Introduction
Lung cancer, a malignant disease of the respiratory
system, is the leading cause of cancer-related mortality (1). According to medical convention, lung
cancer is classified into two main categories: small cell lung
cancer (SCLC) and non-small cell lung cancer (NSCLC) (2). Among the subtypes of NSCLC, lung
squamous cell carcinoma and lung adenocarcinoma (LUAD) are the most
prevalent, with the latter the most frequently occurring
histological type in non-smokers (2). LUAD is often diagnosed at an advanced
or metastatic stage, resulting in the loss of early treatment
opportunities (3). However,
advancements in surgical treatment, including enhanced staging and
video-assisted thoracic surgery, have led to improved survival
rates across all stages of NSCLC (4). Although much research has been
conducted into LUAD therapy, the 5-year survival rate and prognosis
of patients with LUAD remain suboptimal (4). Therefore, identification of novel
therapeutic interventions is an urgent need.
Activin A, originally extracted from porcine
follicular fluid, belongs to the superfamily of transforming growth
factors (TGF-β) and induces pituitary cells to secrete follicle
stimulating hormone (5). Similar to
other TGF-β superfamily members, activin A initiates its signaling
cascade by binding to activin receptor type II (ActRII), and then
recruits activin receptor type I (ActRI) to form a heterodimer that
results in the activation of Smad2 and Smad3 proteins. These
proteins then, form a complex with Smad4 (Smad2/3/4), which
translocates to the nucleus and drives downstream target genes,
leading to biological effects such as cell proliferation, apoptosis
and migration (6). Studies have
revealed that activin A-overexpression in diverse tissues is
involved in tumor progression in various cancers, including LUAD,
pancreatic ductal adenocarcinoma, breast cancer and oral squamous
cell carcinoma (7–10).
Numerous genetic and environmental insults impede
the ability of cells to properly fold and post-translationally
modify secretory and transmembrane proteins in the endoplasmic
reticulum (ER), leading to a buildup of misfolded proteins in this
organelle (11), a phenomenon
referred to as ER stress. If ER stress persists chronically at high
levels, the unfolded protein response (UPR) transforms into an
alternate signaling platform referred to as the terminal UPR that
actively promotes cell death (11,12).
ER stress and sustained UPR signaling have been well documented in
numerous diseases, including diabetes, neurological diseases,
pulmonary fibrosis, cancer (multiple myeloma, breast) and heart
disease (11). However, triggering
excessive ER stress can induce tumor cell death, which may become a
therapeutic strategy against tumor growth (13). Although research has demonstrated
that activin A plays a neuroprotective role in ER stress-mediated
apoptotic and autophagic rat pheochromocytoma PC12 cell death
(14), few studies have examined
the role of activin A and ER stress in other tumor cells.
Prior research has documented that a high
concentration of activin A inhibits the growth of LUAD cells
(7) and can result in apoptosis of
myeloma NS-1 cells through the ER stress pathway (6). Nevertheless, it remains unclear
whether activin A can mediate apoptosis of human LUAD cells via the
ER stress pathway. Through gene data comparison, it was found that
the expression of ActRIIA and non-classical pathway signaling
proteins improved the survival of patients with LUAD. Consequently,
the human LUAD cell line A549 was used as the experimental model to
explore the potential functions of activin A and identify its
underlying mechanisms of inducing apoptosis in LUAD cells.
Materials and methods
Cell culture
The human LUAD cell line A549 (https://www.cellosaurus.org/CVCL_0023;
cat. no. CCL-185; American Type Culture Collection) was cultured in
RPMI-1640 (cat. no. 11875093; Thermo Fisher Scientific, Inc.)
supplemented with 10% fetal bovine serum (FBS) (cat. no. C04001;
Shanghai VivaCell Biosciences, Ltd.) and 1% penicillin-streptomycin
at 37°C in a humidified incubator with 5% CO2. The cells
were passaged every two days.
Cell Counting Kit-8 (CCK-8) assay
A549 cells (1.5×104 cells/well) were
seeded into a 96-well plate and incubated in 2% FBS-RPMI-1640
containing activin A (0–40 ng/ml) (cat. no. 338-AC; R&D
Systems, Inc.) for 24 and 48 h, respectively. Next, 10 µl of CCK-8
reagent (cat. no. GK10001; GLPBIO) was added to the culture medium
of each well, and the cells were incubated for 2 h at 37°C in 5%
CO2. The absorbance values (A value) were obtained at
the wavelengths of 450 and 650 nm. Each experiment was performed in
triplicate compound wells and repeated three times. The percentage
(%) of cell viability was calculated as follows: (%)=(sample well A
value/control well A value) ×100%.
Real-time cell analysis (RTCA)
RTCA is a powerful technology for detecting cell
fate (15). The RTCA instrument
(xCELLigence RTCA S16; ACEA Bioscience, Inc.; Agilent) was used to
analyze the proliferation property of A549 cells. The experimental
method was as previously described (15). In brief, 50 µl of cell-free culture
medium (2% FBS-RPMI-1640) was added to the well of an E16
×CELLigence microtiter plate to test the background impedance of
each well for 1 min. Subsequently, A549 cells (1×104
cells in 50 µl culture medium) were added to the plate and cultured
at 37°C in 5% CO2 for ~2 h, and then treated with
different concentrations of activin A (0–40 ng/ml) for 48 h. Cells
were monitored at 15 min intervals. Each experiment was set up with
double compound holes and repeated thrice. The cell index
indirectly represented cell activity.
5-bromo-2′-deoxyuridine (BrdU), Elisa
method for cell proliferation
A549 cells (1.5×104 cells/well) were
seeded into a 96-well plate and incubated in 2% FBS-RPMI-1640
containing activin A (0–40 ng/ml) for 24 h. In accordance with a
cell proliferation ELISA protocol (cat. no. 11647229001; Merck
KGaA), 10 µl of BrdU labeling reagent was added, and incubation was
continued for 2 h. After discarding the supernatants, 200 µl/well
of FixDenat reagent was added for 30 min to fix the cells and
denature the DNA. After discarding the supernatants, 100 µl/well of
anti-BrdU-POD was added and followed by incubation for 90 min.
After discarding the supernatants and washing, 100 µl/well of
substrate solution was added and incubated for 30 min. The
absorbance values (A value) were obtained at the wavelengths of 370
and 492 nm. Each experiment was performed in triplicate compound
wells. The percentage (%) of cell proliferation was calculated as
follows: (%)=(sample well A value/control well A value) ×100%.
Hoechst 33342 staining
A549 cells (1.5×104 cells/well) were
seeded into a 96-well plate and cultured in 2% FBS-RPMI-1640
culture medium containing activin A (0–40 ng/ml). Cells were then
fixed with 4% paraformaldehyde for 20 min and stained with 10 µg/ml
Hoechst 33342 solution (cat. no. 14533; Sigma-Aldrich; Merck KGaA)
in the dark for 10 min at room temperature and observed under an
inverted fluorescence microscope (IX71; Olympus Corporation).
Experiments were repeated three times.
Flow cytometric analysis of cell
apoptosis
The experiment was performed as previously described
(6). A549 cells (1×105
cells/well) were seeded in a 12-well plate and cultured in 2%
FBS-RPMI-1640 culture medium containing activin A (0–40 ng/ml) for
24 h. Thereafter, the cells were collected and suspended in 100 µl
of 1X Annexin V binding buffer. Next, 1 µl of Annexin
V-YF®488 and 1 µl of PI (cat. no. Y6002; Suzhou Yuheng
Biotechnology Co., Ltd.; www.uebio.com) were used to mark the cells for 15 min
in the dark. Finally, the labeled cells were analyzed using flow
cytometry (guava easyCyte HT; Cytek Biosciences). FlowJo
(FlowJo_v10.6.2; FlowJo LLC) was used to collect and analyze the
data to obtain the percentage of fluorescent cells. The experiments
were repeated three times. BAPTA-AM (2.5 µM; cat. no. HY-100545),
calcium agonist ionomycin (500 nm; cat. no. HY-13434; both from
MedChemExpress) and ERK inhibitor FR180204 (10 µM; cat. no.
abs810541; Absin Bioscience, Inc.) were also used before assessment
of cell apoptosis.
Calcium influx assay
A549 cells were incubated in 2% FBS-RPMI-1640 medium
with 4 µmol/l Fluo-4 (cat. no. F14201; Thermo Fisher Scientific,
Inc.) and were protected from light for 40 min at 37°C. The cells
were then washed twice with 2% FBS-RPMI-1640 medium and seeded into
a 24-well plate. Calcium fluorescence signaling was detected using
a BioTek Cytation5 cell imager multimode reader (Agilent
Technologies, Inc.) after the cells were adherent. The baseline
fluorescence signal (F0) was recorded every 0.25 sec for
a total of 3 sec. Next, the cells were treated with 2%
FBS-RPMI-1640 and activin A 20 ng/ml, respectively, and the
fluorescence signal (F) was immediately detected on the machine for
another 3 sec. The experimental data were exported from BioTek
Cytation5 software, and the normalized fluorescence intensity was
represented by F/F0. Experiments were repeated three
times.
Reverse transcription-polymerase chain
reaction (RT-PCR)
The experiment was performed as previously described
(16). Total RNA from A549 cells
was extracted using RNAiso Plus reagent (cat. no. 9108; Takara
Biomedical Technology Co., Ltd.) and the cDNA synthesis kit (cat.
no. CW2020M; CoWin Biosciences) was used to complete the reverse
transcription according to the manufacturer's instructions. PCR was
performed using 2X Es Taq MasterMix (cat. no. CW0690H; CoWin
Biosciences). The qPCR thermocycling conditions were as follows:
95°C for 1.5 min, 94°C for 0.5 min, 56°C for 0.5 min and 72°C for 1
min for 30 cycles, with a final 10-min extension step at 72°C.
Finally, 2% agarose gel electrophoresis stained with Super Gelred
(cat. no. S2001; Suzhou Yuheng Biotechnology Co., Ltd.; www.uebio.com.) was used to separate PCR products. The
PCR bands were visualized using ImageMaster VDS. ImageJ software
(version 1.53e; National Institutes of Health) was used to
digitalize the bands. Primer sequences are included in Table I.
 | Table I.Sequences of primers used for reverse
transcription-quantitative PCR. |
Table I.
Sequences of primers used for reverse
transcription-quantitative PCR.
| Gene name |
| Fragment size | Tm (°C) |
|---|
| GAPDH | F:
GATTGTTGCCATCAACGACC | 372 | 56 |
|
| R:
GTGCAGGATGCATTGCTGAC |
|
|
| ActRIA | F:
TGGTGTAACAGGAACATCACGG | 118 | 56 |
|
| R:
GACATACTGCGAACACTACAGAG |
|
|
| ActRIB | F:
ATCACAACCGCCAGAGACTG | 136 | 56 |
|
| R:
CGCTGGACAAAGAGGGGTAA |
|
|
| ActRIIA | F:
ATTGGCCAGCATCCATCTCTTGA | 296 | 56 |
|
| R:
CGCAACCATCATAGACTAGATTC |
|
|
| ActRIIB | F:
CCTCCTCTGGGGATCGCTGT | 487 | 56 |
|
| R:
GTCCACATGACCGTAGGGGG |
|
|
| Smad3 | F:
CTCCTACTACGAGCTGAACCA | 572 | 56 |
|
| R:
AAGACACACTGGAACAGCGGA |
|
|
Western blotting
The experiment was performed as previously described
(16). Briefly, A549 cells were
lysed using protein extraction reagent (cat. no. 78503; Thermo
Fisher Scientific, Inc.) containing protease & phosphatase
inhibitor cocktail (cat. no. 87785; Thermo Fisher Scientific, Inc.)
and 5 mmol/l EDTA solution (cat. no. R1021; Thermo Scientific,
Inc.). According to the instructions, the BCA protein assay kit
(cat. no. 23227; Thermo Fisher Scientific, Inc.) was applied to
quantify the proteins, which were obtained from centrifugation of
12,000 g at 4°C for 20 min. Then, 10% SDS-PAGE gel electrophoresis
was performed to separate 30 µg of protein. After electrophoresis,
the product was transferred onto polyvinylidene difluoride membrane
(cat. no. IPVH00010; Sigma-Aldrich; Merck KGaA). Next, the
membranes were blocked with 5% skim milk in TBS-Tween 20 for 2 h at
room temperature (RT), and then incubated with specific primary
antibodies overnight at 4°C. The corresponding horseradish
peroxidase-conjugated secondary antibodies (1:160,000; Goat
anti-mouse IgG for CHOP antibody, cat. no. A3682; Goat anti-rabbit
IgG for other antibodies, cat. no. A0545; all from Merck KGaA) were
further incubated for 2 h at RT. Finally, the bands attached to the
membrane were visualized using an ECL luminescence reagent (cat.
no. BL520; Biosharp Life Sciences) using a Tanon 4600
chemiluminescent imaging system. The immunoblots were quantified
using ImageJ software, and the levels of proteins were normalized
against GAPDH. The following antibodies were used for western
blotting: ActRIIA (1:1,000; cat. no. abs149480), GAPDH (1:10,000;
cat. no. abs132004; both from Absin Bioscience, Inc.), GADD34
(1:1,000; cat. no. ab131402; Abcam), CHOP (1:1,000; cat. no. 2895),
cleaved-caspase3 (1:1,000; cat. no. 9661), ERK1/2 (1:1,000; cat.
no. 4695), phosphorylated (p)-ERK1/2 (1:2,000; cat. no. 4370),
p-Smad3 (1:1,000; cat. no. 9520), Smad3 (1:1,000; cat. no. 9523;
all from Cell Signaling Technology, Inc.), caspase-12 (1:1,000;
cat. no. WL03268), JNK (1:1,000; cat. no. WL01295) and p-JNK
(1:1,000; cat. no. WL01813; all from Wanleibio Co., Ltd.).
Kaplan-Meier (K-M) survival curves
analysis
The data of The Cancer Genome Atlas (TCGA)-LUAD were
obtained from Genomic Data Commons Data Portal (https://portal.gdc.cancer.gov/projects/TCGA-LUAD).
GBM R package (https://www.rdocumentation.org/packages/gbm/versions/2.1.8.1)
was used to analyze the data. Cox proportional hazard regression
model was used to analyze and determine independent risk factors,
and estimate the hazard ratio (HR) and 95% confidence interval.
Microarray-based data analysis
RNA-sequencing expression profiles and corresponding
clinical information for TCGA-LUAD were downloaded from the TCGA
dataset. GSE116959 profiles were obtained from the Gene Expression
Omnibus (GEO) database (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE116959).
The GSE116959 dataset was generated using GPL17077 Agilent-039494
SurePrint G3 Human GE v2 8×60K Microarray 039381 platform and
included 57 LUAD samples and 11 peritumoral normal lung tissues.
The raw data were downloaded as MINiML files and preprocessed via
background adjustment, normalization. Limma package (versions 3.18;
Bioconductor–limma) was used to analyze the data.
Statistical analysis
All data are presented as the mean ± standard
deviation. Statistical analysis was conducted using SPSS software
(version 26; IBM Corp.). Comparisons were performed using unpaired
Student's t-test or one-way analysis of variance (ANOVA), followed
by the least significant difference (LSD) test. P<0.05 was
considered to indicate a statistically significant difference.
Results
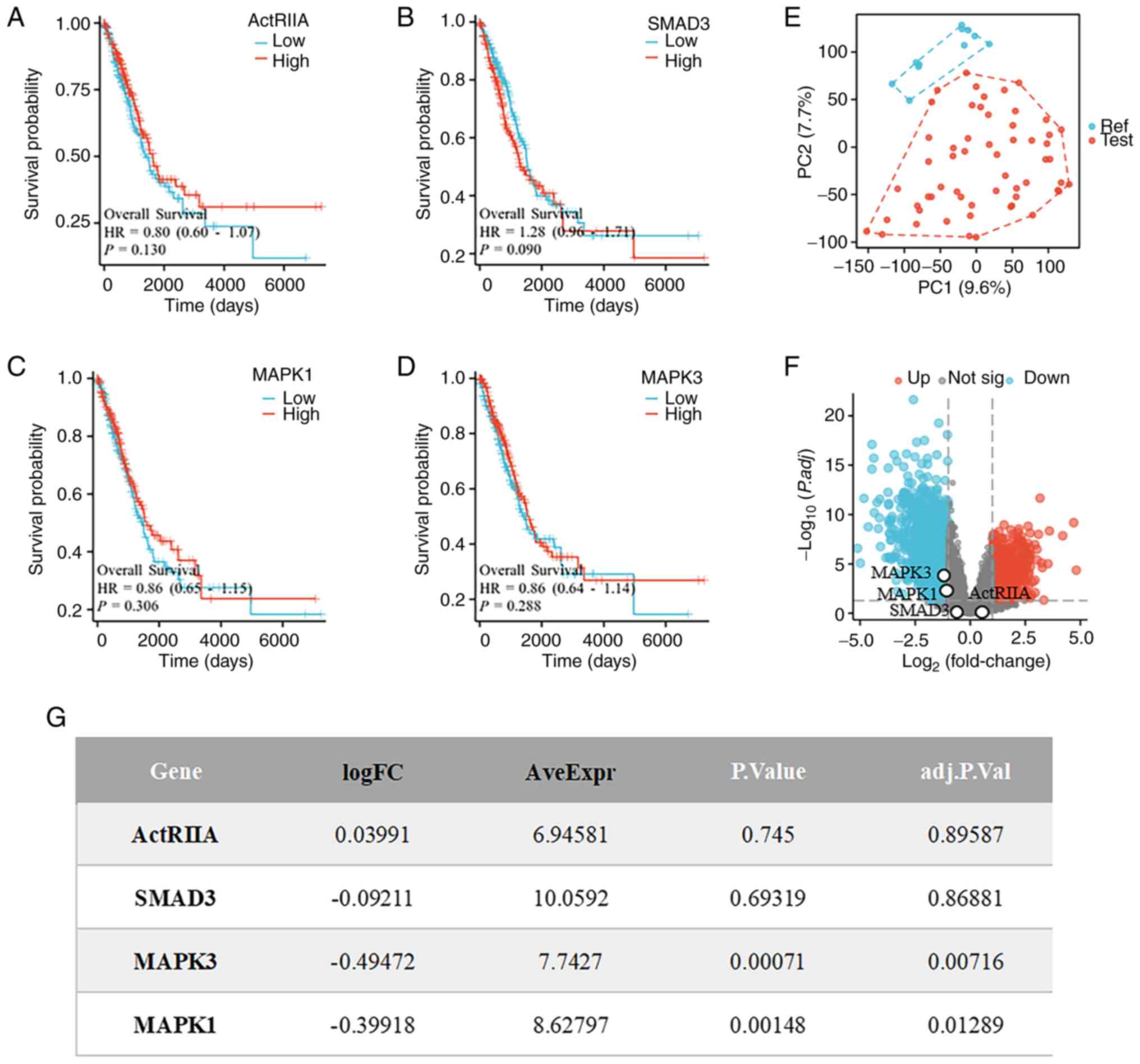
ActRIIA-MAPK signaling affects the
survival of LUAD
To confirm the clinical value of activin signaling
activation in LUAD, LUAD data were first obtained from TCGA
database and the K-M survival curves of ActRIIA and its downstream
signaling proteins in LUAD were analyzed. Although the association
did not reach the level of statistical significance, it was found
that high expression of ActRIIA was positively associated with the
survival of patients with LUAD, which was consistent with the
positive effects of mitogen-activated protein kinase 1 (MAPK1;
ERK2) and MAPK3 (ERK1) on their survival. By contrast, high
expression of Smad3 (HR=1.28 >1) was negatively associated with
their survival (Fig. 1A-D).
Moreover, when the gene expression profiles of 11 non-tumor samples
and 57 LUAD tissue samples from the GEO Series GSE116959 database
for the identification of differentially expressed genes were
examined, we found significant differences between the LUAD and
non-tumor samples (Fig. 1E). In
addition, the results of microarray-based data analysis revealed
that MAPK1 and MAPK3 were negatively associated with LUAD (Fig. 1F and G), suggesting that MAPK1 and
MAPK3 might be signaling molecules positively associated with the
survival of patients with lung cancer. These results indicated that
ActRIIA-ERK1/2 signaling may have inhibitory effects on LUAD
cells.
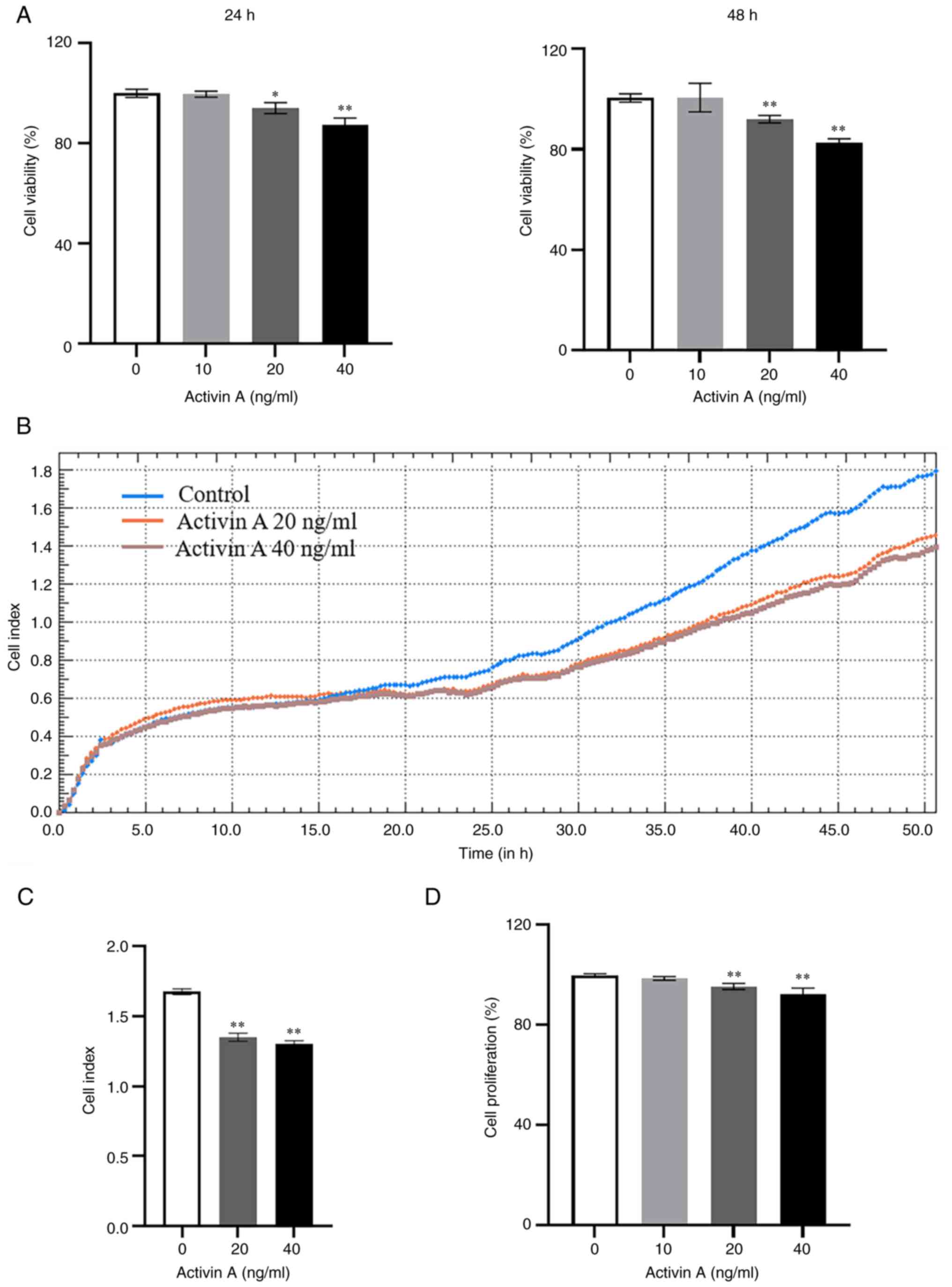
Activin A inhibits A549 cell viability
and proliferation
The results of CCK-8 assay indicated that the
viability of A549 cells decreased significantly after stimulation
with 20 and 40 ng/ml of activin A for 24 and 48 h, respectively
(Fig. 2A). RTCA was then performed,
a non-invasive and effective tool for real-time dynamic monitoring
of cell growth that is widely used for measuring cell proliferation
and adhesion (15,17), to assess the proliferation of A549
cells. The results also demonstrated that activin A significantly
inhibited the proliferation of A549 cells (Fig. 2B and C), which was consistent with
the observed decrease in cell viability. The proliferation of A549
cells was further evaluated by BrdU incorporation. The results
revealed that the proliferation of A549 cells decreased
significantly after treatment with activin A, compared with that in
the control group (Fig. 2D). These
results indicated that activin A can suppress A549 cell
proliferation.
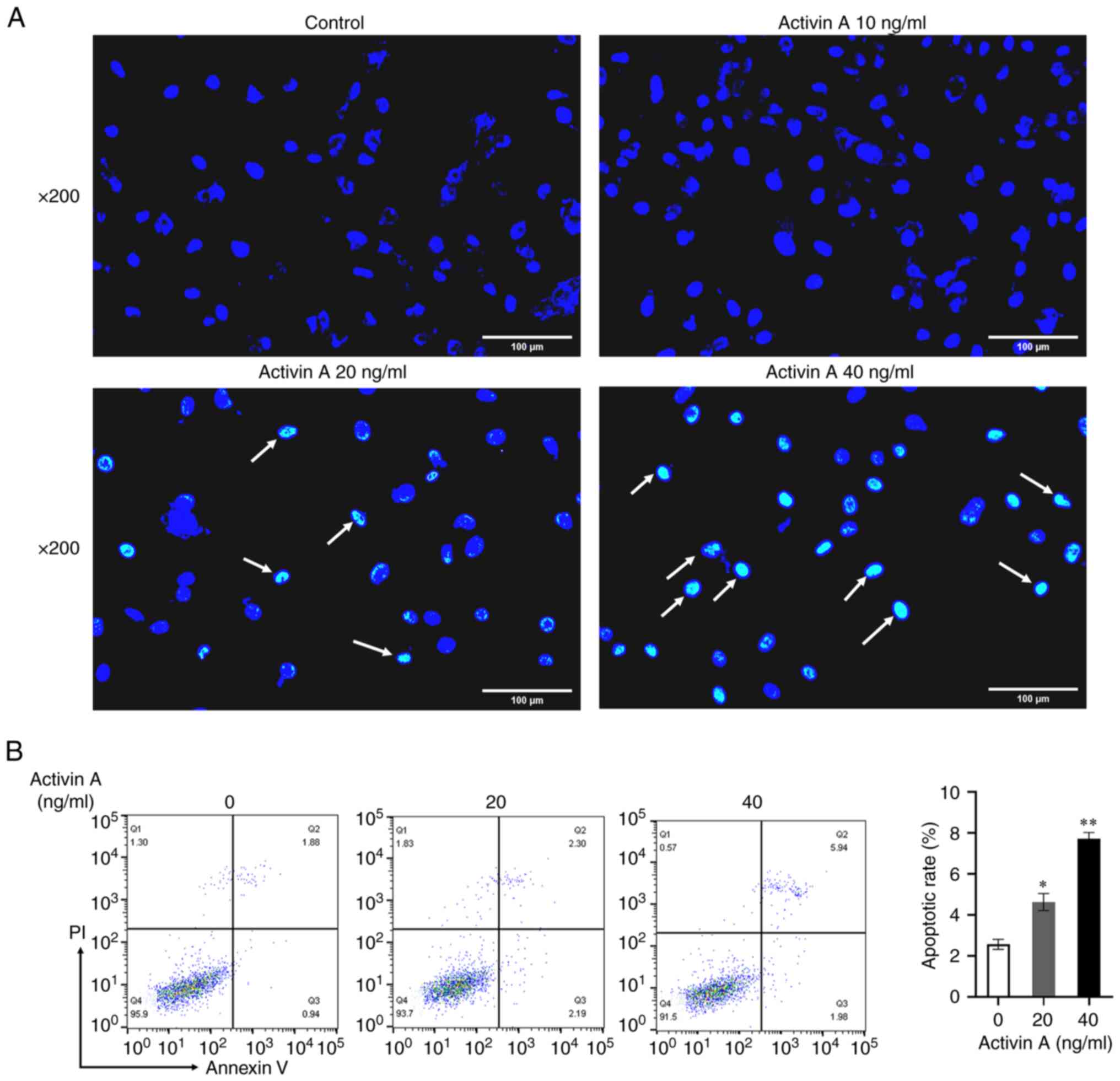
Activin A induces apoptosis of A549
cells
To ascertain whether activin A inhibits A549 cell
proliferation through apoptosis, Hoechst 33342 staining was
performed to observe alterations in cellular morphology. The
results showed that the number of apoptotic cells in the activin
A-stimulated group increased obviously compared with the control
group (Fig. 3A). Flow cytometry was
then used to detect apoptotic ratio of A549 cells. The results
revealed that, compared with the control medium, the apoptotic rate
of A549 cells stimulated with 20 ng/ml and 40 ng/ml of activin A
increased significantly (Fig. 3B),
suggesting that activin A can induce A549 cell apoptosis.
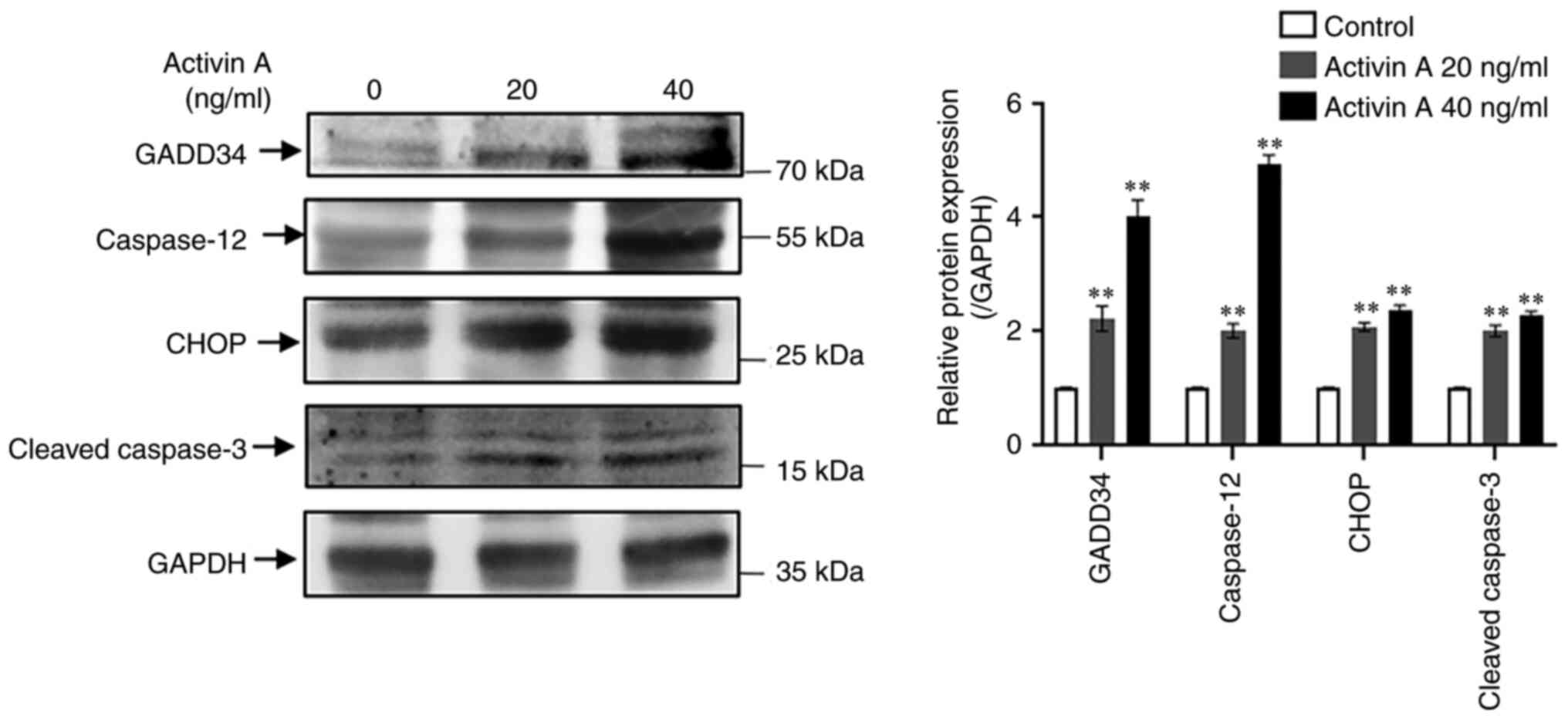
Activin A promotes the expression of
proteins related to the ER stress pathway in A549 cells
To explore whether activin A mediates apoptosis of
A549 cells via the ER stress pathway, western blotting was
performed to detect the expression of proteins associated with the
ER stress pathway in A549 cells. The results revealed that the
levels of GADD34, CHOP, caspase-12 and cleaved-caspase-3 proteins
in A549 cells were significantly upregulated by activin A (Fig. 4), indicating that activin A may
induce apoptosis of A549 cells through the ER stress pathway.
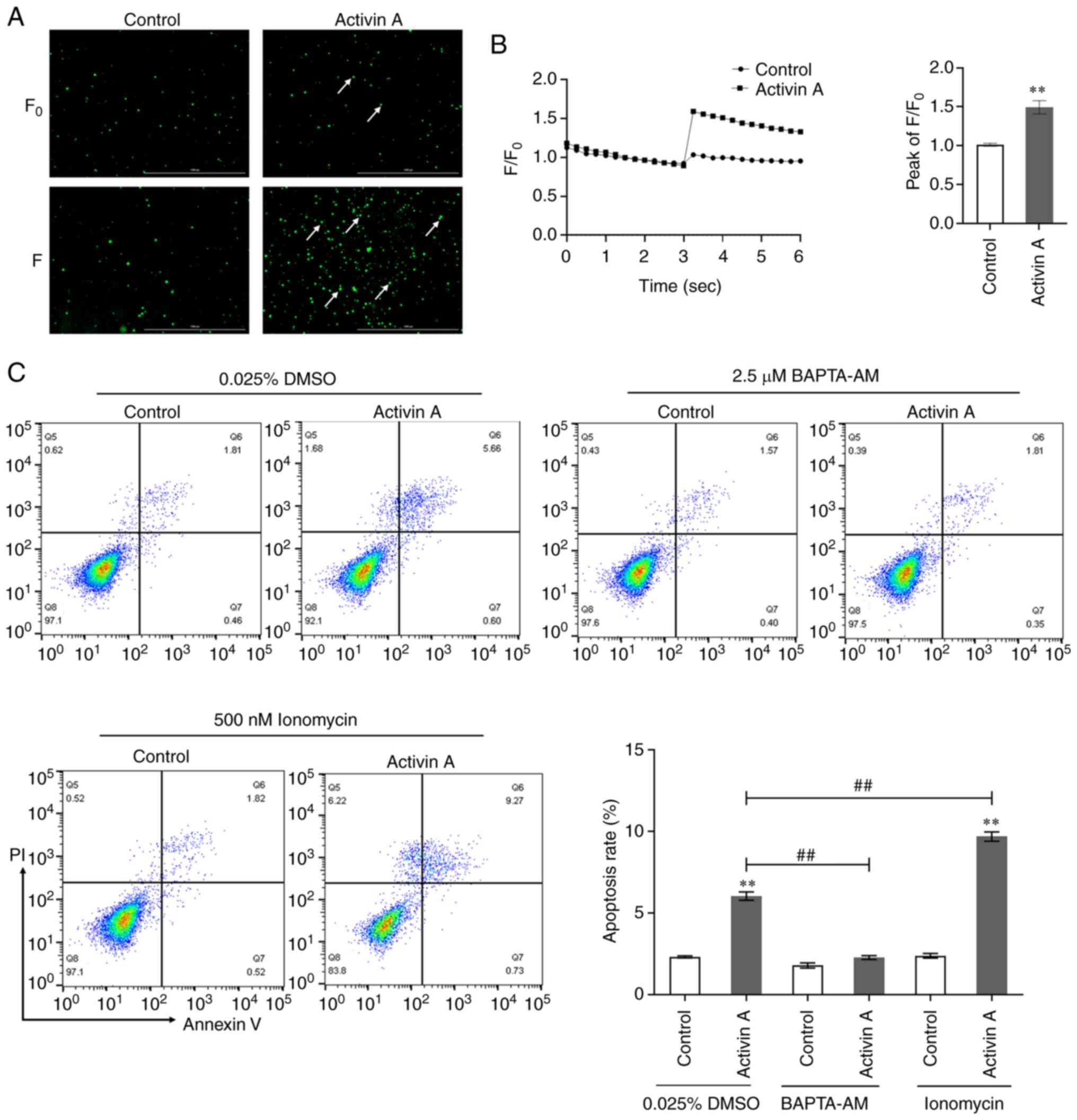
Activin A enhances calcium influx in
A549 cells
Calcium signaling is associated with various
biological behaviors such as cell proliferation, differentiation
and migration (18,19), and exerts a crucial impact on both
ER stress-induced apoptosis and mitochondria-mediated apoptosis
(20). When the calcium
fluorescence probe Fluo-4 AM was used to measure intracellular
calcium levels in A549 cells, it was identified that the
fluorescence signal intensity of calcium in A549 cells was
significantly increased by activin A treatment (Fig. 5A and B). To clarify whether calcium
signaling has a role in the activin A-induced cell apoptosis
process, A549 cells were treated with intracellular calcium ion
chelator BAPTA-AM and calcium agonist ionomycin, before assessing
cell apoptosis by flow cytometry. The present results revealed that
BAPTA-AM reversed activin A-mediated apoptosis of A549 cells,
whereas ionomycin increased the rate of apoptosis induced by
activin A (Fig. 5C). These findings
indicated that activin A induces apoptosis of A549 cells by
promoting cellular calcium influx.
Activin A increases p-ERK1/2 and
ActRIIA protein levels in A549 cells
Activin A can regulate cell activity through the
canonical Smad3 dependent or non-Smad3 signaling pathways.
Initially, RT-qPCR was performed to determine the levels of activin
receptor and Smad3 mRNA expression in A549 cells. Whereas ActRIA,
ActRIB, ActRIIB and Smad3 mRNA levels in A549 cells did not
significantly change with activin A treatment, ActRIIA mRNA level
significantly increased (Fig. 6A).
Similarly, activin A significantly upregulated ActRIIA protein
levels in A549 cells (Fig. 6B). The
results of western blotting indicated that activin A did neither
significantly affect the p-Smad3 and p-JNK protein levels, nor the
p-JNK/JNK or p-Smad3/Smad3 ratio in A549 cells. However, the
p-ERK1/2 protein level and p-ERK/ERK ratio were significantly
elevated by activin A (Fig. 6C).
The aforementioned findings indicated that activin A may regulate
ER stress pathway-triggered A549 cell apoptosis via the
Smad-independent ERK1/2 signaling.
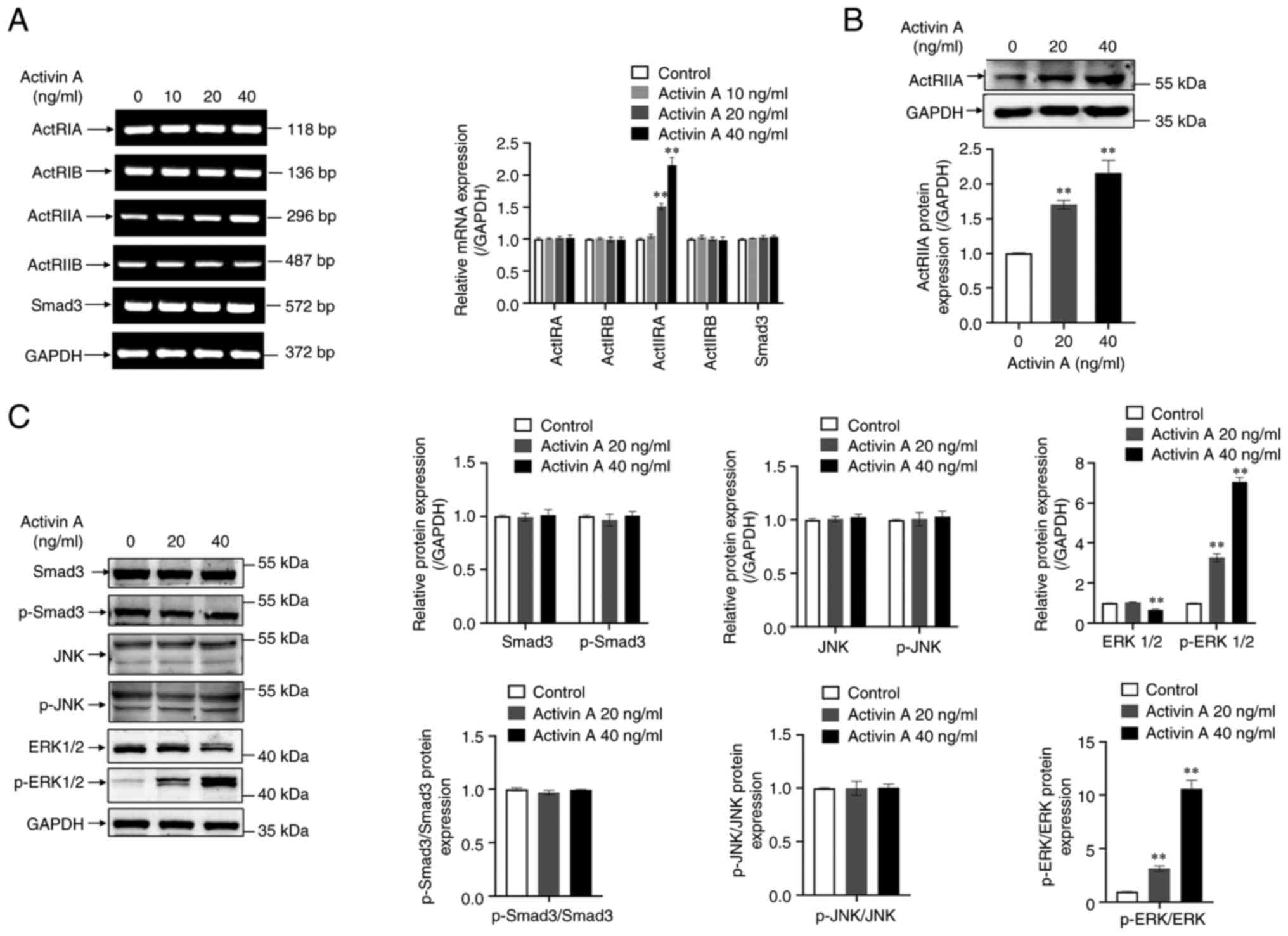 | Figure 6.Effects of activin A on expression of
activin receptors, Smad3 and MAPK signaling proteins in A549 cells.
(A) Levels of ActRIA, ActRIB, ActRIIA, ActRIIB and Smad3 mRNAs were
determined by reverse transcription-quantitative PCR in A549 cells
treated with activin A for 4 h. The graph represented the relative
levels of mRNA in three separate experiments. The levels of mRNA
were normalized against GAPDH expression, and the results were
shown as the fold-increase of the control. (B) Level of ActRIIA
protein was examined by western blotting in A549 cells treated with
activin A for 4 h. The graph represented the relative levels of
proteins in three separate experiments. The levels of ActRIIA
protein were normalized against GAPDH, and the results were
presented as the fold-increase of the control. (C) Levels of Smad3,
p-Smad3, ERK1/2, p-ERK1/2, JNK and p-JNK proteins were determined
by western blotting in A549 cells subject to 0–40 ng/ml of activin
A for 4 h. The graph represented the relative levels of protein in
three separate experiments. The levels of protein were normalized
against GAPDH expression, and the results were presented as the
fold-increase of the control. **P<0.01 compared with control
group. p-, phosphorylated. |
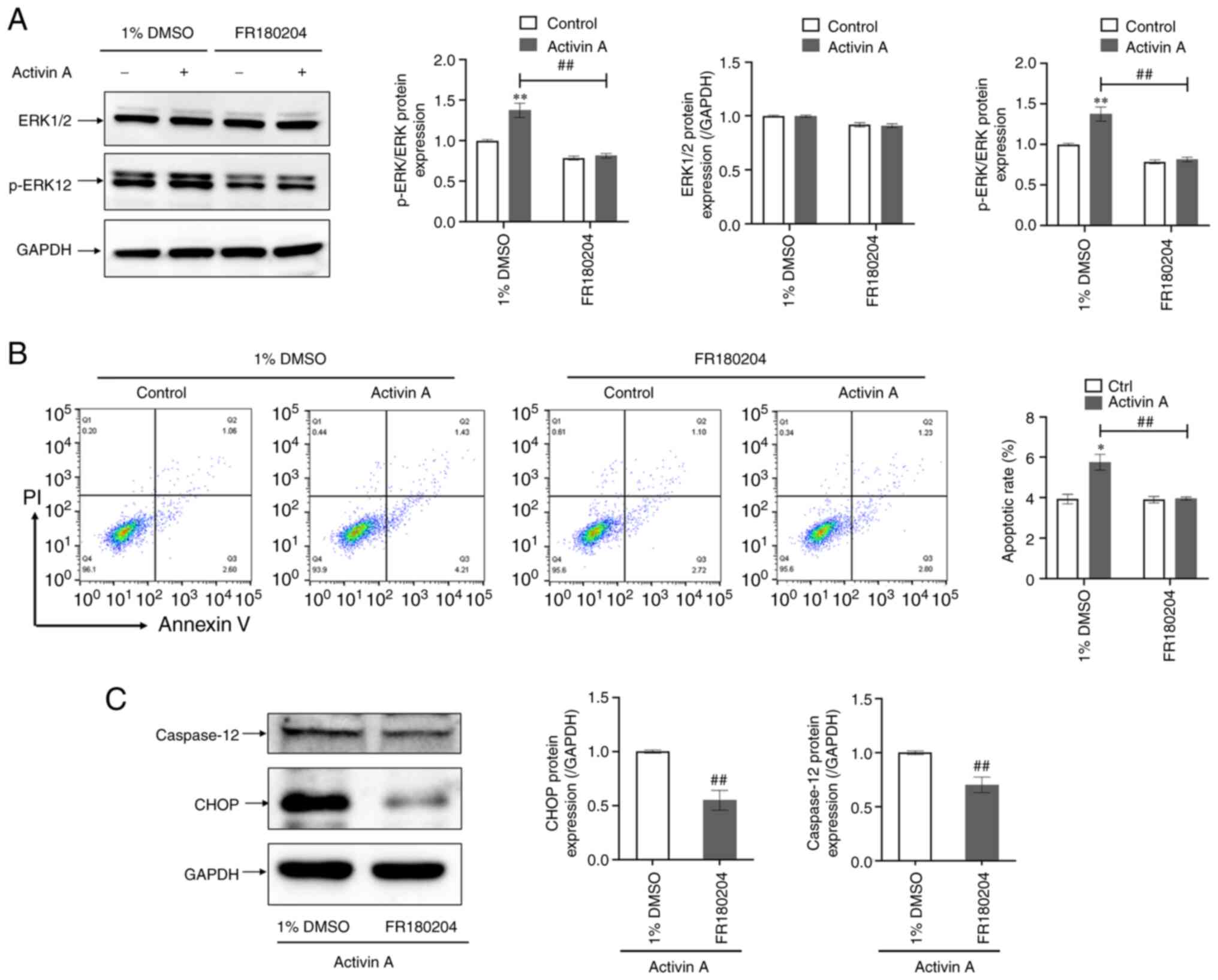
ERK inhibitor FR180204 attenuates
activin A-induced apoptosis of A549 cells
To confirm that ERK1/2 is involved in the ER stress
pathway-mediated apoptosis of A549 cells triggered by activin A,
the ERK inhibitor FR180204 was used to pretreat A549 cells for
further experiments. It was found that administration of FR180204
resulted in a significant reduction in the p-ERK protein level and
the p-ERK/ERK ratio in activin A-treated cells (Fig. 7A) as well as in the apoptotic rate
of activin A-induced A549 cells (Fig.
7B). In the next experiment, it was identified that FR180204
significantly downregulated caspase-12 and CHOP protein levels in
activin A-treated A549 cells (Fig.
7C). These findings indicated that the ERK signaling pathway
participates in activin A-induced A549 cell apoptosis through the
ER stress pathway.
Discussion
Unlike inhibin, which contains an α-subunit,
activin, a homologous or heterodimeric dimer glycoprotein, is
composed of two β-subunits linked by disulfide bonds (21). To date, four different molecular
forms of activins have been identified, specifically activin A
(βA/βA), activin B (βB/βB), activin AB (βA/βB) and activin E
(βE/βE). Only activin A, B and AB have been determined to have
defined biological activities (22). However, a previous study found that
activin E also has important biological effects as a liver cytokine
(23). The activin family member
activin A has garnered significant attention in academic research
owing to its ubiquitous tissue distribution and multifaceted
biological impact. Specifically, activin A has been found to play
crucial roles in embryogenesis, inflammatory responses, cellular
proliferation and differentiation, arterial pressure modulation and
neoplastic progression (24–27).
Recent studies have reported that, similar to TGF-β, activin A
plays a dual role in inhibiting and promoting cancer. For example,
activin A can inhibit the proliferation of mouse liver cancer cells
and human gastric cancer cells in vitro (28,29),
but can enhance the proliferation of human ovarian cancer cells and
human osteosarcoma cells (30,31).
It is evident that activin A elicits diverse
biological responses in distinct tumor cells, having both positive
and negative effects on cell proliferation. Therefore, the role of
activin A cannot be generalized. Overexpression of inhibin βA
(INHβA) has been confirmed in the tumor tissues of patients with
LUAD (7). Moreover, subsequent
studies have shown that lung cancer highly expresses the activin
binding protein-follistatin (FST), which is positively correlated
with the occurrence of LUAD and inhibits activin A-induced
apoptosis of A549 cells (32,33).
Therefore, the production of activin A by the body may serve as a
protective mechanism to counteract the effects of high levels of
FST. These studies provided evidence that activin A may have a
significant influence on the pathogenesis and development of LUAD;
targeting this protein holds promise as a therapeutic strategy for
lung neoplasms. Apoptosis is an evolutionarily conserved form of
programmed cell death that plays a crucial role in maintaining
animal growth and development and tissue homeostasis (34,35).
Apoptotic cells exhibit characteristic cytoplasmic contraction,
plasma membrane budding, plasma membrane translocation of
phosphatidylserine, DNA fragmentation and chromatin condensation
(35–37). It was found that activin A reduced
the proliferation of LUAD A549 cells, as evidenced by the current
morphological observations of an increase in apoptotic body
production. Additionally, the present flow cytometric results
indicated a higher rate of apoptosis in A549 cells treated with
activin A, suggesting that activin A induces apoptosis of A549
cells. Activin A as a multifunctional cytokine can be detrimental
for LUAD cells by inducing LUAD cell apoptosis, but activation of
activin signaling in tumor treatment may also lead to disease
damage, such as promoting the development of renal injury and renal
fibrosis (38,39). Therefore, how to make reasonable use
of activin A to treat tumors is also a problem that needs to be
solved in the future.
The three primary pathways of cell apoptosis
comprise the death receptor pathway, mitochondrial pathway and ER
stress pathway (40,41). Studies have reported that the death
receptor pathway is triggered mainly by the binding of specific
members of the tumor necrosis factor (TNF) superfamily to members
of the cell surface TNF receptor family (TNF-R) (40,42).
The mitochondrial apoptosis of cells is mediated by the interaction
between the anti-apoptotic and pro-apoptotic components of the B
cell lymphoma 2 (Bcl-2) family (43). Downregulation of Bcl-2 and
upregulation of Bcl-2-associated X protein (Bax) and Bcl-2
antagonist killer (Bak) can result in the mitochondrial membrane
permeability transition, which consequently augments the release of
cytochrome c, leading to the activation of caspase-3 and the
initiation of cellular apoptosis (40,44,45).
The ER has various functions such as maintaining the
homeostasis of intracellular Ca2+ storage, the folding
and maturation of protein destined for the plasma membrane, the
extracellular space and other secretory compartments. Impairment of
the proper function of the ER results in ER stress, and prolonged
exposure to ER stress will lead to apoptosis (41). ER stress apoptosis can be triggered
by inositol-requiring enzyme 1 (IRE1)/ASK1/JNK signaling and
CHOP/GADD153 and caspase-12 kinase upregulation (46). Upon ER stress induction, CHOP, an
apoptotic response effector, is activated, thereby causing an
increase in GADD34, a transcription target of CHOP (42). Previous studies have revealed that
activin A can activate Bcl-2 family proteins and caspase-3 protein,
thereby inducing apoptosis in A549 and NS-1 cells through the
mitochondrial pathway (44,47). The data collected in the present
study demonstrated that activin A has the potential to induce
apoptosis in A549 cells through the ER stress pathway, as evidenced
by the significant upregulation of CHOP, GADD34, caspase-12 and
cleaved caspase-3 protein levels.
Ca2+, an important secondary messenger in
cells, plays a crucial role in cell growth, differentiation,
migration and apoptosis (18–20).
Ca2+ released from the ER is a key factor in regulation
of apoptosis (19). The imbalance
in calcium levels in ER interferes with protein folding, leading to
the accumulation of unfolded or misfolded proteins, which
eventually causes ER stress (20).
During a state of ER stress, CHOP-induced expression of ERO1α can
activate the ER Ca2+ release channel. The cytoplasmic
Ca2+ released from the ER triggers the activation of
calcium/calmodulin-dependent protein kinase II (CaMKII), ultimately
resulting in the induction of apoptotic pathways (41), indicating that Ca2+ also
has a significant effect on CHOP-mediated apoptosis. The present
findings demonstrated that activin A promotes intracellular calcium
influx in A549 cells. It was observed that utilization of the
calcium antagonist BAPTA-AM significantly inhibited activin-induced
apoptosis in A549 cells, whereas the calcium agonist ionomycin
exacerbated activin A-induced cell apoptosis. As the present
findings indicated that calcium signaling has a pivotal role in
activin A-induced A549 cell apoptosis, they suggest that the
rational use of calcium agonists to increase the intracellular
calcium flux has the potential to induce apoptosis in LUAD
cells.
It has been reported that the signaling pathways of
activin A mainly include Smad3-mediated canonical pathways and
non-Smad-mediated alternative signaling pathways (48). In the canonical pathway mediated by
Smad3, activin A activates Smad2/3 by binding to ActRII and
recruiting ActRI, thereby activating downstream molecules to exert
biological effects. Alternative signaling pathways mainly include
the ERK1/2-MAPK, p38-MAPK, JNK and phosphatidylinositol-3-kinase
(PI3K)/protein kinase B (AKT) pathways (17,48,49).
ERK1/2 are downstream components of the phosphorylation cascade of
MAPK, which is closely related to biological functions including
cell proliferation, differentiation, migration and invasion
(50,51). Although ERK1/2 protein often acts as
a survival promoting factor in the MAPK family (52,53),
an increasing number of studies have demonstrated that activation
of the ERK1/2 protein can induce cell apoptosis (54–56).
The results of the present study indicated that
activin A upregulated ActRIIA receptor expression in A549 cells,
but had no discernible impact on ActRIIB receptor expression. It
was found that activin A preferentially activated the ERK1/2
signaling over Smad3 and JNK signaling. Notably, it was observed
that administration of the ERK inhibitor FR180204 effectively
inhibited the apoptosis of A549 cells triggered by activin A and
concomitantly reduced the level of CHOP protein stimulated by
activin A. Analysis of K-M survival curves and microarray-based
data analysis also indicated that ActRIIA and ERK1/2 activation was
positively associated with the survival of patients with LUAD.
Collectively, the present findings corroborated that activin
A-induced A549 cell apoptosis can be mediated by the ER stress
pathway via ActRIIA-ERK signaling.
In summary, the findings of the present study
demonstrated that activin A induces ER stress pathway apoptosis in
LUAD A549 cells by activating ActRIIA-ERK1/2 signaling, which is
also related to calcium signaling. Therefore, they provide evidence
that agonists of ERK signaling and calcium signaling have potential
for inducing apoptosis of LUAD cells in clinical therapy for
treatment of patients with LUAD.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Science Foundation of
Jilin (grant no. 20220402079GH) and the Science Foundation of Jilin
Provincial Health Commission (grant nos. 2019J013 and
2021JC030).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author upon reasonable
request. The datasets generated and/or analyzed during the current
study are available in the TCGA (https://portal.gdc.cancer.gov/projects/TCGA-LUAD)
and GEO (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE116959)
repositories.
Authors' contributions
ZL and XC designed the experiments. FZ and YQ
executed the experiments. FZ, JL and BL analyzed the data. FZ
drafted the original manuscript. JL, ZL and XC revised and edited
the manuscript. FZ, ZL and XC confirm the authenticity of all the
raw data. ZL and XC provided funding projects. All authors read and
approved the final version of the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD, Wagle NS and Jemal
A: Cancer statistics, 2023. CA Cancer J Clin. 73:17–48. 2023.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Herbst RS, Morgensztern D and Boshoff C:
The biology and management of non-small cell lung cancer. Nature.
553:446–454. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yu Z, Tang H, Chen S, Xie Y, Shi L, Xia S,
Jiang M, Li J and Chen D: Exosomal LOC85009 inhibits docetaxel
resistance in lung adenocarcinoma through regulating ATG5-induced
autophagy. Drug Resist Updat. 67:1009152023. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Miller KD, Nogueira L, Devasia T, Mariotto
AB, Yabroff KR, Jemal A, Kramer J and Siegel RL: Cancer treatment
and survivorship statistics, 2022. CA Cancer J Clin. 72:409–436.
2022. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhao C, Zhai Y, Geng R, Wu K, Song W, Ai N
and Ge W: Genetic analysis of activin/inhibin beta subunits in
zebrafish development and reproduction. PLoS Genet.
18:e10105232022. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ge J, Sun H, Li J, Shan Y, Zhao Y, Liao F,
Yang Y, Cui X and Liu Z: Involvement of CHOP in activin A-induced
myeloma NS-1 cell apoptosis. Oncol Rep. 42:2644–2654.
2019.PubMed/NCBI
|
|
7
|
Seder CW, Hartojo W, Lin L, Silvers AL,
Wang Z, Thomas DG, Giordano TJ, Chen G, Chang AC, Orringer MB and
Beer DG: Upregulated INHBA expression may promote cell
proliferation and is associated with poor survival in lung
adenocarcinoma. Neoplasia. 11:388–396. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yu SY, Luan Y, Tang S, Abazarikia A, Dong
R, Caffrey TC, Hollingsworth MA, Oupicky D and Kim SY: Uncovering
tumor-promoting roles of activin a in pancreatic ductal
adenocarcinoma. Adv Sci (Weinh). 10:e22070102023. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kalli M, Mpekris F, Wong CK, Panagi M,
Ozturk S, Thiagalingam S, Stylianopoulos T and Papageorgis P:
Activin a signaling regulates IL13Rα2 expression to promote breast
cancer metastasis. Front Oncol. 9:322019. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ervolino De Oliveira C, Dourado MR,
Sawazaki-Calone Í, Costa De Medeiros M, Rossa Júnior C, De Karla
Cervigne N, Esquiche León J, Lambert D, Salo T, Graner E and
Coletta RD: Activin A triggers angiogenesis via regulation of VEGFA
and its overexpression is associated with poor prognosis of oral
squamous cell carcinoma. Int J Oncol. 57:364–376. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Oakes SA and Papa FR: The role of
endoplasmic reticulum stress in human pathology. Annu Rev Pathol.
10:173–194. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Di Conza G and Ho PC: ER stress responses:
An emerging modulator for innate immunity. Cells. 9:6952020.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Fu X, Cui J, Meng X, Jiang P, Zheng Q,
Zhao W and Chen X: Endoplasmic reticulum stress, cell death and
tumor: Association between endoplasmic reticulum stress and the
apoptosis pathway in tumors (Review). Oncol Rep. 45:801–808. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Xue LX, Liu HY, Cui Y, Dong Y, Wang JQ, Ji
QY, He JT, Yao M, Wang YY, Shao YK, et al: Neuroprotective effects
of Activin A on endoplasmic reticulum stress-mediated apoptotic and
autophagic PC12 cell death. Neural Regen Res. 12:779–786. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Jiang L, Qi Y, Kong X, Wang R, Qi J, Lin
F, Cui X and Liu Z: Activin A as a novel chemokine induces
migration of L929 fibroblasts by ERK signaling in microfluidic
devices. Front Cell Dev Biol. 9:6603162021. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Jiang L, Liu B, Qi Y, Zhu L, Cui X and Liu
Z: Antagonistic effects of activin A and TNF-alpha on the
activation of L929 fibroblast cells via Smad3-independent
signaling. Sci Rep. 10:206232020. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Li J, Qi Y, Yang K, Zhu L, Cui X and Liu
Z: Follistatin is a novel chemoattractant for migration and
invasion of placental trophoblasts of mice. Cells. 11:38162022.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Norgard RJ, Pitarresi JR, Maddipati R,
Aiello-Couzo NM, Balli D, Li J, Yamazoe T, Wengyn MD, Millstein ID,
Folkert IW, et al: Calcium signaling induces a partial EMT. EMBO
Rep. 22:e518722021. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Patergnani S, Danese A, Bouhamida E,
Aguiari G, Previati M, Pinton P and Giorgi C: Various aspects of
calcium signaling in the regulation of apoptosis, autophagy, cell
proliferation, and cancer. Int J Mol Sci. 21:83232020. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Moon DO: Calcium's role in orchestrating
cancer apoptosis: Mitochondrial-centric perspective. Int J Mol Sci.
24:89822023. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Valero-Aracama MJ, Zheng F and Alzheimer
C: Dorsal-Ventral gradient of activin regulates strength of
GABAergic inhibition along longitudinal axis of mouse hippocampus
in an Activity-Dependent fashion. Int J Mol Sci. 24:131452023.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Bloise E, Ciarmela P, Dela Cruz C, Luisi
S, Petraglia F and Reis FM: Activin a in mammalian physiology.
Physiol Rev. 99:739–780. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hashimoto O, Funaba M, Sekiyama K, Doi S,
Shindo D, Satoh R, Itoi H, Oiwa H, Morita M, Suzuki C, et al:
Activin e controls energy homeostasis in both brown and white
adipose tissues as a hepatokine. Cell Rep. 25:1193–1203. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liu G, Qi Y, Wu J, Lin F, Liu Z and Cui X:
Follistatin is a crucial chemoattractant for mouse decidualized
endometrial stromal cell migration by JNK signalling. J Cell Mol
Med. 27:127–140. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Locci M, Wu JE, Arumemi F, Mikulski Z,
Dahlberg C, Miller AT and Crotty S: Activin A programs the
differentiation of human TFH cells. Nat Immunol. 17:976–984. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ge J, Fan Y, Lu Y, Qi Y, Wang M and Liu Z:
Activin A increases arterial pressure in the hypothalamic
paraventricular nucleus in rats by angiotension II. Neuroreport.
27:683–688. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Dzierlega K, Chakraborty M, Lee M, Soliman
AM, Parker D, Khan S, Chan YT, Akbari M, Yokota T, Winer S, et al:
Activin A-Expressing polymorphonuclear Myeloid-Derived suppressor
cells infiltrate skeletal and cardiac muscle and promote cancer
cachexia. J Immunol. 211:497–507. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Staudacher JJ, Arnold A, Kühl AA, Pötzsch
M, Daum S, Winterfeld M, Berg E, Hummel M, Rau B, Stein U and
Treese C: Prognostic impact of activin subunit inhibin beta A in
gastric and esophageal adenocarcinomas. BMC Cancer. 22:9532022.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hamang M, Yaden B and Dai G:
Gastrointestinal pharmacology activins in liver health and disease.
Biochem Pharmacol. 214:1156682023. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Meier D, Lodberg A, Gvozdenovic A,
Pellegrini G, Neklyudova O, Born W, Fuchs B, Eijken M and M Botter
S: Inhibition of the activin receptor signaling pathway: A novel
intervention against osteosarcoma. Cancer Med. 10:286–296. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Tao JJ, Cangemi NA, Makker V, Cadoo KA,
Liu JF, Rasco DW, Navarro WH, Haqq CM and Hyman DM: First-in-human
phase I study of the activin A inhibitor, STM 434, in patients with
granulosa cell ovarian cancer and other advanced solid tumors. Clin
Cancer Res. 25:5458–5465. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Cheng X and Ferrell JE Jr: Apoptosis
propagates through the cytoplasm as trigger waves. Science.
361:607–612. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhang P, Ruan Y, Xiao J, Chen F and Zhang
X: Association of serum follistatin levels with histological types
and progression of tumor in human lung cancer. Cancer Cell Int.
18:1622018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chen F, Ren P, Feng Y, Liu H, Sun Y, Liu
Z, Ge J and Cui X: Follistatin is a novel biomarker for lung
adenocarcinoma in humans. PLoS One. 9:e1113982014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Pistritto G, Trisciuoglio D, Ceci C,
Garufi A and D'Orazi G: Apoptosis as anticancer mechanism: Function
and dysfunction of its modulators and targeted therapeutic
strategies. Aging (Albany NY). 8:603–619. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Liu S, Luo L, Zuo F, Geng Y, Ou Y, Chen D,
Yang S, Luo W, Wang Y, Wang J and Huang X: Immunosuppression and
apoptosis activation mediated by p53-Bcl2/Bax signaling pathway-The
potential mechanism of goldfish (Carassius auratus Linnaeus) gill
disease caused by Myxobolus ampullicapsulatus. Front Immunol.
13:9989752022. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Geng Y, Liu P, Xie Y, Liu Y, Zhang X, Hou
X and Zhang L: Xanthatin suppresses pancreatic cancer cell growth
via the ROS/RBL1 signaling pathway: In vitro and in vivo insights.
Phytomedicine. 119:1550042023. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Marini KD, Croucher DR, McCloy RA,
Vaghjiani V, Gonzalez-Rajal A, Hastings JF, Chin V, Szczepny A,
Kostyrko K, Marquez C, et al: Inhibition of activin signaling in
lung adenocarcinoma increases the therapeutic index of platinum
chemotherapy. Sci Transl Med. 10:eaat35042018. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Yuan C, Ni L and Wu X: Activin A
activation drives renal fibrosis through the STAT3 signaling
pathway. Int J Biochem Cell Biol. 134:1059502021. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Hotchkiss RS, Strasser A, McDunn JE and
Swanson PE: Cell death. N Engl J Med. 361:1570–1583. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Kim C and Kim B: Anti-Cancer natural
products and their bioactive compounds inducing ER Stress-Mediated
apoptosis: A review. Nutrients. 10:10212018. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Roberts JZ, Crawford N and Longley DB: The
role of ubiquitination in apoptosis and necroptosis. Cell Death
Differ. 29:272–284. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Wong HY, Hui Q, Hao Z, Warnock GL, Woo M,
Luciani DS and Marzban L: The role of mitochondrial apoptotic
pathway in islet amyloid-induced β-cell death. Mol Cell Endocrinol.
537:1114242021. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Zhang Y, Qi Y, Zhao Y, Sun H, Ge J and Liu
Z: Activin A induces apoptosis of mouse myeloma cells via the
mitochondrial pathway. Oncol Lett. 15:2590–2594. 2018.PubMed/NCBI
|
|
45
|
Green DR: The mitochondrial pathway of
apoptosis Part II: The BCL-2 protein family. Cold Spring Harb
Perspect Biol. 14:a0410462022. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Hu H, Tian M, Ding C and Yu S: The C/EBP
Homologous protein (CHOP) transcription factor functions in
endoplasmic reticulum stress-induced apoptosis and microbial
infection. Front Immunol. 9:30832018. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Wang B, Feng Y, Song X, Liu Q, Ning Y, Ou
X, Yang J, Zhang X and Wen F: Involvement of ERK, Bcl-2 family and
caspase 3 in recombinant human activin A-induced apoptosis in A549.
Toxicology. 258:176–183. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Wiley MB, Bauer J, Mehrotra K,
Zessner-Spitzenberg J, Kolics Z, Cheng W, Castellanos K, Nash MG,
Gui X, Kone L, et al: Non-Canonical activin A signaling stimulates
Context-dependent and Cellular-Specific outcomes in CRC to promote
tumor cell migration and immune tolerance. Cancers (Basel).
15:30032023. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Sugatani T: Systemic activation of activin
a signaling causes chronic kidney Disease-Mineral bone disorder.
Int J Mol Sci. 19:24902018. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Zhang X, Liu T, Huang J and He J: PICALM
exerts a role in promoting CRC progression through ERK/MAPK
signaling pathway. Cancer Cell Int. 22:1782022. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Lucas RM, Luo L and Stow JL: ERK1/2 in
immune signalling. Biochem Soc Trans. 50:1341–1352. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Lu Z and Xu S: ERK1/2 MAP kinases in cell
survival and apoptosis. IUBMB Life. 58:621–631. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Zhang J, Zhang J, Liu W, Ge R, Gao T, Tian
Q, Mu X, Zhao L and Li X: UBTF facilitates melanoma progression via
modulating MEK1/2-ERK1/2 signalling pathways by promoting GIT1
transcription. Cancer Cell Int. 21:5432021. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Leung GP, Feng T, Sigoillot FD, Geyer FC,
Shirley MD, Ruddy DA, Rakiec DP, Freeman AK, Engelman JA,
Jaskelioff M and Stuart DD: Hyperactivation of MAPK signaling is
deleterious to RAS/RAF-mutant melanoma. Mol Cancer Res. 17:199–211.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Lee CY, Hsiao YH, Chen PN, Wu HH, Lu CY,
Yang SF and Wang PH: CLEFMA induces intrinsic and extrinsic
apoptotic pathways through ERK1/2 and p38 signalling in uterine
cervical cancer cells. J Cell Mol Med. 27:446–455. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Sugiura R, Satoh R and Takasaki T: ERK: A
Double-Edged sword in cancer. ERK-Dependent apoptosis as a
potential therapeutic strategy for cancer. Cells. 10:25092021.
View Article : Google Scholar : PubMed/NCBI
|