Introduction
Lung cancer is a main cause of cancer-associated
mortality worldwide. Non-small cell lung cancer (NSCLC) accounts
for ~80% of all lung cancer cases (1–4). Despite
the rapid development in medical science in recent years, the
prognosis of patients with NSCLC remains unsatisfactory, with an
overall 5-year relative survival rate of only 18%, which is reduced
to 4% for metastatic cancer (4).
Therefore, it is essential to seek a novel therapy, and to identify
an exact signaling pathway or molecular target protein responsible
for NSCLC proliferation and metastasis.
The principal constituents of turmeric extracts
include curcumin, demethoxycurcumin, bisdemethoxycurcumin and
cyclocurcumin (5). Among these
compounds, curcumin has been the most extensively studied natural
compound in the last few decades due to its pleiotropic effects on
numerous molecular targets in different diseases (6–9). Previous
studies regarding curcumin have mainly focused on its
anti-inflammatory abilities, whereas its effective anticancer
activity was not recognized until the early 1990s (10,11).
The Toll-like receptor 4 (TLR4)/MyD88 pathway is a
classic signaling pathway associated with inflammation. Cancer is
closely related to inflammation, particularly chronic inflammation
(12–15). Previous studies have reported that
aberrant expression of TLR4 is involved in several types of human
cancer, including gastric cancer (16), lung cancer (17), prostate cancer (18), glioblastoma (19) and colorectal cancer (20). However, the specific role of TLR4 in
NSCLC cell proliferation and metastasis is currently unknown.
Epidermal growth factor receptor (EGFR) is a
receptor tyrosine kinase, which is overexpressed or aberrantly
activated in various cancer cells and regulates numerous complex
pathological processes, including cell proliferation,
differentiation, invasion and migration (21). A previous study reported that
downregulation of EGFR by curcumin inhibits cell growth and induces
apoptosis in NSCLC cells (22).
Furthermore, several studies have detected intracellular cross talk
between EGFR signaling and the TLR4 pathway (23–25).
Research in gastric epithelial cells has demonstrated that TLR4 is
able to transactivate the EGFR pathway through Helicobacter
pylori protein HP0175, resulting in the pathophysiology of
ulcerogenesis and/or carcinogenesis (24). Therefore, it was hypothesized that
curcumin may suppress the proliferation and metastasis of NSCLC
through the TLR4/MyD88 and EGFR pathways.
Materials and methods
Clinical samples
Tissue specimens were obtained from the Department
of Pathology, The Third Affiliated Hospital of Kunming Medical
University (Tumor Hospital of Yunnan Province) between May 2003 and
July 2010. Specimens were fixed with 10% neutral formalin for 72 h
at room temperature and were then embedded in paraffin. The
specimens consisted of 52 primary NSCLC tumor tissues and 49 benign
lung tissues. The NSCLC specimens were obtained from 52 patients:
40 with adenocarcinoma and 12 with squamous cell carcinoma,
including 30 men and 22 women, with ages ranging between 34 and 70
years (mean age, 59 years). The benign lung tissues were obtained
from 49 patients with benign pulmonary diseases: 29 men and 20
women, with ages ranging between 32 and 70 years (mean age, 57
years). All patients underwent primary tumor resection, and the
majority also received lymph node dissection. Patients with a
diagnosis of relapse who had received preoperative radiation,
chemotherapy or biotherapy were excluded to avoid any alterations
in tumor marker determination resulting from treatment. Patients
diagnosed with multiple primary cancers in other organs or tissues
were also excluded. The study was approved by the ethics committee
of The Third Affiliated Hospital of Kunming Medical University, and
all patients provided written informed consent and authorization
for use of biological specimens. Demographic and clinical data were
obtained from the patients' medical records.
Pathology
A routine histological examination was performed
with hematoxylin-eosin staining at room temperature; the stained
slices were reviewed independently by three pathologists under a
light microscope. Benign lung tissues were collected from a normal
section of the lung in patients with a benign pulmonary disease
identified by pathologists. All carcinomas were classified in
accordance with the 7th edition of the American Joint Committee on
Cancer staging system (26).
Immunohistochemistry (IHC)
Samples were processed for immunohistochemical
analysis, in order to detect TLR4 and MyD88 expression levels and
distribution patterns. Briefly, 4-µm sections of paraffin-embedded
tissues were mounted on charged glass slides and baked at 70°C for
1 h. The slides were allowed to cool to room temperature,
deparaffinized in xylene and rehydrated in a graded alcohol series.
Sections were then microwave-treated for 5 min in citrate buffer
(pH 6.0) for antigen retrieval, and endogenous peroxidase activity
was blocked by incubation in 0.3% hydrogen peroxide for 20 min at
room temperature. Mouse monoclonal TLR4 (cat. no. SAB1404475;
Sigma-Aldrich; Merck KGaA) and rabbit monoclonal Myd88 (cat. no.
ab133739; Abcam) antibodies were used to detect TLR4 and Myd88
protein expression, respectively, at 1:600 and 1:250 dilutions in
PBS; sections were incubated with these antibodies at 4°C
overnight. After two washes in PBS, slides were incubated with
undiluted rabbit secondary antibodies from a Dako REAL EnVision
detection system/Horseradish Peroxidase for rabbit/mouse secondary
antibodies kit (cat. no. K5007; Agilent Technologies, Inc.) for 30
min at room temperature, The peroxidase reaction was developed
using 3,3′-diaminobenzidine (DAB) chromogen solution (Dako; Agilent
Technologies, Inc.). Sections were visualized with DAB and
counterstained with hematoxylin for 2 min at room temperature.
Sections were scored semi-quantitatively under a light microscope
for the extent of the immunoreaction as follows: 0, 0%
immunoreactive cells; 1, <20% immunoreactive cells; 2, 20–50%
immunoreactive cells; and 3, >50% immunoreactive cells. In
addition, the intensity of staining was scored semi-quantitatively
as 0, negative; 1, weak; 2, intermediate; and 3, strong. The final
immunoreaction score was defined as the sum of both parameters
(extent and intensity), and samples were grouped according to the
summed score as negative (0), weak staining (1–3), moderate
staining (4–6) and strong staining (7–9). Final
immunoreaction scores >0 were defined as positive. All slides
were evaluated independently for protein expression by three
separate observers; slides with an incongruent grading were
scrutinized a second time, and a consensus was reached.
Cell lines
The NSCLC cell lines NCI-A549 (human lung
adenocarcinoma) and NCI-H226 (human lung squamous cancer) were used
in the present study. The H226 cell line was purchased from the
Cell Bank of Chinese Academy of Sciences, and the A549 cell line
was obtained from the American Type Culture Collection. A549 and
H226 cells were respectively grown in DMEM F12 (Gibco; Thermo
Fisher Scientific, Inc.) and RPMI-1640 (Gibco; Thermo Fisher
Scientific, Inc.) supplemented with 10% fetal bovine serum (FBS;
Gibco; Thermo Fisher Scientific, Inc.) and penicillin (1,000
U/ml)/streptomycin (10 mg/ml) (cat. no. SV30010; Hyclone; GE
Healthcare) at 37°C in an incubator containing 5%
CO2.
Cell treatment
For the dose response analysis, A549 and H226 cells
were treated with different concentrations (A549, 5, 10, 15 µM;
H226, 5, 10, 20 µM) of curcumin for 72 h in a 37°C incubator
containing 5% CO2. For the analysis of time-dependent
response, A549 and H226 cells were treated with curcumin (10 µM)
for various durations (0, 24, 48 and 72 h). To investigate the
signaling pathway, A549 and H226 cells were pretreated with either
DMSO or curcumin (10 µM) for 72 h, followed by a 12-h
serum-starvation period. After the aforementioned treatments, cells
were sequentially grown in an incubator with 5% CO2 at
37°C. EGF (100 ng/ml) was added to the cell culture dish at
different time points (1.5, 3 and 6 h) after serum starvation.
Following the application of the aforementioned treatments, cells
were placed in an incubator with 5% CO2 at 37°C. Then,
cells in the logarithmic growth phase were collected to perform
western blot analysis.
Western blot analysis
Cells (A549 and H226) were harvested after drug
treatment and lysed in RIPA buffer (cat. no. 89900; Pierce; Thermo
Fisher Scientific, Inc.). The lysates were centrifuged at 12,000 ×
g for 20 min at 4°C, and the supernatants were collected. Protein
concentration was measured using the bicinchoninic acid assay (cat.
no. 23227; Pierce; Thermo Fisher Scientific, Inc.). For each
sample, 50 µg protein lysate was loaded per well, and protein
samples were separated by 10% SDS-PAGE and transferred onto
polyvinylidene fluoride membranes (cat. no. ISEQ00010; EMD
Millipore) by electroblotting. Membranes were pretreated with 8%
non-fat dry milk in TBS-Tween-20 (0.05–0.1%) (TBS-T) for 2 h at
room temperature, followed by incubation with primary antibodies
for 16 h at 4°C. The primary antibodies used targeting the
following proteins were diluted at 1:1,000 and were purchased from
Abcam: Cyclin B1 (cat. no. ab72), cyclin A1 (cat. no. ab53699),
cyclin A2 (cat. no. ab38), cyclin D1 (cat. no. ab6152), cyclin E1
(cat. no. ab3927), cyclin-dependent kinase (CDK)4 (cat. no.
ab108357), CDK1 (cat. no. ab18), CDK6 (cat. no. ab124821), p21
(cat. no. ab109520), p15 (cat. no. ab94688), c-Jun (cat. no.
ab32137), phosphorylated (p)-c-Jun Ser63 (cat. no. ab32385), JunB
(cat. no. ab31421), p-JunB Ser259 (cat. no. ab30628), c-fos (cat.
no. 190289), and p-c-fosSer32 (cat. no. ab79319). The
epithelial-mesenchymal transition (EMT) antibody sampler kit (cat.
no. 9782) was purchased from Cell Signaling Technology, Inc., and
the primary antibodies were diluted at 1:1,000. The other primary
antibodies, including TLR4 monoclonal antibody (cat. no.
66350-1-Ig), MyD88 monoclonal antibody (cat. no. 66660-1-Ig), the
LY96/MD2 polyclonal antibody (cat. no. 11784-1-AP), EGFR rabbit
monoclonal antibody (cat. no. 4405) and p-EGFR (Tyr1068) mouse
monoclonal antibody (cat. no. 2236), were obtained from ProteinTech
Group, Inc. and were diluted at 1:1,000. Following primary antibody
incubation, the membranes were washed three times with TBS-T.
Subsequently, the membranes were incubated with horseradish
peroxidase-labeled secondary antibody (1:10,000; cat. nos. 7076 and
7074; Cell Signaling Technology, Inc.) for 1 h, prior to detection
using an ECL reagent (cat. no. RPN2135; GE Healthcare); all blots
were exposed for the same duration (5 min). α-tubulin was used as
an internal loading control (1:1,000; cat. no. 2125; Cell Signaling
Technology, Inc.). Blots were semi-quantified using Gel-Pro
Analyzer, version 4.0 (Media Cybernetics, Inc.).
MTS assay
Cell proliferation was determined using the MTS
assay. Briefly, 2×103 cells/well were plated in 96-well
culture plates in 150 µl medium; six parallel wells were assigned
to each group, as well as a negative control (without cells). Over
a 5-day period, 30 µl MTS substrate was added to each well every 24
h and incubated for 2 h at 37°C in the dark. Absorbance was
measured at 490 nm for each sample using a plate reader (BMG
Labtech). The half maximal inhibitory concentration
(IC50) values were calculated from survival curves using
the Bliss method (27). All
experiments were performed three times independently.
Flow cytometric analysis
Following treatment with curcumin, cells were
harvested using 0.1% trypsin/EDTA and washed three times with cold
PBS by centrifugation at 300 × g for 5 min. For cell cycle
analysis, cells were fixed in pre-chilled 70% ethanol overnight at
4°C. The fixed cells were collected, washed twice with PBS,
suspended in staining buffer (cat. no. C1052-1; Cell Cycle and
Apoptosis Analysis Kit; Beyotime Institute of Biotechnology)
containing 10 µg/ml propidium iodide (20X, cat. no. C1052-2;
Beyotime Institute of Biotechnology) and 100 µg/ml RNase A (50X,
cat. no. C1052-2; Beyotime Institute of Biotechnology), and then
incubated at 37°C for ≥30 min in the dark to eliminate
intracellular RNA. Analysis was performed on a FACSAria system (BD
Biosciences) and data were analyzed using CellQuest software
(version 5.1; BD Biosciences). All of the samples were assessed
three times.
Colony formation assay
A549 cells were seeded into 6-well plates at a
density of 100 cells/well. The medium containing curcumin (5, 10
and 15 µM) and 10% FBS was changed every 3 days. After 2 weeks,
colonies resulting from the surviving cells were fixed with 3.7%
methanol for 30 min, stained with 10% Giemsa for 15 min at room
temperature and counted. H226 cells were plated into 6-well plates
at a density of 300 cells/well and were cultured in 10% FBS and
medium containing curcumin (5, 10 and 20 µM). Similarly, the medium
was also changed every 3 days. After 3 weeks, the developed
colonies were fixed with 3.7% methanol and stained with Giemsa.
Finally, colonies containing ≥50 cells were counted. Each assay was
performed in triplicate.
Transwell migration assay
The Transwell migration assay was used to determine
the migratory ability of cells. Briefly, A549 cells were grown in
DMEM F12 containing 10% FBS and curcumin (5 and 10 µM). After 48 h,
the cells were harvested by trypsinization and washed once with
PBS. To measure cell migration, 8-µm pore size culture inserts
(Costar; Corning Incorporated) were placed into the wells of
24-well culture plates, separating the upper and the lower
chambers. In the lower chamber, 500 µl DMEM F12 containing 10% FBS
was added, which was used as a chemoattractant. Subsequently,
serum-free medium containing 1×105 cells/well was added
to the upper chamber for migration assays. After 7–8 h of
incubation at 37°C in an atmosphere containing 5% CO2,
the cells on the upper surface of the membrane were removed using a
cotton swab, and the cells that had migrated through the pores were
fixed with 3.7% methanol for 30 min and stained with hematoxylin
for 5 min at room temperature. Finally, cell migration was
determined by calculating the number of migrated cells in three
visual fields per well by light microscopy (Olympus Corporation) at
×100 magnification. Each experiment was performed at least three
times.
Statistical analysis
Data are presented as the mean ± standard deviation.
SPSS software for Windows (version 18.0; SPSS, Inc.) was used for
statistical analysis. The association between TLR4 or MyD88
expression and clinicopathological parameters was analyzed by
χ2 test, and contingency coefficient test was used to
assess the association between TLR4 and MyD88. One-way ANOVA was
used to analyze multiple groups, followed by post hoc analysis
using Student-Newman-Keuls test for comparison between any two
groups or Dunnett's test for comparison among experimental groups
and the control group. Log-rank test was used for survival
analysis. The hazard ratio was determined using Cox regression. The
Kaplan-Meier Plotter (www.kmplot.com) was used to estimate the survival
probability of TLR4 and MyD88 in NSCLC. P<0.05 was considered to
indicate a statistically significant difference.
Results
Curcumin inhibits NSCLC cell
proliferation and colony formation in vitro
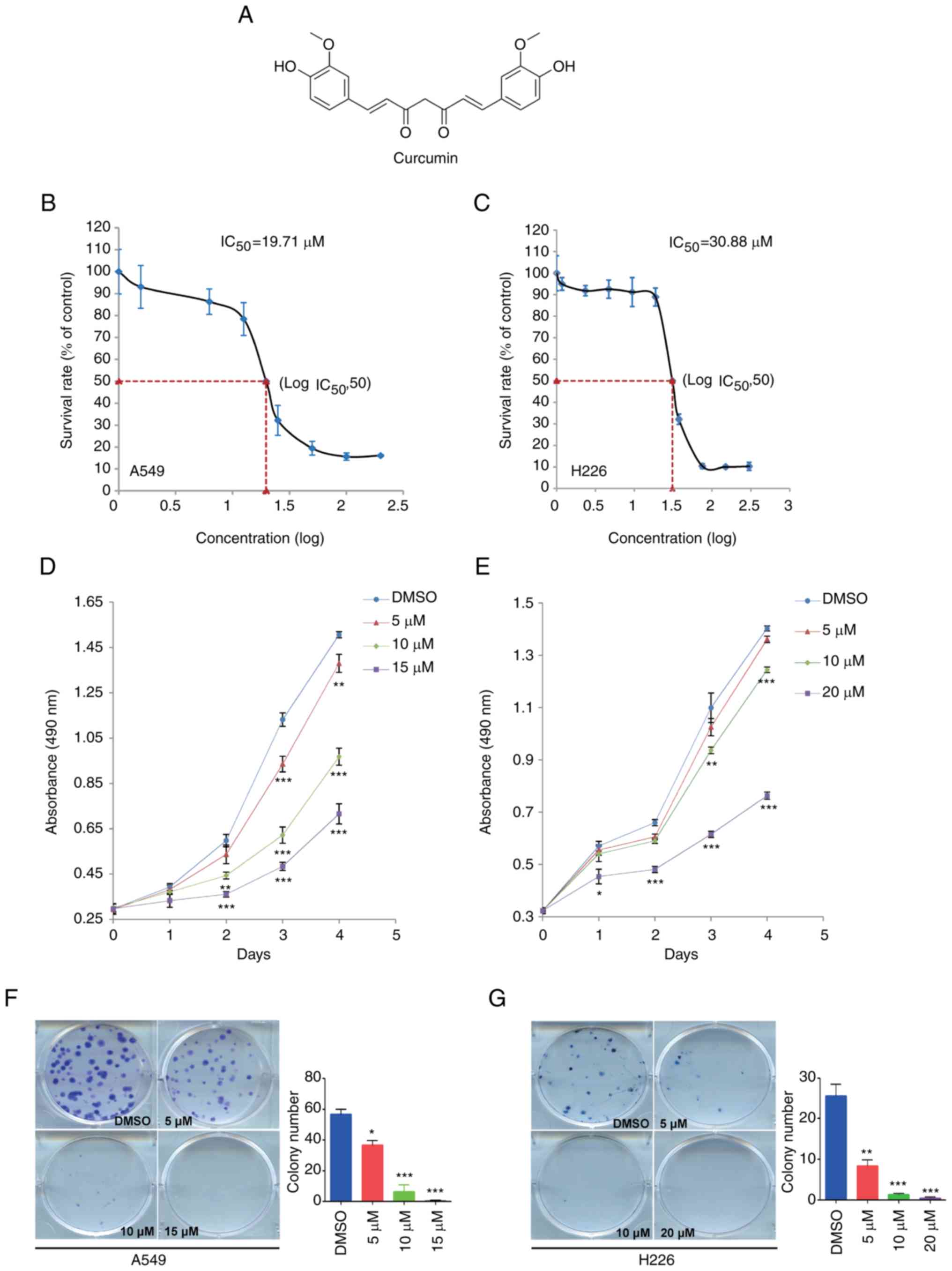
Curcumin is a low molecular weight (368.39 Da)
polyphenol, the chemical structure of which is shown in Fig. 1A. To investigate the effect of
curcumin on NSCLC cell proliferation, the IC50 was
determined in A549 and H226 cells from a dose-response curve after
72 h of treatment. The IC50 values of curcumin were
19.71 and 30.88 µM in A549 and H226 cells, respectively (Fig. 1B and C). According to these results,
15 µM curcumin caused a 20–30% decrease in A549 cell survival rate;
a similar decrease was found in H226 cells in response to 20 µM
curcumin. Subsequently, the MTS assay and colony formation assay
were conducted to evaluate the proliferation of NSCLC cells treated
with 5, 10 and 15 µM, or 5, 10 and 20 µM curcumin. As shown in
Fig. 1D and E, treatment with
curcumin for 4 days significantly reduced cell proliferation in a
dose-dependent manner. Subsequently, the colony formation assay
detected fewer colonies in the curcumin-treated groups compared
with in the DMSO group (Fig. 1F and
G). These data suggested that curcumin may inhibit cell
proliferation in NSCLC.
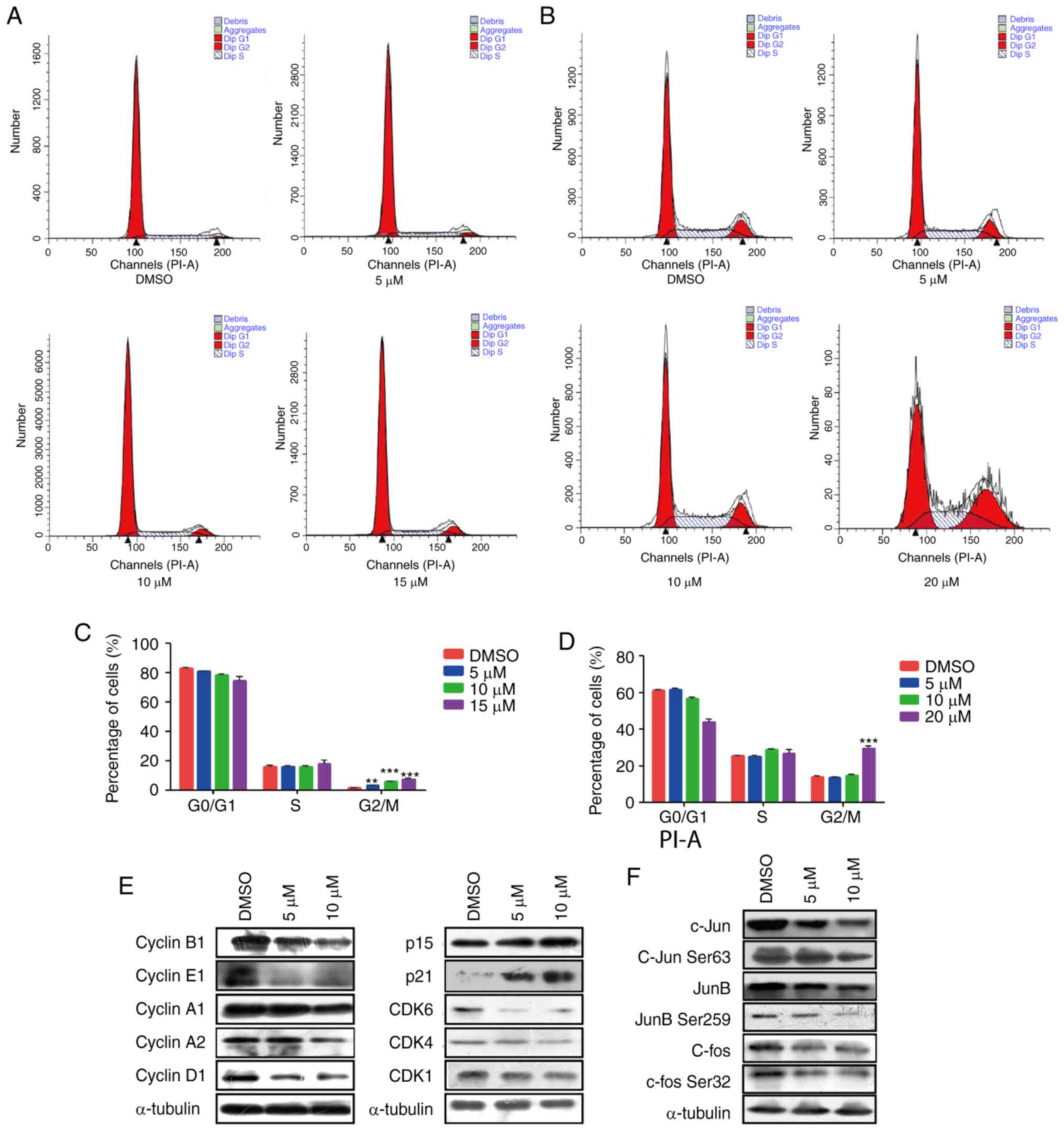
Curcumin induces G2/M phase
cell cycle arrest in NSCLC cell lines
To better understand the mechanism by which curcumin
inhibited cell proliferation, the present study further analyzed
the effects of curcumin on cell cycle distribution in A549 and H226
cells by flow cytometry. The proportion of cancer cells in
G2/M phase was significantly increased in the
curcumin-treated groups compared with in the DMSO group in A549
cells, and the proportion of cells in G0/G1
phase was concomitantly decreased (Fig.
2A and C). However, in H226 cells, 5 and 10 µM curcumin
produced very little change in all phases of the cell cycle.
However, 20 µM curcumin markedly increased the population of cells
in G2/M phase (Fig. 2B and
D). These data indicated that curcumin may modulate
G2/M transition in the cell cycle progression of NSCLC
cells.
The cell cycle process is well known to be
controlled by several types of checkpoints, including cyclins, CDKs
and CDK inhibitors (CDKIs) (28). To
explain the molecular mechanism by which curcumin induced
G2/M phase arrest, the expression of various cell cycle
checkpoints were detected. The results demonstrated that the
protein expression levels of the cell cycle promoters cyclin A1,
cyclin A2, cyclin B1, cyclin E1, cyclin D1 and CDK1 were markedly
decreased, whereas the expression levels of CDKIs, including p15
and p21, were significantly upregulated (Figs. 2E, and S1A
and B). Activator protein-1 (AP-1) has been reported to be
closely associated with the proliferation of tumor cells (29), and activation of c-Jun and JunD
stimulate cyclin D1 (30,31). Therefore, this study examined the
alteration in AP-1 and observed an obvious decrease in the protein
expression levels of c-Jun and c-fos, as well as phosphorylation on
c-Jun Ser63 and c-fos Ser32 (Figs. 2F
and S1C). These findings indicated
that curcumin may slow cell cycle progression by downregulating
interrelated cyclins and CDKs modulated by c-Jun and c-Fos.
However, no significant differences were observed in CDK4, CDK6,
JunB and p-JunB Ser259 expression, thus suggesting that these
signaling molecules were not affected by curcumin.
Briefly, these data indicated that curcumin may
serve a key role in modulating the cell cycle transition in NSCLC
via altering the protein expression levels of c-Jun and c-fos,
thereby further affecting cell cycle regulators, including cyclins
(cyclin A1, cyclin A2, cyclin B1, cyclin E1 and cyclin D1) and
CDK1.
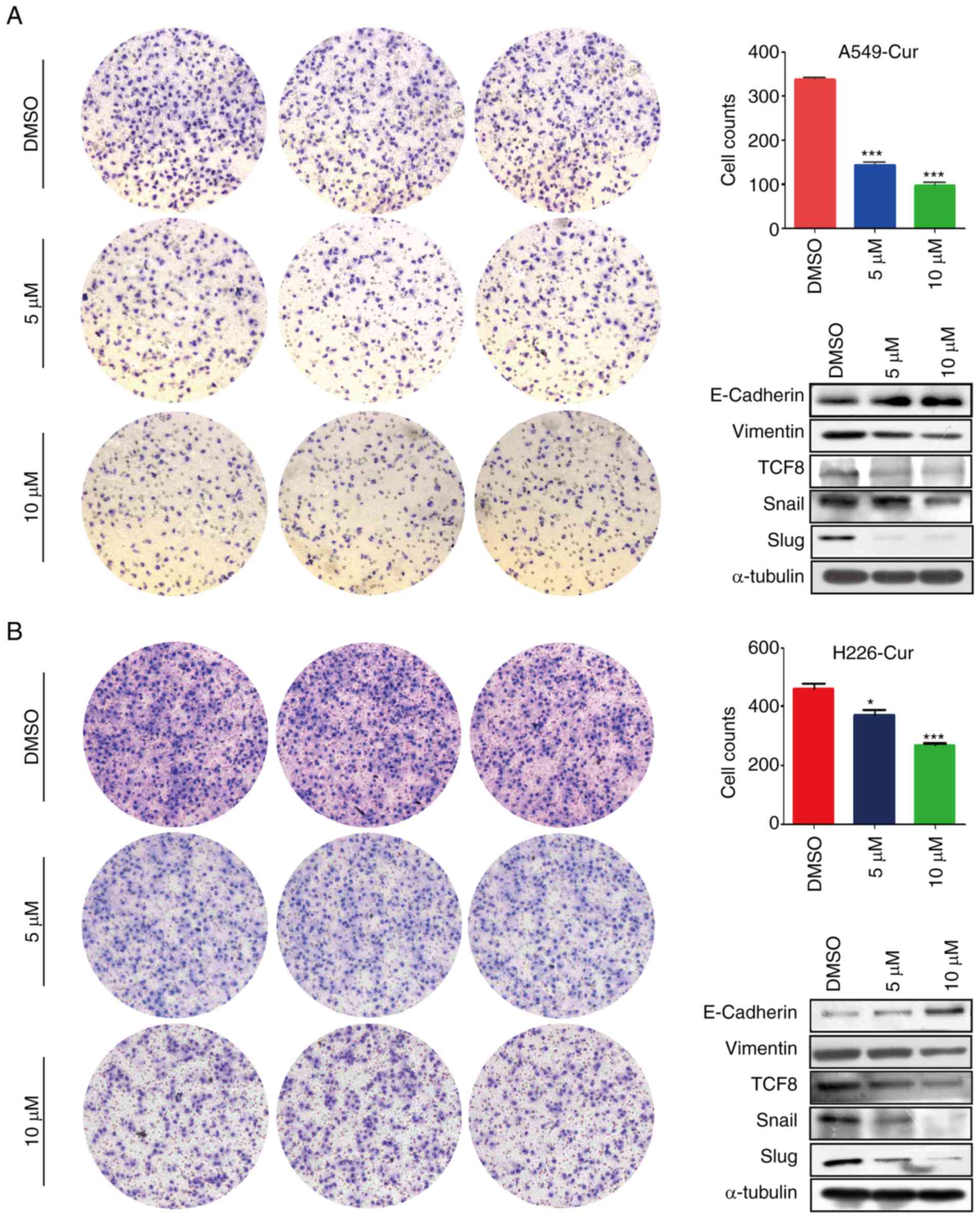
Curcumin suppresses NSCLC cell
migration via EMT-related proteins
A Transwell migration assay was conducted to study
the effects of curcumin on NSCLC cell migration. As shown in
Fig. 3A and B, the number of migrated
A549 and H226 cells in the curcumin-treated group was markedly
decreased compared with in the DMSO-treated group, which indicated
that curcumin may exert anti-metastatic effects. Subsequently,
alterations in EMT-related proteins were detected and E-cadherin
was revealed to be markedly upregulated, whereas vimentin was
markedly downregulated in the curcumin-treated group (Figs. 3 and S2). Furthermore, the expression levels of
transcription factors associated with E-cadherin, including
transcription factor 8 (TCF8), Snail and Slug, were determined. In
line with our expectations, the expression levels of these
transcription factors were markedly decreased in response to
curcumin treatment (Figs. 3 and
S2). These findings suggested that
curcumin inhibited NSCLC cell migration via EMT-related
proteins.
Curcumin downregulates the expression
levels of TLR4/MyD88 and EGFR
TLR4 and EGFR are not only membrane receptors but
also upstream effectors of the AP-1 protein (32). The present study detected MD-2, TLR4,
MyD88 and EGFR protein expression using western blot analysis. It
has previously been reported that curcumin regulates the TLR4/MyD88
signaling pathway by affecting MD-2 expression (33). Consistently, when cultured with
curcumin, A549 and H226 cells exhibited a dose-dependent inhibition
of MD-2, MyD88 and EGFR protein levels. Notably, TLR4 was
significantly inhibited by curcumin in H226 cells; however, only a
non-significant downward trend in TLR4 was detected in response to
curcumin in A549 cells (Figs. 4A and
B, and S3A and B). In addition,
a time-dependent significant decrease in TLR4, MyD88 and EGFR
protein expression was detected following curcumin treatment in
vitro (Figs. 4C and D, and
S3C and D). These results indicated
that curcumin inhibited the TLR4/MyD88 and EGFR signaling pathways
in NSCLC cells.
EGF reverses the inhibitory effects of
curcumin on TLR4/MyD88 in NSCLC cells
Studies regarding the interactions between TLR4 and
EGFR are emerging; however, the underlying mechanism remains
unclear. A lack of a physical association between TLR4 and EGFR has
been demonstrated by coimmunoprecipitation assays in mouse
macrophage cells, and EGFR requires the participation of Lyn kinase
to activate the TLR4 signaling pathway in mouse septic shock
(34). Therefore, whether there is a
synergistic relationship between EGFR and TLR4 in curcumin-treated
NSCLC cells, and whether TLR4/MyD88 is downstream of EGFR, is
currently unknown. This study aimed to determine whether cross talk
existed between TLR4 and EGFR in curcumin-treated NSCLC cells. For
this purpose, A549 and H226 cells were treated successively with
DMSO or curcumin (10 µM) and EGF (100 ng/ml). Western blotting
revealed that EGF significantly promoted MyD88 expression, and EGF
also recovered MyD88 expression in curcumin-treated A549 cells.
Conversely, the inhibitory effect of curcumin on TLR4 was slight
and the difference was not statistically significant in A549 cells.
In addition, no significant difference in TLR4/MyD88 expression was
detected in H226 cells. Furthermore, EGF significantly promoted
p-EGFR expression and could recover p-EGFR levels in
curcumin-treated NSCLC cells (Figs. 4E
and F, and S3E and F). These
results indicated that the TLR4/MyD88 and EGFR pathways may
synergistically regulate cell proliferation and migration in
curcumin-treated NSCLC cells.
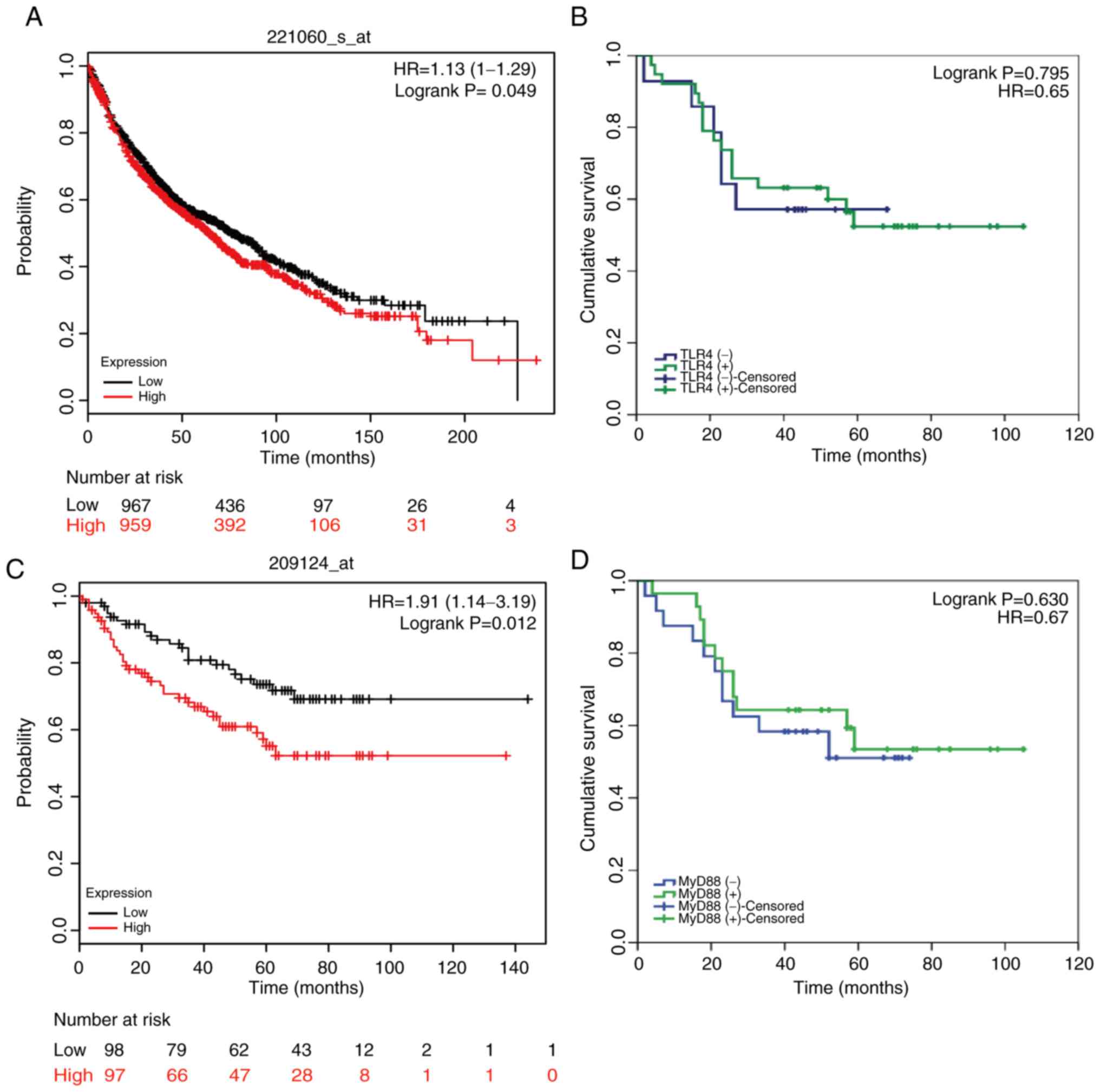
TLR4/MyD88 expression is upregulated
in NSCLC tissues
The present study examined the expression levels of
TLR4/MyD88 in 52 NSCLC tissue samples and 49 benign lung tissue
samples. The panels presented in Fig.
5 are representative images of TLR4 and MyD88 expression in
lung adenocarcinoma and benign lung tissues. TLR4-positive staining
and MyD88-positive staining were confined mainly to the
cytomembrane and cytoplasm in NSCLC tissue (Fig. 5A and B), whereas benign lung tissue
was negatively or weakly stained (Fig. 5C
and D). TLR4-positive staining was detected in 38 out of 52
(73.1%) NSCLC samples but only in 10 out of 49 (20.4%) benign lung
tissue samples (P<0.001; Table I).
Similarly, MyD88-positive staining was detected in 28 out of 52
(53.8%) NSCLC samples but was only found in 12 out of 49 (24.5%)
benign lung tissue samples (P=0.003; Table II). However, there was no association
between TLR4/MyD88 expression and numerous clinical
characteristics, including sex, age, histological type,
differentiation, tumor size, lymph node metastasis and pathological
stage. Notably, the contingency coefficient analysis between TLR4
and MyD88 expression exhibited a significant positive association
(φ=0.308, P=0.026; Table III).
 | Table I.Association between
clinicopathological factors and Toll-like receptor 4 expression in
non-small cell lung cancer. |
Table I.
Association between
clinicopathological factors and Toll-like receptor 4 expression in
non-small cell lung cancer.
| Characteristic | No. of samples | No. of positive
samples (%) | P-value |
|---|
| Histological
type |
|
|
|
| Benign
lung tissues | 49 | 10 (20.4) | <0.001 |
| Lung
cancer tissues | 52 | 38 (73.1) |
|
| Age (years) |
|
|
|
|
<60 | 33 | 23 (69.7) | 0.469 |
|
≥60 | 19 | 15 (79.0) |
|
| Sex |
|
|
|
|
Male | 30 | 20 (66.7) | 0.224 |
|
Female | 22 | 18 (81.8) |
|
| Pathology |
|
|
|
|
Squamous cell carcinoma | 12 | 8 (66.7) | 0.568 |
|
Adenocarcinoma | 40 | 30 (75.0) |
|
| Histopathological
differentiation |
|
|
|
|
Poor | 19 | 11 (57.9) | 0.158 |
|
Moderate | 19 | 15 (78.9) |
|
|
High | 14 | 12 (85.7) |
|
| Tumor size |
|
|
|
|
T1/T2 | 45 | 33 (73.3) | 0.916 |
|
T3/T4 | 7 | 5 (71.4) |
|
| Lymph node
metastasis |
|
|
|
| N0 | 21 | 16 (76.2) | 0.677 |
|
N1-2 | 31 | 22 (71.0) |
|
| Pathological
stage |
|
|
|
|
I–IIB | 35 | 24 (68.6) | 0.293 |
|
IIIA-IIIB | 17 | 14 (82.4) |
|
| Table II.Association between the
clinicopathological factors and MyD88 expression in non-small cell
lung cancer. |
Table II.
Association between the
clinicopathological factors and MyD88 expression in non-small cell
lung cancer.
| Characteristic | No. of samples | No. of positive
samples (%) | P-value |
|---|
| Histological
type |
|
|
|
| Benign
lung tissues | 49 | 12 (24.5) | 0.003 |
| Lung
cancer tissues | 52 | 28 (53.8) |
|
| Age, years |
|
|
|
|
<60 | 33 | 16 (48.5) | 0.307 |
|
≥60 | 19 | 12 (63.2) |
|
| Sex |
|
|
|
|
Male | 30 | 18 (60.0) | 0.299 |
|
Female | 22 | 10 (45.5) |
|
| Pathology |
|
|
|
|
Squamous cell carcinoma | 12 | 8 (66.7) | 0.310 |
|
Adenocarcinoma | 40 | 20 (50.0) |
|
| Histopathological
differentiation |
|
|
|
|
Poor | 19 | 8 (42.1) | 0.411 |
|
Moderate | 19 | 12 (63.2) |
|
|
High | 14 | 8 (57.1) |
|
| Tumor size |
|
|
|
|
T1/T2 | 45 | 24 (53.3) | 0.851 |
|
T3/T4 | 7 | 4 (57.1) |
|
| Lymph node
metastasis |
|
|
|
| N0 | 21 | 10 (47.6) | 0.458 |
|
N1-2 | 31 | 18 (58.1) |
|
| Pathological
stage |
|
|
|
|
I–IIB | 35 | 19 (54.3) | 0.927 |
|
IIIA-IIIB | 17 | 9 (52.9) |
|
| Table III.Association between TLR4 and MyD88
expression in non-small cell lung cancer tissues. |
Table III.
Association between TLR4 and MyD88
expression in non-small cell lung cancer tissues.
|
| MyD88 |
|
|
|---|
|
|
|
|
|
|---|
| TLR4 | + | − | P-value | φ |
|---|
| + | 24 | 14 | 0.026 | 0.308 |
| − | 4 | 10 |
|
|
TLR4/MyD88 expression and EGFR
expression may be relevant to the survival of patients with
NSCLC
The follow-up period for all 52 patients (I–IIB
stage) enrolled in this research began at the discharge date until
September 2015. At the end of the follow-up period, six patients
were lost to follow-up and 24 had died, leaving 22 survivors. By
differentiating patients into positive or negative TLR4/MyD88
staining groups, the prognostic effect of TLR4/MyD88 on the overall
survival of patients with NSCLC was analyzed. Kaplan-Meier curve
analysis demonstrated that there was no association between
TLR4/MyD88 staining status and overall survival (OS) of patients
with NSCLC (n=52; P=0.795 and 0.630, respectively; Fig. 6B and D). However, this result differed
from the results of Kaplan-Meier data analysis on a larger sample
size. As shown in Fig. 6A and C,
these results suggested that TLR4/MyD88 expression levels were
significantly associated with OS in patients with NSCLC. Of the
possible reasons for this disagreement, one may be the small sample
size (n=52). Similarly, the Kaplan-Meier Plotter database was used
to analyze the association between EGFR expression and OS. The
results shown in Fig. S4 suggested
that high EGFR expression was closely associated with poor
prognosis.
Discussion
The currently available treatments for NSCLC have
poor curative effects, which are an obstacle to therapy; therefore,
the development of safe and effective therapies is urgently
required to improve treatment. Tumorigenesis is usually accompanied
by aberrant alterations in the expression levels of gene involved
in cell proliferation and metastasis. As a promising anticancer
agent, curcumin targets multiple signaling pathways and regulates
the expression of genes related to tumor signal transduction
(35).
It has previously been suggested that TLR4 is one of
the key factors in tumor growth regulation, independent of its
immune activity, in breast cancer and that the oncogene is closely
associated with the outcome of TP53 wild-type breast cancer
(36). Another study in breast cancer
indicated that TLR4 activation by lipopolysaccharide (LPS) triggers
the Akt/glycogen synthase kinase-3β/β-catenin signaling pathway,
leading to breast cancer migration and metastasis (37). In colorectal cancer, TLR4 expression,
as determined by IHC staining, is strongly associated with survival
and recurrence assessed by survival analysis and prediction
(38). In addition, the demethylation
of TLR4 by Sp1 promotes activation of the NF-κB pathway in gastric
cancer (39). Taken together, TLR4
may serve a key role in carcinogenesis.
The present study used IHC to confirm that
TLR4/MyD88 expression was upregulated in NSCLC tissues compared
with in benign lung tissues. The results indicated that TLR4 may be
a biomarker for NSCLC diagnosis. Due to the small sample size, no
association was detected between TLR4 and clinical characteristics.
In addition, the TLR4-positive rate was similar in N1-2 tissues
compared with in N0 tissues, with no significant difference; more
cases should be assessed in future to test for a difference.
Nevertheless, lower TLR4/MyD88 expression levels were markedly
associated with a longer survival rate in patients with NSCLC, as
determined using Kaplan-Meier database analysis. Survival
prediction in oral squamous cell carcinoma previously demonstrated
that high TLR4 and MyD88 expression likely results in the
production of proinflammatory cytokines, chemokines and growth
factors; suppression of TLR4 could enhance anticancer immunity and
lead to a good prognosis (40).
EMT has been reported to be very important in
metastasis and is closely linked to E-cadherin. Previous studies
have reported that the TLR4/MyD88 and EGFR signaling pathways are
associated with cancer metastasis (41–43), and
several lines of evidence have indicated that TLR4 and EGFR promote
metastasis of several types of cancer by affecting the EMT process
(44–46). Furthermore, a previous study revealed
that curcumin liposome exerts antitumor effects by affecting
proliferation- and apoptosis-related molecules in rabbits (47). In addition, curcumin inhibits lung
cancer growth through the regulation of vascular endothelial growth
factor signaling in mice (48). This
study demonstrated that curcumin could, to a large extent, block
migration, upregulate the expression of E-cadherin and suppress the
expression levels of vimentin. Furthermore, the expression of
transcription factors associated with E-cadherin was markedly
decreased. Combining these data and the previous results, it was
hypothesized that curcumin could inhibit EMT via downregulating the
TLR4 and EGFR signaling pathways, and increasing E-cadherin
expression, in order to control NSCLC cell migration.
A previous study confirmed that curcumin can
directly bind to MD-2, which is the upstream regulator of TLR4,
similar to LPS, and form a curcumin-MD-2/TLR4 complex (49). In vivo, the curcumin analog
L48H37 improves survival and protects lungs against LPS-induced
injury in septic mice via its interaction with MD-2 (50). In this study, a series of functional
experiments were performed and the results demonstrated an
anti-proliferative effect in NSCLC cells treated with curcumin.
Subsequently, the anti-proliferative mechanism of curcumin was
investigated. Curcumin induced G2/M phase arrest by
controlling certain checkpoints of the cell cycle, including cyclin
A1, cyclin A2, cyclin B1, cyclin D1, cyclin E1, CDK1, p15 and p21.
Activation of the AP-1 protein upregulates cell cycle promoters,
such as cyclins and CDKs, and suppresses cell cycle inhibitors such
as CDKIs (51). The present results
demonstrated that the expression levels of AP-1 were markedly
decreased following curcumin treatment, which explains the decrease
in cyclin A1, cyclin A2, cyclin B1, cyclin D1, cyclin E1 and CDK1,
and the increase in p15 and p21; these results indicated that
curcumin may serve a key role in modulating the cell cycle
transition in NSCLC via altering the levels of JUN family proteins,
thereby further affecting cell cycle regulators, particularly
cyclins, CDKs and CDKIs.
The present study demonstrated that curcumin
significantly inhibited the expression of TLR4/MyD88 and EGFR in a
dose- and time-dependent manner. Furthermore, the inhibitory action
could be reversed by EGF in curcumin-treated NSCLC cells. The
results of a Transwell migration assay provided evidence of the
anti-migratory effect of curcumin, and curcumin was revealed to
affect the expression of metastasis-related proteins. In addition,
an analysis was performed to determine the association between
lymph node metastasis and TLR4/MyD88 expression; however, no
statistical significance was determined. The possible reason for
this discrepancy may be the small sample size. Previous studies
have reported that TLR4/MyD88 cooperates with EGFR and takes part
in cancer metastases (52,53). Therefore, it was hypothesized that
curcumin may affect cell proliferation and migration in NSCLC
through a synergistic effect on the TLR4/MyD88 and EGFR pathways.
However, there was a limitation to the study. The bioavailability
of curcumin was very low, which limited the progress of animal
experiments; further studies aim to conduct in vivo
experiments once the bioavailability problem has been resolved.
In conclusion, this study reported that TLR4/MyD88
expression was significantly upregulated in NSCLC. Curcumin
treatment significantly reduced the proliferation and migration of
NSCLC cells, possibly through TLR4/MyD88-EGFR-mediated inhibition
of AP-1 protein and suppression of the EMT process. In the present
study, preliminary evidence of curcumin-induced synergistic
regulation of the TLR4/MyD88 and EGFR signaling pathways was
obtained; however, there remains a lack of direct evidence;
therefore, further studies are required.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
This study was supported by the China Postdoctoral
Science Foundation (grant no. 2017M613008), the National Natural
Science Foundation of China (grant nos. U1502222, 81702295 and
81602029), and the Yunnan Province Applied Foundation Project
(grant no. 2018FB138).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
LZ, XT, QF, CG, RL, ZL, YZ, HT, QL, ML, HH, BZ, ZL
and CL performed the research. XS and RL designed the study; LZ,
RL, XT and QF performed the statistical analysis; QF, LZ, XT and XS
wrote the study. All authors have read and approved the final
manuscript.
Ethics approval and consent to
participate
This study was approved by the Ethics Committees of
The Third Affiliated Hospital of Kunming Medical University, and
informed consent was obtained from the patients.
Patient consent for publication
Not applicable.
Competing interests
The authors declare no that they have no competing
interests.
References
|
1
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China,
2015. CA Cancer J Clin. 66:115–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2017. CA Cancer J Clin. 67:7–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Torre LA, Siegel RL and Jemal A: Lung
cancer statistics. Adv Exp Med Biol. 893:1–19. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kiuchi F, Goto Y, Sugimoto N, Akao N,
Kondo K and Tsuda Y: Nematocidal activity of turmeric: Synergistic
action of curcuminoids. Chem Pharm Bull (Tokyo). 41:1640–1643.
1993. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Shishodia S, Sethi G and Aggarwal BB:
Curcumin: Getting back to the roots. Ann NY Acad Sci. 1056:206–217.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Shishodia S: Molecular mechanisms of
curcumin action: Gene expression. Biofactors. 39:37–55. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Aggarwal BB and Sung B: Pharmacological
basis for the role of curcumin in chronic diseases: An age-old
spice with modern targets. Trends Pharmacol Sci. 30:85–94. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Aggarwal BB and Harikumar KB: Potential
therapeutic effects of curcumin, the anti-inflammatory agent,
against neurodegenerative, cardiovascular, pulmonary, metabolic,
autoimmune and neoplastic diseases. Int J Biochem Cell Biol.
41:40–59. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kuttan R, Bhanumathy P, Nirmala K and
George MC: Potential anticancer activity of turmeric (Curcuma
longa). Cancer Lett. 29:197–202. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kuttan R, Sudheeran PC and Josph CD:
Turmeric and curcumin as topical agents in cancer therapy. Tumori.
73:29–31. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Mishra A, Liu S, Sams GH, Curphey DP,
Santhanam R, Rush LJ, Schaefer D, Falkenberg LG, Sullivan L,
Jaroncyk L, et al: Aberrant overexpression of IL-15 initiates large
granular lymphocyte leukemia through chromosomal instability and
DNA hypermethylation. Cancer Cell. 22:645–655. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Demaria S, Pikarsky E, Karin M, Coussens
LM, Chen YC, El-Omar EM, Trinchieri G, Dubinett SM, Mao JT, Szabo
E, et al: Cancer and inflammation: Promise for biologic therapy. J
Immunother. 33:335–351. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Katoh H, Wang D, Daikoku T, Sun H, Dey SK
and Dubois RN: CXCR2-expressing myeloid-derived suppressor cells
are essential to promote colitis-associated tumorigenesis. Cancer
Cell. 24:631–644. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang L, Zhang H, Rodriguez S, Cao L,
Parish J, Mumaw C, Zollman A, Kamoka MM, Mu J, Chen DZ, et al:
Notch-dependent repression of miR-155 in the bone marrow niche
regulates hematopoiesis in an NF-κB-dependent manner. Cell Stem
Cell. 15:51–65. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Garza-Gonzalez E, Bosques-Padilla FJ,
Mendoza-Ibarra SI, Flores-Gutierrez JP, Maldonado-Garza HJ and
Perez-Perez GI: Assessment of the toll-like receptor 4 Asp299Gly,
Thr399Ile and interleukin-8-251 polymorphisms in the risk for the
development of distal gastric cancer. BMC Cancer. 7:702007.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
He W, Liu Q, Wang L, Chen W, Li N and Cao
X: TLR4 signaling promotes immune escape of human lung cancer cells
by inducing immunosuppressive cytokines and apoptosis resistance.
Mol Immunol. 44:2850–2859. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Cheng I, Plummer SJ, Casey G and Witte JS:
Toll-like receptor 4 genetic variation and advanced prostate cancer
risk. Cancer Epidemiol Biomarkers Prev. 16:352–355. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chicoine MR, Zahner M, Won EK, Kalra RR,
Kitamura T, Perry A and Higashikubo R: The in vivo antitumoral
effects of lipopolysaccharide against glioblastoma multiforme are
mediated in part by Toll-like receptor 4. Neurosurgery. 60:372–381.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Semlali A, Reddy Parine N, Arafah M,
Mansour L, Azzi A, Al Shahrani O, Al Amri A, Shaik JP, Aljebreen
AM, Alharbi O, et al: Expression and polymorphism of toll-like
receptor 4 and effect on NF-κB mediated inflammation in colon
cancer patients. PLoS One. 11:e01463332016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Köe M: GPCRs and EGFR-Cross-talk of
membrane receptors in cancer. Bioorg Med Chem Lett. 27:3611–3620.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lev-Ari S, Starr A, Vexler A, Karaush V,
Loew V, Greif J, Fenig E, Aderka D and Ben-Yosef R: Inhibition of
pancreatic and lung adenocarcinoma cell survival by curcumin is
associated with increased apoptosis, down-regulation of COX-2 and
EGFR and inhibition of Erk1/2 activity. Anticancer Res.
26:4423–4430. 2006.PubMed/NCBI
|
|
23
|
De S, Zhou H, DeSantis D, Croniger CM, Li
X and Stark GR: Erlotinib protects against LPS-induced endotoxicity
because TLR4 needs EGFR to signal. Proc Natl Acad Sci USA.
112:9680–9685. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Basu S, Pathak SK, Chatterjee G, Pathak S,
Basu J and Kundu M: Helicobacter pylori protein HP0175
transactivates epidermal growth factor receptor through TLR4 in
gastric epithelial cells. J Biol Chem. 283:32369–32376. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hsu D, Fukata M, Hernandez YG, Sotolongo
JP, Goo T, Maki J, Hayes LA, Ungaro RC, Chen A, Breglio KJ, et al:
Toll-like receptor 4 differentially regulates epidermal growth
factor-related growth factors in response to intestinal mucosal
injury. Lab Invest. 90:1295–1305. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Edge SB and Compton CC: The American Joint
Committee on Cancer: The 7th edition of the AJCC cancer staging
manual and the future of TNM. Ann Surg Oncol. 17:1471–1474. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Guo H, Luo H, Yuan H, Xia Y, Shu P, Huang
X, Lu Y, Liu X, Keller ET, Sun D, et al: Litchi seed extracts
diminish prostate cancer progression via induction of apoptosis and
attenuation of EMT through Akt/GSK-3β signaling. Sci Rep.
7:416562017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Barnum KJ and O'Connell MJ: Cell cycle
regulation by checkpoints. Methods Mol Biol. 1170:29–40. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Karin M, Liu ZG and Zandi E: AP-1 function
and regulation. Curr Opin Cell Biol. 9:240–246. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Schwabe RF, Bradham CA, Uehara T, Hatano
E, Bennett BL, Schoonhoven R and Brenner DA: c-Jun-N-terminal
kinase drives cyclin D1 expression and proliferation during liver
regeneration. Hepatology. 37:824–832. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Toualbi-Abed K, Daniel F, Güller MC,
Legrand A, Mauriz JL, Mauviel A and Bernuau D: Jun D cooperates
with p65 to activate the proximal kappaB site of the cyclin D1
promoter: Role of PI3K/PDK-1. Carcinogenesis. 29:536–543. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lv X, Wang H, Su A, Xu S and Chu Y: Herpes
simplex virus type 2 infection triggers AP-1 transcription activity
through TLR4 signaling in genital epithelial cells. Virol J.
15:1732018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Gradisar H, Keber MM, Pristovsek P and
Jerala R: MD-2 as the target of curcumin in the inhibition of
response to LPS. J Leukoc Biol. 82:968–974. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chattopadhyay S, Veleeparambil M, Poddar
D, Abdulkhalek S, Bandyopadhyay SK, Fensterl V and Sen GC: EGFR
kinase activity is required for TLR4 signaling and the septic shock
response. EMBO Rep. 16:1535–1547. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Shehzad A, Lee J and Lee YS: Curcumin in
various cancers. Biofactors. 39:56–68. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Haricharan S and Brown P: TLR4 has a
TP53-dependent dual role in regulating breast cancer cell growth.
Proc Natl Acad Sci USA. 112:E3216–E3225. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Li J, Yin J, Shen W, Gao R, Liu Y, Chen Y,
Li X, Liu C, Xiang R and Luo N: TLR4 promotes breast cancer
metastasis via Akt/GSK3β/β-catenin pathway upon LPS stimulation.
Anat Rec (Hoboken). 300:1219–1229. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Sussman DA, Santaolalla R, Bejarano PA,
Garcia-Buitrago MT, Perez MT, Abreu MT and Clarke J: In silico and
ex vivo approaches identify a role for toll-like receptor 4 in
colorectal cancer. J Exp Clin Cancer Res. 33:452014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Kim TW, Lee SJ, Oh BM, Lee H, Uhm TG, Min
JK, Park YJ, Yoon SR, Kim BY, Kim JW, et al: Epigenetic
modification of TLR4 promotes activation of NF-κB by regulating
methyl-CpG-binding domain protein 2 and Sp1 in gastric cancer.
Oncotarget. 7:4195–4209. 2016.PubMed/NCBI
|
|
40
|
Sun Z, Luo Q, Ye D, Chen W and Chen F:
Role of toll-like receptor 4 on the immune escape of human oral
squamous cell carcinoma and resistance of cisplatin-induced
apoptosis. Mol Cancer. 11:332012. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Wang EL, Qian ZR, Nakasono M, Tanahashi T,
Yoshimoto K, Bando Y, Kudo E, Shimada M and Sano T: High expression
of Toll-like receptor 4/myeloid differentiation factor 88 signals
correlates with poor prognosis in colorectal cancer. Br J Cancer.
102:908–915. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Wu K, Zhang H, Fu Y, Zhu Y, Kong L, Chen
L, Zhao F, Yu L and Chen X: TLR4/MyD88 signaling determines the
metastatic potential of breast cancer cells. Mol Med Rep.
18:3411–3420. 2018.PubMed/NCBI
|
|
43
|
Miyamoto Y, Suyama K and Baba H: Recent
advances in targeting the EGFR signaling pathway for the treatment
of metastatic colorectal cancer. Int J Mol Sci. 18(pii): E7522017.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Radisky DC: Epithelial-mesenchymal
transition. J Cell Sci. 118:4325–4326. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
He Z, Deng R, Huang X, Ni Y, Yang X, Wang
Z and Hu Q: Lipopolysaccharide enhances OSCC migration by promoting
epithelial-mesenchymal transition. J Oral Pathol Med. 44:685–692.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Ye K, Chen QW, Sun YF, Lin JA and Xu JH:
Loss of BMI-1 dampens migration and EMT of colorectal cancer in
inflammatory microenvironment through TLR4/MD-2/MyD88-mediated
NF-κB signaling. J Cell Biochem. 119:1922–1930. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Zhang X, Dai F, Chen J, Xie X, Xu H, Bai
C, Qiao W and Shen W: Antitumor effect of curcumin liposome after
transcatheter arterial embolization in VX2 rabbits. Cancer Biol
Ther. 20:642–652. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Li X, Ma S, Yang P, Sun B, Zhang Y, Sun Y,
Hao M, Mou R and Jia Y: Anticancer effects of curcumin on nude mice
bearing lung cancer A549 cell subsets SP and NSP cells. Oncol Lett.
16:6756–6762. 2018.PubMed/NCBI
|
|
49
|
Wang Z, Chen G, Chen L, Liu X, Fu W, Zhang
Y, Li C, Liang G and Cai Y: Insights into the binding mode of
curcumin to MD-2: Studies from molecular docking, molecular
dynamics simulations and experimental assessments. Mol Biosyst.
11:1933–1938. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Wang Y, Shan X, Dai Y, Jiang L, Chen G,
Zhang Y, Wang Z, Dong L, Wu J, Guo G and Liang G: Curcumin analog
L48H37 prevents lipopolysaccharide-induced TLR4 signaling pathway
activation and sepsis via targeting MD2. J Pharmacol Exp Ther.
353:539–550. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Liu Y, Lu C, Shen Q, Munoz-Medellin D, Kim
H and Brown PH: AP-1 blockade in breast cancer cells causes cell
cycle arrest by suppressing G1 cyclin expression and reducing
cyclin-dependent kinase activity. Oncogene. 23:8238–8246. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Gong WJ, Liu JY, Yin JY, Cui JJ, Xiao D,
Zhuo W, Luo C, Liu RJ, Li X, Zhang W, et al: Resistin facilitates
metastasis of lung adenocarcinoma through the
TLR4/Src/EGFR/PI3K/NF-κB pathway. Cancer Sci. 109:2391–2400. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Thuringer D, Hammann A, Benikhlef N,
Fourmaux E, Bouchot A, Wettstein G, Solary E and Garrido C:
Transactivation of the epidermal growth factor receptor by heat
shock protein 90 via Toll-like receptor 4 contributes to the
migration of glioblastoma cells. J Biol Chem. 286:3418–3428. 2011.
View Article : Google Scholar : PubMed/NCBI
|