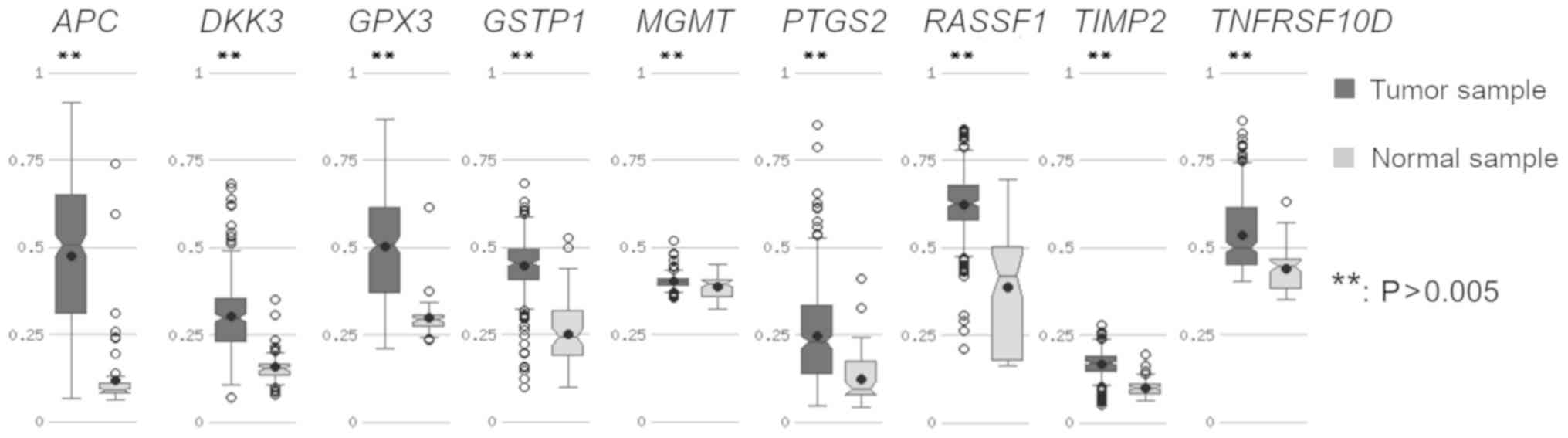
APC, DKK3, GPX3, GSTP1, MGMT, PTGS2,
RASSF1, TIMP2, TNFRSF10D methylation pattern in prostate cancer
tissue-in silico analysis
This stage aimed to establish whether the obtained
methylation results were consistent with the methylation pattern in
prostate cancer tissue. As TCGA is a project which uses
high-throughput technologies, it was decided to use integrated
human data from TCGA deposited in the MethHC web base (39,40).
The APC, DKK3, GPX3, GSTP1, MGMT, PTGS2, RASSF1,
TIMP2 and TNFRSF10D genes were hypermethylated in
prostate adenocarcinoma samples compared to normal samples (which
represented a normal tissue from the same group of prostate
adenocarcinoma patients) with statistical significance at
P≤0.005 (Fig. 4). The
greatest difference in the level of methylation between tumor and
normal sample was detected in APC, the lowest in MGMT
(Fig. 4).
PDLIM4 hypomethylation upregulates
gene expression
Genes whose hypomethylation was found to be
associated with a significantly higher expression were analyzed.
One gene classified to this group, PDLIM4, was
hypomethylated in most tested cell lines.
PDLIM4 expression was statistically
significantly higher in the PC3M, PC3MLN4 and PC3MPro4 cell lines
as well as in fibroblasts compared to that in PC3 cells.
PDLIM4 was hypermethylated in PC3. In other cell lines,
PDLIM4 was hypomethylated which was associated with a
various degree of gradual increasing expression: PC3M (0.77±0.13),
PC3MPro4 (1.56±0.08) and PC3MLN4 (3.63±0.44). In the VH10 cell
line, PDLIM4 expression was 1.0±0.14. According to the TCGA
data deposited in MethHC, the PDLIM4 gene was
hypermethylated in prostate adenocarcinoma compared to normal
samples with statistical significance at P≤0.005 (Fig. 5).
Methylation pattern of the DLC1, CAV1,
MSX1, ZNF185, CDKN2A, CDH1, RARB genes is not associated with gene
expression
Seven genes (DLC1, CAV1, MSX1, ZNF185, CDKN2A,
CDH1 and RARB) whose methylation pattern was not
associated with gene expression were distinguished (Fig. 6).
Two genes, DLC1 and CAV1, were
hypomethylated in fibroblasts as well as in prostate cancer cell
lines. Nevertheless, differences in the expression levels of these
genes were shown. In respect to PBGD as the reference gene,
DLC1 expression was at similar level in the PC3 and VH10
cell lines. In the other prostate cancer cell lines, a decrease in
expression was found: 5-fold in PC3MPro4, 7.5-fold in PC3MLN4 and
12-fold in PC3M compared to that in the VH10 cell line (1.0±0.2).
CAV1 expression was downregulated in all analyzed prostate
cancer cell lines: From 40-fold in PC3M to 6-fold in PC3MLN4.
Two other genes, ZNF185 and MSX1, were
hypermethylated in all analyzed cell lines. An interesting
observation concerned MSX1: When only prostate cancer cell
lines were considered, a gradual increase in the relative
expression level was observed for PC3 (0.34±0.03), PC3M
(0.50±0.05), PC3MPro4 (0.69±0.15), PC3MLN4 (0.99±0.15). However,
MSX1 expression in prostate cancer cell lines was lower than
in VH10. The ZNF185 gene was shown to be upregulated in
prostate cancer cell lines: 27-fold in PC3, 19-fold in PC3M,
28-fold in PC3MPro4, and 116-fold in PC3MLN4 compared to VH10
cells. Interestingly, the values of relative expression of
ZNF185 obtained in PC3, PC3M and PC3MPro4 were comparable
with the expression level in the second WT control, VH25.
Therefore, they can be considered as falling within the normal
range. The ZNF185 expression results were presented with
hMRPL19 as the reference gene. In respect to PBGD as
the reference, the gene expression pattern was consistent; however,
the difference in the expression level between VH10 and PC3MLN4 was
much higher (data not shown).
In the case of the CDKN2A gene, its
methylation pattern as well as expression level differed between
prostate cell lines and fibroblasts. Although the CDKN2A
gene was hypermethylated in prostate cancer cell lines, it was
upregulated in these cells compared to VH10 and VH25. The increase
in expression was gradual: From 7-fold in PC3, 19-fold in PC3M,
21-fold in PC3MPro4, to 31-fold in the PC3MLN4 cell line.
Expression of DLC1, CAV1, MSX1 and
CDKN2A after normalization in respect to hMRPL19
indicated a pattern similar to that obtained with PBGD as
the reference gene. Despite the varied methylation pattern of the
CDH1 and RARB genes in prostate cancer cell lines and
fibroblasts, no statistically significant differences in expression
level were noted (data not shown).
Discussion
Abnormal expression of genes in cancer cells can
arise from epigenetic changes, but also changes in the number of
copies and/or the presence of sequence mutations. Changes in the
epigenome are often crucial for the functioning of cells and thus
are often associated with carcinogenesis, metastasis and response
to chemotherapy. Therefore, unique expression and methylation
patterns have been introduced in diagnostics as prognostic and
predictive biomarkers. Since the best-known expression profiling
performed 15 years ago in breast cancer (41), determination of the unique signature
of gene expression (6,42) or promoter methylation is used more
frequently, for example MGMT methylation in tumor tissue as
a biomarker in glioma (43) or
SEPT9 methylation in plasma as a biomarker in colorectal
adenocarcinoma (44).
Over the last few years, a broad spectrum of
different technologies has been introduced for the quantitative and
qualitative measurement of DNA methylation status. The most
commonly used method is sodium bisulfite conversion of genomic DNA
to differentiate and detect unmethylated and methylated cytosines
using methylation-specific PCR, MassARRAY EpiTYPER,
hybridization-based promoter and CpG island microarrays. In this
study, a less-known methodology based on input DNA treatment with a
methylation-sensitive and methylation-dependent restriction enzyme
followed by qPCR was used. Commercial EpiTect Methyl II PCR Array,
Signature Panel: EAHS-051Z (Qiagen) kit was selected for
methylation analysis in prostate cancer to examine the methylation
profile of CpG islands in 22 cancer-related genes in prostate
cancer cell lines with increasing tumorigenic and metastatic
potential (30). According to the
EpiTect Methyl II PCR Array System manufacturer, every target
region is selected within one CpG island or CpG-dense area
predicted from both the UCSC database and published data with
functional annotation (45).
Further analyses included determination of gene expression and
comparison of data from prostate cancer cell lines with the
methylation status of CpG islands in the promoter regions of human
prostate cancer from the MethHC database (39).
The expression and methylation status of GSTP1,
APC, RASSF1A, MGMT and PTGS2 have previously been well
characterized in prostate cancer (46), and gene expression has frequently
been evaluated using microarrays. High-throughput methods, such as
microarrays, are an excellent screening tool, although the results
require validation using quantitative real-time RT-PCR (qPCR)
assays. Likewise, the results of methylation profiling have often
been obtained using less advanced technologies, such as
methylation-specific PCR (MSP) (46). Our study for the first time analyzed
these genes in PC3-derived cell lines (PC3M, PC3MLN4 and PC3MPro4)
in reference to fibroblasts (VH10 and VH25). qPCR used in this
study allowed a precise determination of relative gene expression,
normalized in respect to the expression of different reference
genes.
According to the results obtained earlier in
prostate cancer, it was shown that 12 genes out of 20 analyzed were
hypermethylated: 8 genes were hypermethylated in prostate cancer
cell lines compared to VH10 (APC, CDKN2A, EDNRB, GPX3, PTGS2,
SLC5A8, TIMP2, RASSF1), and 5 genes were hypermethylated in
PC3M and PC3MLN4 compared to PC3 and VH10 cell lines (DKK3,
MGMT, TNFRSF10D, RARB and GSTP1). Furthermore, 3 genes
(CAV1, DLC1, PDLIM4) were found to be hypomethylated in
prostate cancer cell lines. Interestingly, methylation was found to
regulate the expression of half of the analyzed genes (APC,
DKK3, GPX3, GSTP1, MGMT, PDLIM4, PTGS2, RASSF1, TIMP2 and
TNFRSF10D) in PC3-derived cell lines and fibroblasts VH10
and VH25.
APC hypermethylation leads to the
stabilization of β-catenin in the cytoplasm due to deregulation of
β-catenin degradation (10).
Previous studies have also demonstrated that APC
hypermethylation is a common occurrence in the PC3 cell line
(34), prostate cancer and its
progression (47), as well as in
other cancers, such as colon and gastric cancer (48).
DKK3 hypermethylation has been observed in
pancreatic cancer lines and in breast cancer tissue (49,50).
It has been found that this gene is also associated with β-catenin
expression. DKK3 overexpression in transfected cells
resulted in a decrease in β-catenin expression (49). Inactivation of the DKK3 gene
is also common in prostate cancer, in which the level of the Dkk3
protein is inversely correlated with the Gleason degree, and the
lowest level was noted in tumors that are probably metastatic
(51,52).
A wide-spectrum analysis of androgen-dependent
(LNCaP, and Du145) and androgen-independent (PC3) prostate cancer
cell lines allowed characterization of GPX3 as a novel
tumor-suppressor gene, as the level of GPX3 expression was
associated with prostate tumor stage (35). Our findings in cell lines and the
MethHC database are consistent with the widespread hypermethylation
of GPX3 in prostate cancer (53–55).
The methylation status of GSTP1, as for
APC, is common in prostate cancer (56,57).
However, evidence of a relationship between the level of
GSTP1 methylation and disease progression is contradictory.
Some studies have shown that the hypermethylation of this gene is
associated with prostate cancer progression (58–60).
On the other hand, the prognostic value of GSTP1 methylation
has not been demonstrated (47). In
our study, methylation as well as the expression level provided
prognostic information, but the methylation level increased only to
15% in PC3MLN4 cells.
MGMT is an important glioblastoma prognostic
and predictive biomarker in clinical use (43). Reports regarding prostate cancer are
inconclusive (46,56,61).
The TCGA results showed that the increase in methylation between
tumor and normal tissue was small; however, the difference was
statistically significant. On the other hand, MGMT
hypermethylation in prostate cancer has been previously reported
(46,61). Our results revealed no significant
difference in the MGMT methylation level between the PC3 and
VH10 cell lines. The MGMT gene was hypermethylated only in
the PC3M and PC3MLN4 lines, which was associated with a decrease in
expression in these cell lines.
PTGS2 was also hypermethylated in all
prostate cancer cell lines compared to fibroblasts. However, the
methylation status of PTGS2 varied between the prostate
cancer cell lines and it seems that methylation is the main
mechanism of PTGS2 regulation in prostate cancer cell lines,
as well as in prostate cancer patients-TCGA data (39). Recent studies employing qPCR for
methylation analysis have shown that PTGS2 hypermethylation
is a potential sensitive and specific prostate cancer biomarker in
ctDNA isolated from the blood of PCa patients (62), as well as in prostate cancer tissue
(63). Although PTGS2
hypermethylation has been observed in 68% cases of PCa versus 15%
of BPH tissues (62), it seems that
the methylation status of PTGS2 alone is not sufficient.
However, it definitely should be included in molecular profiling to
improve efficiency.
A previous study demonstrated that PDLIM4
can function as a tumor suppressor in prostate cancer cells
(21). PDLIM4 mRNA
expression was found to be reduced in PC3 prostate cancer cells
(21), which is consistent with our
results, but notably, in other prostate cancer cell lines derived
from PC3, PDLIM4 expression increased gradually with the
increase in PNC. Our results may suggest a novel oncogenic function
of PDLIM4 in prostate cancer cell lines derived from PC3.
Putatively, methylation is the main mechanism of PDLIM4
regulation in prostate cancer cell lines, as in renal cancer and
acute myelogenous leukemia (64,65).
RASSF1 hypermethylation, which has been
reported in many types of cancers, including prostate cancer, can
lead to disorders in the DNA repair pathway and cell cycle control
(66). The relationship between the
RASSF1 methylation level and prostate cancer aggressiveness
has been noted (66,67). Downregulation of TIMP2 has
also been correlated with cancer progression and metastasis
(68). The results obtained in
prostate cancer indicate an antitumor effect of the Timp2 protein
(69). However, reports of this
phenomenon are contradictory (70).
Our results showed hypermethylation of the RASSF1 and
TIMP2 genes in prostate cancer cell lines compared to
fibroblasts.
Hypermethylation of TNFRSF10D has been
noted; for example, in melanoma and prostate cancer (71,72).
In prostate cancer cell lines in our study, the TNFRSF10D
gene was hypermethylated only in the PC3M and PC3MLN4 cells.
Nevertheless, a decrease in expression was observed in all tested
prostate cancer cell lines. The TNFRSF10D expression level
was similar in all prostate cancer cell lines in respect to PBGD as
the reference gene, while with hMRPL19, greater downregulation in
PC3M, PC3MLN4 and PC3MPro4 was observed. This observation confirms
how important it is to use at least two reference genes for the
analysis of relative gene mRNA expression.
It should be noted that the results of CpG island
methylation analysis in the promoter regions obtained in the
prostate cancer cell lines for those 10 genes were consistent with
clinical data obtained from 336 prostate cancer patients in the
TCGA project. Moreover, the methylation signature panel used in
this study included genes methylated in prostate cancer cell lines,
but no changes in their expression (EDNRB and SLC5A8)
were shown. This also included genes with an altered expression
level between prostate cancer cell lines and fibroblasts, but was
not consistent with the methylation pattern. Those genes are
probably regulated by other mechanisms, such as small RNA
molecules, e.g., miRNAs (73),
changes in chromatin conformation (74) or histone modifications (75).
Acknowledgements
Not applicable.
Funding
This research was funded by the Foundation for
Polish Science (grant no. HOMING PLUS/2010-2/7) and The Ludwik
Rydygier Collegium Medicum, Nicolaus Copernicus University (grant
no. MN-SDL-5/WL/2017).
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
Conceptualization of the study design was achieved
by MAL. The research methodology was conceived by KK, MAL, AB and
SH. Software analysis of data was performed by KK and MAL;
validation of the data was accomplished by KK; formal analysis was
conducted by KK and MAL. Investigation was carried out by KK;
resources were the responsibility of JK and MAL. Data curation was
performed by MAL. Writing; original draft preparation was carried
out by KK and MAL; Writing; review and editing was accomplished by
AB, JK and SH; visualization was conducted by KK; supervision was
conducted by SH and MAL. Software analysis of data was performed by
KK; validation of the data was accomplished by KK; formal analysis
was conducted by KK and MAL. Investigation was carried out by KK;
resources were the responsibility of JK and MAL; project
administration was conducted by JK and MAL; and funding acquisition
by MAL. All authors read and approved the manuscript and agree to
be accountable for all aspects of the research in ensuring that the
accuracy or integrity of any part of the work are appropriately
investigated and resolved.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare no conflict of interest. The
funders had no role in the design of the study; in the collection,
analyses, or interpretation of data; in the writing of the
manuscript, or in the decision to publish the results.
References
|
1
|
The Cancer Genome Atlas-National Cancer
Institute. https://cancergenome.nih.gov/2018 10 222011
|
|
2
|
Baylin SB and Herman JG: DNA
hypermethylation in tumorigenesis: Epigenetics joins genetics.
Trends Genet. 16:168–174. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yang M and Park JY: DNA methylation in
promoter region as biomarkers in prostate cancer. Methods Mol Biol.
863:67–109. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Golub TR, Slonim DK, Tamayo P, Huard C,
Gaasenbeek M, Mesirov JP, Coller H, Loh ML, Downing JR, Caligiuri
MA, et al: Molecular classification of cancer: Class discovery and
class prediction by gene expression monitoring. Science.
286:531–537. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
van't Veer LJ, Dai H, van de Vijver MJ, He
YD, Hart AA, Mao M, Peterse HL, van der Kooy K, Marton MJ,
Witteveen AT, et al: Gene expression profiling predicts clinical
outcome of breast cancer. Nature. 415:530–536. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Salazar R, Roepman P, Capella G, Moreno V,
Simon I, Dreezen C, Lopez-Doriga A, Santos C, Marijnen C, Westerga
J, et al: Gene expression signature to improve prognosis prediction
of stage II and III colorectal cancer. J Clin Oncol. 29:17–24.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Guo L, Ma Y, Ward R, Castranova V, Shi X
and Qian Y: Constructing molecular classifiers for the accurate
prognosis of lung adenocarcinoma. Clin Cancer Res. 12:3344–3354.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Toyota M, Ahuja N, Ohe-Toyota M, Herman
JG, Baylin SB and Issa JP: CpG island methylator phenotype in
colorectal cancer. Proc Natl Acad Sci USA. 96:8681–8686. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Shi H, Chen J, Li Y, Li G, Zhong R, Du D,
Meng R, Kong W and Lu M: Identification of a six microRNA signature
as a novel potential prognostic biomarker in patients with head and
neck squamous cell carcinoma. Oncotarget. 7:21579–21590.
2016.PubMed/NCBI
|
|
10
|
Voronkov A and Krauss S: Wnt/beta-catenin
signaling and small molecule inhibitors. Curr Pharm Des.
19:634–664. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Liu P, Rudick M and Anderson RG: Multiple
functions of caveolin-1. J Biol Chem. 277:41295–41298. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Rayess H, Wang MB and Srivatsan ES:
Cellular senescence and tumor suppressor gene p16. Int J Cancer.
130:1715–1725. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Angst BD, Marcozzi C and Magee AI: The
cadherin superfamily: Diversity in form and function. J Cell Sci.
114:629–641. 2001.PubMed/NCBI
|
|
14
|
Kim TY, Vigil D, Der CJ and Juliano RL:
Role of DLC-1, a tumor suppressor protein with RhoGAP activity, in
regulation of the cytoskeleton and cell motility. Cancer Metastasis
Rev. 28:77–83. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Niehrs C: Function and biological roles of
the Dickkopf family of Wnt modulators. Oncogene. 25:7469–7481.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Arai H, Nakao K, Takaya K, Hosoda K, Ogawa
Y, Nakanishi S and Imura H: The human endothelin-B receptor gene.
Structural organization and chromosomal assignment. J Biol Chem.
268:3463–3470. 1993.PubMed/NCBI
|
|
17
|
Chen B, Rao X, House MG, Nephew KP, Cullen
KJ and Guo Z: GPx3 promoter hypermethylation is a frequent event in
human cancer and is associated with tumorigenesis and chemotherapy
response. Cancer Lett. 309:37–45. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hayes JD and Pulford DJ: The glutathione
S-transferase supergene family: Regulation of GST and the
contribution of the isoenzymes to cancer chemoprotection and drug
resistance. Crit Rev Biochem Mol Biol. 30:445–600. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kaina B, Christmann M, Naumann S and Roos
WP: MGMT: Key node in the battle against genotoxicity,
carcinogenicity and apoptosis induced by alkylating agents. DNA
Repair. 6:1079–1099. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Bonito NA, Borley J, Wilhelm-Benartzi CS,
Ghaem-Maghami S and Brown R: Epigenetic regulation of the homeobox
gene MSX1 associates with platinum-resistant disease in high-grade
serous epithelial ovarian cancer. Clin Cancer Res. 22:3097–3104.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Vanaja DK, Grossmann ME, Cheville JC, Gazi
MH, Gong A, Zhang JS, Ajtai K, Burghardt TP and Young CY: PDLIM4,
an actin binding protein, suppresses prostate cancer cell growth.
Cancer Invest. 27:264–272. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chandrasekharan NV and Simmons DL: The
cyclooxygenases. Genome Biol. 5:2412004. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Tang D, Kryvenko ON, Mitrache N, Do KC,
Jankowski M, Chitale DA, Trudeau S, Rundle A, Belinsky SA and
Rybicki BA: Methylation of the RARB gene increases prostate cancer
risk in black Americans. J Urol. 190:317–324. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Volodko N, Gordon M, Salla M, Ghazaleh HA
and Baksh S: RASSF tumor suppressor gene family: Biological
functions and regulation. FEBS Lett. 588:2671–2684. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Mo S, Su Z, Heng B, Chen W, Shi L, Du X
and Lai C: SFRP1 promoter methylation and renal carcinoma risk: A
systematic review and meta-analysis. J Nippon Med Sch. 85:78–86.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ganapathy V, Thangaraju M, Gopal E, Martin
PM, Itagaki S, Miyauchi S and Prasad PD: Sodium-coupled
monocarboxylate transporters in normal tissues and in cancer. AAPS
J. 10:193–199. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Brew K and Nagase H: The tissue inhibitors
of metalloproteinases (TIMPs): An ancient family with structural
and functional diversity. Biochim Biophys Acta. 1803:55–71. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Pan G, Ni J, Wei YF, Yu G, Gentz R and
Dixit VM: An antagonist decoy receptor and a death
domain-containing receptor for TRAIL. Science. 277:815–818. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhang JS, Gong A and Young CY: ZNF185, an
actin-cytoskeleton-associated growth inhibitory LIM protein in
prostate cancer. Oncogene. 26:111–122. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Pettaway CA, Pathak S, Greene G, Ramirez
E, Wilson MR, Killion JJ and Fidler IJ: Selection of highly
metastatic variants of different human prostatic carcinomas using
orthotopic implantation in nude mice. Clin Cancer Res. 2:1627–1636.
1996.PubMed/NCBI
|
|
31
|
Wang C, Norton JT, Ghosh S, Kim J, Fushimi
K, Wu JY, Stack MS and Huang S: Polypyrimidine tract-binding
protein (PTB) differentially affects malignancy in a cell
line-dependent manner. J Biol Chem. 283:20277–20287. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Godthelp BC, van Buul PP, Jaspers NG,
Elghalbzouri-Maghrani E, van Duijn-Goedhart A, Arwert F, Joenje H
and Zdzienicka MZ: Cellular characterization of cells from the
Fanconi anemia complementation group, FA-D1/BRCA2. Mutat Res.
601:191–201. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wiegant WW, Meyers M, Verkaik NS, van der
Burg M, Darroudi F, Romeijn R, Bernatowska E, Wolska-Kusnierz B,
Mikoluc B, Jaspers NG, et al: A novel radiosensitive SCID patient
with a pronounced G2/M sensitivity. DNA Repair.
9:365–373. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Bastian PJ, Ellinger J, Wellmann A,
Wernert N, Heukamp LC, Müller SC and von Ruecker A: Diagnostic and
prognostic information in prostate cancer with the help of a small
set of hypermethylated gene loci. Clin Cancer Res. 11:4097–4106.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yu YP, Yu G, Tseng G, Cieply K, Nelson J,
Defrances M, Zarnegar R, Michalopoulos G and Luo JH: Glutathione
peroxidase 3, deleted or methylated in prostate cancer, suppresses
prostate cancer growth and metastasis. Cancer Res. 67:8043–8050.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Norton JT, Pollock CB, Wang C, Schink JC,
Kim JJ and Huang S: Perinucleolar Compartment prevalence is a
phenotypic pancancer marker of malignancy. Cancer. 113:861–869.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Frycz B, Pinczewska A and Jagodziński PP:
Maślan sodu obniża ekspresję dehydrogenazy 17β-hydroksysteroidowej
typu 1-szego W linii komórkowej raka gruczołu krokowego LNCaP.
Nowiny Lekarskie. 80:283–287. 2011.
|
|
38
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2−ΔΔCT method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Huang WY, Hsu SD, Huang HY, Sun YM, Chou
CH, Weng SL and Huang HD: MethHC: A database of DNA methylation and
gene expression in human cancer. Nucleic Acids Res. 43:D856–D861.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
MethHC, . A database of DNA methylation
and gene expression in human cancers. http://methhc.mbc.nctu.edu.tw/php/index.phpOctober
23–2018
|
|
41
|
van de Vijver MJ, He YD, van't Veer LJ,
Dai H, Hart AA, Voskuil DW, Schreiber GJ, Peterse JL, Roberts C,
Marton MJ, et al: A gene-expression signature as a predictor of
survival in breast cancer. N Engl J Med. 347:1999–2009. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
O'Connell MJ, Lavery I, Yothers G, Paik S,
Clark-Langone KM, Lopatin M, Watson D, Baehner FL, Shak S, Baker J,
et al: Relationship between tumor gene expression and recurrence in
four independent studies of patients with stage II/III colon cancer
treated with surgery alone or surgery plus adjuvant fluorouracil
plus leucovorin. J Clin Oncol. 28:3937–3944. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Roszkowski K, Furtak J, Zurawski B,
Szylberg T and Lewandowska MA: Potential role of methylation marker
in glioma supporting clinical decisions. Int J Mol Sci. 17(pii):
E18762016. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Song L, Peng X, Li Y, Xiao W, Jia J, Dong
C, Gong Y, Zhou G and Han X: The SEPT9 gene methylation assay is
capable of detecting colorectal adenoma in opportunistic screening.
Epigenomics. 9:599–610. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Jiang Q, Liu CX, Gu X, Wilt G, Shaffer J,
Zhang Y and Devgan V: EpiTect Methyl II PCR Array System: A simple
tool for screening regional DNA methylation of a large number of
genes or samples without bisulfite conversion. Qiagen. https://www.qiagen.com/ch/resources/resourcedetail?id=39ec06aa-ec53-4acd-aa15-67b5882efbb6&lang=en(cited
2018-11-09).
|
|
46
|
Kang GH, Lee S, Lee HJ and Hwang KS:
Aberrant CpG island hypermethylation of multiple genes in prostate
cancer and prostatic intraepithelial neoplasia. J Pathol.
202:233–240. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Richiardi L, Fiano V, Vizzini L, De Marco
L, Delsedime L, Akre O, Tos AG and Merletti F: Promoter methylation
in APC, RUNX3, and GSTP1 and mortality in prostate cancer patients.
J Clin Oncol. 27:3161–3168. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Llorca-Cardeñosa MJ, Fleitas T,
Ibarrola-Villava M, Peña-Chilet M, Mongort C, Martinez-Ciarpaglini
C, Navarro L, Gambardella V, Castillo J, Roselló S, et al:
Epigenetic changes in localized gastric cancer: The role of RUNX3
in tumor progression and the immune microenvironment. Oncotarget.
7:63424–63436. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Gu YM, Ma YH, Zhao WG and Chen J:
Dickkopf3 overexpression inhibits pancreatic cancer cell growth in
vitro. World J Gastroenterol. 17:3810–3817. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Veeck J, Wild PJ, Fuchs T, Schüffler PJ,
Hartmann A, Knüchel R and Dahl E: Prognostic relevance of
Wnt-inhibitory factor-1 (WIF1) and Dickkopf-3 (DKK3) promoter
methylation in human breast cancer. BMC Cancer. 9:2172009.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Romero D and Kypta R: Dickkopf-3 function
in the prostate: Implications for epithelial homeostasis and tumor
progression. Bioarchitecture. 3:42–44. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Romero D, Kawano Y, Bengoa N, Walker MM,
Maltry N, Niehrs C, Waxman J and Kypta R: Downregulation of
Dickkopf-3 disrupts prostate acinar morphogenesis through
TGF-β/Smad signalling. J Cell Sci. 126:1858–1867. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Lodygin D, Epanchintsev A, Menssen A,
Diebold J and Hermeking H: Functional epigenomics identifies genes
frequently silenced in prostate cancer. Cancer Res. 65:4218–4227.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Yu YP, Paranjpe S, Nelson J, Finkelstein
S, Ren B, Kokkinakis D, Michalopoulos G and Luo JH: High throughput
screening of methylation status of genes in prostate cancer using
an oligonucleotide methylation array. Carcinogenesis. 26:471–479.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Falck E, Karlsson S, Carlsson J, Helenius
G, Karlsson M and Klinga-Levan K: Loss of glutathione peroxidase 3
expression is correlated with epigenetic mechanisms in endometrial
adenocarcinoma. Cancer Cell Int. 10:462010. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Yegnasubramanian S, Kowalski J, Gonzalgo
ML, Zahurak M, Piantadosi S, Walsh PC, Bova GS, De Marzo AM, Isaacs
WB and Nelson WG: Hypermethylation of CpG islands in primary and
metastatic human prostate cancer. Cancer Res. 64:1975–1986. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Strand SH, Orntoft TF and Sorensen KD:
Prognostic DNA methylation markers for prostate cancer. Int J Mol
Sci. 15:16544–16576. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Rosenbaum E, Hoque MO, Cohen Y, Zahurak M,
Eisenberger MA, Epstein JI, Partin AW and Sidransky D: Promoter
hypermethylation as an independent prognostic factor for relapse in
patients with prostate cancer following radical prostatectomy. Clin
Cancer Res. 11:8321–8325. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Richiardi L, Fiano V, Grasso C, Zugna D,
Delsedime L, Gillio-Tos A and Merletti F: Methylation of APC and
GSTP1 in non-neoplastic tissue adjacent to prostate tumour and
mortality from prostate cancer. PLoS One. 8:e681622013. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Bastian PJ, Palapattu GS, Lin X,
Yegnasubramanian S, Mangold LA, Trock B, Eisenberger MA, Partin AW
and Nelson WG: Preoperative serum DNA GSTP1 CpG island
hypermethylation and the risk of early prostate-specific antigen
recurrence following radical prostatectomy. Clin Cancer Res.
11:4037–4043. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Konishi N, Nakamura M, Kishi M, Nishimine
M, Ishida E and Shimada K: DNA hypermethylation status of multiple
genes in prostate adenocarcinomas. Jpn J Cancer Res. 93:767–773.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Ellinger J, Bastian PJ, Haan KI, Heukamp
LC, Buettner R, Fimmers R, Mueller SC and von Ruecker A:
Noncancerous PTGS2 DNA fragments of apoptotic origin in sera of
prostate cancer patients qualify as diagnostic and prognostic
indicators. Int J Cancer. 122:138–143. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Vasiljević N, Wu K, Brentnall AR, Kim DC,
Thorat MA, Kudahetti SC, Mao X, Xue L, Yu Y, Shaw GL, et al:
Absolute quantitation of DNA methylation of 28 candidate genes in
prostate cancer using pyrosequencing. Dis Markers. 30:151–161.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Morris MR, Ricketts C, Gentle D,
Abdulrahman M, Clarke N, Brown M, Kishida T, Yao M, Latif F and
Maher ER: Identification of candidate tumour suppressor genes
frequently methylated in renal cell carcinoma. Oncogene.
29:2104–2117. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Boumber YA, Kondo Y, Chen X, Shen L,
Gharibyan V, Konishi K, Estey E, Kantarjian H, Garcia-Manero G and
Issa JP: RIL, a LIM gene on 5q31, is silenced by methylation in
cancer and sensitizes cancer cells to apoptosis. Cancer Res.
67:1997–2005. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Liu L, Yoon JH, Dammann R and Pfeifer GP:
Frequent hypermethylation of the RASSF1A gene in prostate cancer.
Oncogene. 21:6835–6840. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Maruyama R, Toyooka S, Toyooka KO, Virmani
AK, Zöchbauer- Müller S, Farinas AJ, Minna JD, McConnell J, Frenkel
EP and Gazdar AF: Aberrant promoter methylation profile of prostate
cancers and its relationship to clinicopathological features. Clin
Cancer Res. 8:514–519. 2002.PubMed/NCBI
|
|
68
|
Imren S, Kohn DB, Shimada H, Blavier L and
DeClerck YA: Overexpression of tissue inhibitor of
metalloproteinases-2 retroviral-mediated gene transfer in vivo
inhibits tumor growth and invasion. Cancer Res. 56:2891–2895.
1996.PubMed/NCBI
|
|
69
|
Pulukuri SM, Patibandla S, Patel J, Estes
N and Rao JS: Epigenetic inactivation of the tissue inhibitor of
metalloproteinase-2 (TIMP-2) gene in human prostate tumors.
Oncogene. 26:5229–5237. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Ross JS, Kaur P, Sheehan CE, Fisher HA,
Kaufman RA Jr and Kallakury BV: Prognostic significance of matrix
metalloproteinase 2 and tissue inhibitor of metalloproteinase 2
expression in prostate cancer. Mod Pathol. 16:198–205. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Ratzinger G, Mitteregger S, Wolf B, Berger
R, Zelger B, Weinlich G, Fritsch P, Goebel G and Fiegl H:
Association of TNFRSF10D DNA-methylation with the survival of
melanoma patients. Int J Mol Sci. 15:11984–11995. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Hornstein M, Hoffmann MJ, Alexa A,
Yamanaka M, Müller M, Jung V, Rahnenführer J and Schulz WA: Protein
phosphatase and TRAIL receptor genes as new candidate tumor genes
on chromosome 8p in prostate cancer. Cancer Genomics Proteomics.
5:123–136. 2008.PubMed/NCBI
|
|
73
|
Catalanotto C, Cogoni C and Zardo G:
MicroRNA in control of gene expression: An overview of nuclear
functions. Int J Mol Sci. 17(pii): E17122016. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Kubiak M and Lewandowska MA: Can chromatin
conformation technologies bring light into human molecular
pathology? Acta Biochim Pol. 62:483–489. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Audia JE and Campbell RM: Histone
modifications and cancer. Cold Spring Harb Perspect Biol.
8:a0195212016. View Article : Google Scholar : PubMed/NCBI
|