Introduction
Each mitotic cell division results in the shortening
of telomeres (1). In somatic cells,
subsequent shortening of telomeres eventually leads to senescence.
The multiunit enzyme telomerase maintains telomere length by adding
repetitive sequences to telomeres at each replicative cycle.
However, the telomerase activity is absent in somatic cells. By
contrast, malignant cells (re-)express telomerase or use an
alternative mechanism to maintain telomere length, known as
alternative lengthening of telomeres (ALT) (2).
Telomerase reverse transcriptase (TERT) is an
essential part of the telomerase enzyme complex (3). Most recently, specific point mutations
were described in the promoter region of the TERT gene in
familial and sporadic melanoma (4).
These point mutations lead to a cytosine to thymine exchange at the
positions 228 or 250 upstream of the start codon of TERT,
further referred to as C228T and C250T, respectively (3–5). Through
these mutations, the TERT promoter acquires novel binding
sites for the E twenty-six transcription factor, resulting in
increased TERT expression (6).
Subsequently, TERT promoter mutations were described in a
variety of tumors, including several neuronal tumors as
medulloblastoma (7) and glioma, as
well as other tumor entities (4,5). Of note,
the presence of TERT promoter mutations often correlated
with aggressive disease and an adverse outcome.
Neuroblastoma is the most common extracranial tumor
during childhood arising from the neural-crest (8,9). Only few
recurrent mutations or genomic rearrangements have been described
thus far (10). Among these are the
genomic amplifications of MYCN and activating mutations of
anaplastic lymphoma kinase. Most recently, two independent studies
described ALT through mutations in the transcriptional regulator
α-thalassemia/mental retardation X-linked (ATRX) in a minor
fraction of neuroblastomas (11,12).
Although the ALT phenotype may not be limited to neuroblastomas
with ATRX mutations, the majority of neuroblastomas express
telomerase and in particular TERT (13). Of note, high TERT expression was
detected in MYCN-amplified tumors, but even in MYCN
single copy neuroblastomas a high TERT expression correlated with
adverse outcome (13). By contrast,
low or absent TERT expression was detected in favorable
neuroblastomas and absence of telomerase activity has been proposed
as a potential mechanism to explain the high rate of spontaneous
regression of favorable neuroblastomas (14). While MYCN is known to induce
TERT transcription (15,16) and
TERT amplification has been described in few neuroblastoma
cases (17), the mechanism that drives
TERT expression remains elusive in the majority of
neuroblastomas.
As TERT promoter mutations are frequent in
neural-crest-derived melanoma, as well as in various neuronal
tumors including medulloblastoma, the present study analyzed the
frequency of TERT promoter mutations in neuroblastoma.
Materials and methods
Primary tumor samples
Primary tumor samples and patient data were obtained
from the German Society of Pediatric Oncology and Hematology Tumor
Bank and Neuroblastoma Study Center (Cologne, Germany),
respectively. All the patients were registered with the German
neuroblastoma study and written informed consent was obtained. A
tumor content of ≥60% was confirmed by a pathologist. DNA was
isolated from ~20 mg of snap-frozen tissue obtained prior to
cytotoxic treatment using the Puregene Blood Core kit B (Qiagen,
Hilden, Germany), according to the manufacturer's instructions.
Cell culture
Neuroblastoma cell lines BEC(2), CHP134, IMR-32, IMR-5-75, KCN, KCN(R),
Kelly, LAN1, LAN5, LAN6, NGP, NLF, SH-EP, SH-SY5Y, SK-N-AS,
SK-N-BE, SK-N-FI and SK-N-SH were cultured in RPMI-1640
supplemented with 10% fetal calf serum, 5% penicillin (100 U/ml)
and streptomycin (100 µg/ml) until reaching a confluence of 70%.
Cells were collected and DNA was isolated using the QIAamp DNA kit
(Qiagen). All the cell lines were authenticated by short tandem
repeat genotyping performed by DSMZ (Braunschweig, Germany) or
IDEXX GmbH (Ludwigsburg, Germany).
Polymerase chain reaction (PCR) and
sequencing
DNA (100 ng) was used for the PCR reaction. To each
PCR reaction, TERT forward (ACGAACGTGGCCAGCGGCAG) and
reverse primers (CTGGCGTCCCTGCACCCTGG) were added to amplify a
474-base pair (bp) long region of the TERT promoter (390-bp
upstream and 80-bp downstream of the start codon). The PCR program
used was as follows: 95°C for 180 sec, followed by 95°C for 30 sec,
62°C for 45 sec and 72°C for 60 sec repeated 35 times, and a final
amplification step at 72°C for 600 sec. Sanger sequencing was
performed by Sequence Laboratories Göttingen GmbH (Göttingen,
Germany).
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was isolated from cells using the
RNeasyMini kit (Qiagen) and cDNA synthesis was performed using the
SuperScript® RT kit (Invitrogen Life Technologies, Darmstadt,
Germany). TERT expression was monitored using the QuantiTect
Primer Assay™ (Qiagen). Expression values were normalized to the
geometric mean of GAPDH (18).
Data analysis and error propagation were performed using the
qbasePLUS software version 1.5 (http://www.biogazelle.com).
Pyrosequencing
To determine the presence of the mutant C228T allele
or the wild-type allele using pyrosequencing, the following primers
were used to amplify a 151-bp amplicon: Biotinylated forward primer
5′-BIOTIN-CTTCACCTTCCAGCTCCGC-3′, and reverse primer
5′-CGCTGCCTGAAACTCGC-3′. The PCR products were analyzed by
pyrosequencing using the primer 5′-GAGGGGCTGGGAGGGCCC-3′, on the
PyroMark Q96 MD system according to the manufacturer's instructions
(Qiagen) and as previously described (19). Results were analyzed using PyroMark MD
1.0 software (Biotage, Uppsala, Sweden).
Genome-wide single-nucleotide
polymorphism (SNP) analysis
SNP array experiments were performed according to
the standard protocol for Affymetrix CytoScanHD arrays (Affymetrix,
Inc., Santa Clara, CA, USA). In brief, a 250-ng sample of genomic
DNA was digested with NspI, ligated to adaptors, amplified by PCR,
fragmented and biotin-labeled. The labeled samples were hybridized
to Affymetrix CytoscanHD arrays, followed by washing, staining and
scanning in the Affymetrix GeneChip Scanner 3000. Analysis was
performed using the Affymetrix Chromosome Analysis Suite v2.1.
Results
TERT promoter mutations are absent in
primary neuroblastomas
To determine the frequency of TERT promoter
mutations in neuroblastoma, including the previously described
C228T and C250T mutations, the respective TERT promoter
region of 131 primary neuroblastomas was sequenced. These tumors
represented the whole spectrum of neuroblastoma regarding stage,
MYCN and 1p36 status (Table I).
Of note, none of the analyzed neuroblastomas harbored a C228T or a
C250T mutation in the TERT promoter region.
 | Table I.Patient data of the 131 sequenced
primary neuroblastomas tumors. |
Table I.
Patient data of the 131 sequenced
primary neuroblastomas tumors.
| Characteristics | Values, n |
|---|
| Patients | 131 |
| Gender |
|
|
Female | 47 |
| Male | 84 |
| Stage (INSS) |
|
| 1 | 29 |
| 2a | 12 |
| 2b | 7 |
| 3 | 19 |
| 4 | 50 |
| 4S | 14 |
| MYCN status |
|
|
Non-amplified | 96 |
|
Amplified | 33 |
| Not
detectable | 2 |
| 1p36 status |
|
|
Wild-type | 90 |
|
Deletion | 24 |
|
Imbalance | 12 |
| Not
detectable | 5 |
| Age at diagnosis,
years |
|
|
<1 | 36 |
|
1–2 | 50 |
|
>2 | 45 |
TERT promoter mutations in
neuroblastoma cell lines
In addition to primary neuroblastomas, 19
neuroblastoma cell lines were analyzed. In 3 out of 19 analyzed
cell lines (16%) a mutation of the TERT promoter was
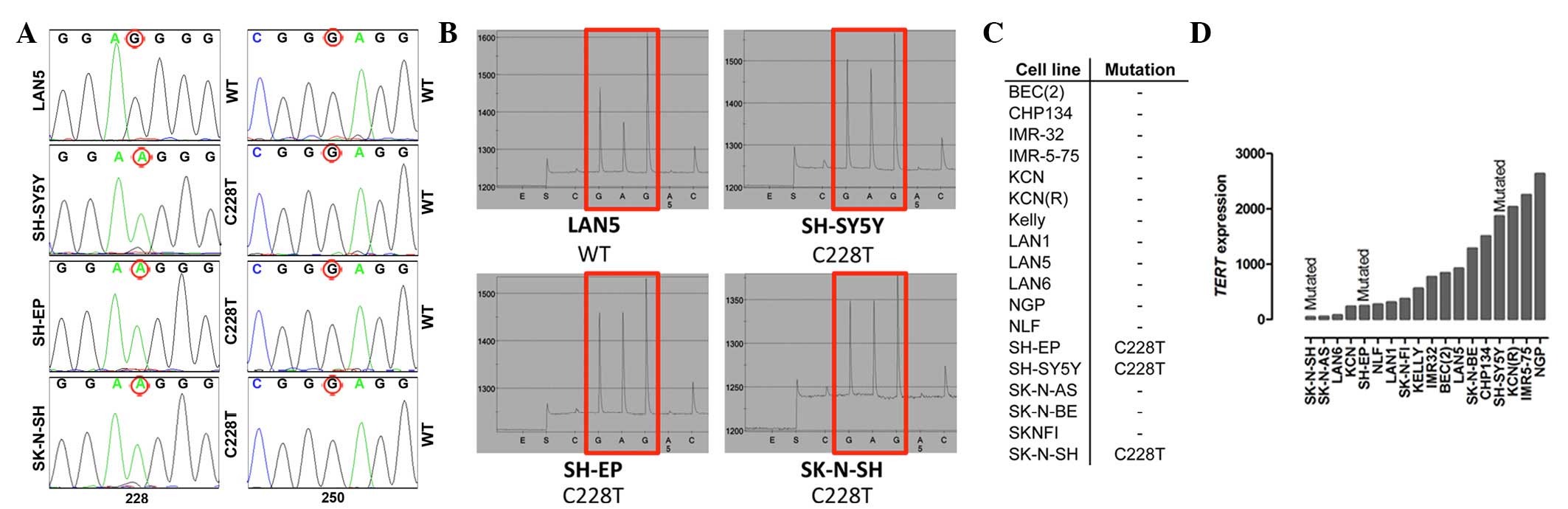
detected. The neuroblastoma cell lines SK-N-SH, SH-SY5Y and SH-EP
harbor the C228T mutation (Fig. 1A–C).
Of note, SH-SY5Y and SH-EP are subclones of SK-N-SH (20). SK-N-SH was established from a bone
marrow biopsy of a 4-year-old girl (21,22). The
presence of the C228T TERT promoter mutation in SK-N-SH,
SH-SY5Y and SH-EP was independently validated using pyrosequencing
(Fig. 1B). As the C228T mutation
appeared to be homozygous in SK-N-SH, SH-SY5Y and SH-EP, Affymetrix
Human SNP array 6.0 arrays were used to analyze zygosity. The 3
probes located within the TERT gene were homozygeous in all
three cell lines, which could point to the presence of a small
loss-of-heterozygosity region spanning the TERT gene
(Table II). To analyze if the
mutations in the TERT promoter correlate with increased
TERT expression, the expression was analyzed in all the
neuroblastoma cell lines using RT-qPCR. No correlation between
mutations and TERT expression was detected (Fig. 1D).
| Table II.Location of the Affymetrix CytoScanHD
array probes located within or near the TERT gene. The
probes lying within the TERT gene showed a loss of heterozygousity
for SH-EP, SH-SY5Y and SK-N-SH. Probes lying outside of the TERT
gene showed a heterozygous status. |
Table II.
Location of the Affymetrix CytoScanHD
array probes located within or near the TERT gene. The
probes lying within the TERT gene showed a loss of heterozygousity
for SH-EP, SH-SY5Y and SK-N-SH. Probes lying outside of the TERT
gene showed a heterozygous status.
|
|
| Zygosity
status |
|
|---|
|
|
|
|
|
|---|
| Probe ID | Probe start
position | SK-N-SH | SH-EP | SH-SY5Y | Comment |
|---|
| S-4ELKX | 1240103 | Heterozygous | Heterozygous | Heterozygous | Probe downstream of
TERT gene |
| S-4OOCR | 1267356 | Homozygous | Homozygous | Homozygous | Probe inside of
TERT gene |
| S-4FACZ | 1269006 | Homozygous | Homozygous | Homozygous | Probe inside of
TERT gene |
| S-4IBBK | 1269365 | Homozygous | Homozygous | Homozygous | Probe inside of
TERT gene |
| S-3MNHV | 1299087 | Heterozygous | Heterozygous | Heterozygous | Probe upstream of
TERT gene |
Discussion
Analysis of the TERT gene promotor region in
20 neuroblastoma cell lines and 131 primary neuroblastoma tumors
revealed mutations in 3 cell lines and no mutations in primary
tumors.
Of note, the only 3 mutated cell lines, which harbor
the C228T mutation, are SK-N-SH and its 2 subclones SH-EP and
SH-SY5Y, further limiting the general indication of this finding.
It is uncommon that TERT mutations occur homozygously, as
detected in SK-N-SH, SH-EP and SY5Y. However, further analysis
confirmed a loss-of-heterozygosity region in the region of the
TERT gene, pointing to a hemizygous rather than a homozygous
status of the C228T mutation in these cases. Taking into
consideration the variable levels of TERT expression in SK-N-SH,
SH-EP and SH-SY5Y cells, the functional indication of the C228T
mutations in these cell lines remain questionable.
The absence of TERT mutations in primary
neuroblastomas and the majority of analyzed cell lines is
unexpected, as TERT expression is of pathogenic relevance in
the majority of neuroblastomas (13)
and TERT mutations are frequent in related neuroectodermal
tumors, such as melanoma and medulloblastoma (4,7,23,24).
However, the present results are in line with previous studies by
Papathomas et al (25), who did
not find TERT promoter mutations in 15 primary
neuroblastomas and Killela et al (23), who identified only 2 TERT
promoter mutations in 22 neuroblastomas.
The mechanisms by which TERT expression is
induced, particularly in MYCN non-amplified high-risk
neuroblastomas, remains elusive and warrants further studies.
Potential alternative mechanisms underlying TERT induction
in neuroblastoma include epigenetic changes, as well as mutations
in more distant regulatory elements that were not analyzed in the
present study.
Taken together, we conclude that TERT (core)
promoter mutations are not relevant events in neuroblastoma
pathogenesis.
Acknowledgements
The authors acknowledge the funding from the German
Ministry for Education and Research (e:MED grant nos. SMOOSE FKZ:
01ZX1303B and SYSMED-NB FKZ: 01ZX1307E to J.H.S.) and the German
Cancer Aid (grant no. 111301 to J.H.S.).
References
|
1
|
Shay JW and Wright WE: Role of telomeres
and telomerase in cancer. Semin Cancer Biol. 21:349–353. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Nabetani A and Ishikawa F: Alternative
lengthening of telomeres pathway: Recombination-mediated telomere
maintenance mechanism in human cells. J Biochem. 149:5–14. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Griewank KG, Murali R, Schilling B, Scholz
S, Sucker A, Song M, Süsskind D, Grabellus F, Zimmer L, Hillen U,
et al: TERT promoter mutations in ocular melanoma distinguish
between conjunctival and uveal tumours. Br J Cancer. 109:497–501.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Horn S, Figl A, Rachakonda PS, Fischer C,
Sucker A, Gast A, Kadel S, Moll I, Nagore E, Hemminki K, et al:
TERT promoter mutations in familial and sporadic melanoma. Science.
339:959–961. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Huang FW, Hodis E, Xu MJ, Kryukov GV, Chin
L and Garraway LA: Highly recurrent TERT promoter mutations in
human melanoma. Science. 339:957–959. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Xu D, Dwyer J, Li H, Duan W and Liu JP:
Ets2 maintains hTERT gene expression and breast cancer cell
proliferation by interacting with c-Myc. J Biol Chem.
283:23567–23580. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Remke M, Ramaswamy V, Peacock J, Shih DJ,
Koelsche C, Northcott PA, Hill N, Cavalli FM, Kool M, Wang X, et
al: TERT promoter mutations are highly recurrent in SHH subgroup
medulloblastoma. Acta Neuropathol. 126:917–929. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Weiss WA, Aldape K, Mohapatra G,
Feuerstein BG and Bishop JM: Targeted expression of MYCN causes
neuroblastoma in transgenic mice. EMBO J. 16:2985–2995. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Maris JM, Hogarty MD, Bagatell R and Cohn
SL: Neuroblastoma. Lancet. 369:2106–2120. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Pugh TJ, Morozova O, Attiyeh EF,
Asgharzadeh S, Wei JS, Auclair D, Carter SL, Cibulskis K, Hanna M,
Kiezun A, et al: The genetic landscape of high-risk neuroblastoma.
Nat Genet. 45:279–284. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
Molenaar JJ, Koster J, Zwijnenburg DA, van
Sluis P, Valentijn LJ, van der Ploeg I, Hamdi M, van Nes J,
Westerman BA, van Arkel J, et al: Sequencing of neuroblastoma
identifies chromothripsis and defects in neuritogenesis genes.
Nature. 483:589–593. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Cheung NK, Zhang J, Lu C, Parker M,
Bahrami A, Tickoo SK, Heguy A, Pappo AS, Federico S, Dalton J, et
al: St Jude Children's Research Hospital-Washington University
Pediatric Cancer Genome Project: Association of age at diagnosis
and genetic mutations in patients with neuroblastoma. JAMA.
307:1062–1071. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Poremba C, Scheel C, Hero B, Christiansen
H, Schaefer KL, Nakayama Ji, Berthold F, Juergens H, Boecker W and
Dockhorn-Dworniczak B: Telomerase activity and telomerase subunits
gene expression patterns in neuroblastoma: A molecular and
immunohistochemical study establishing prognostic tools for
fresh-frozen and paraffin-embedded tissues. J Clin Oncol.
18:2582–2592. 2000.PubMed/NCBI
|
|
14
|
Brodeur GM and Bagatell R: Mechanisms of
neuroblastoma regression. Nat Rev Clin Oncol. 11:704–713. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Mac SM, D'Cunha CA and Farnham PJ: Direct
recruitment of N-myc to target gene promoters. Mol Carcinog.
29:76–86. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Schulte JH, Horn S, Otto T, et al: MYCN
regulates oncogenic MicroRNAs in neuroblastoma. Int J Cancer.
122:699–704. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Cobrinik D, Ostrovnaya I, Hassimi M,
Tickoo SK, Cheung IY and Cheung NK: Recurrent pre-existing and
acquired DNA copy number alterations, including focal TERT gains,
in neuroblastoma central nervous system metastases. Genes
Chromosomes Cancer. 52:1150–1166. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Vandesompele J, De Preter K, Pattyn F, et
al: Accurate normalization of real-time quantitative RT-PCR data by
geometric averaging of multiple internal control genes. Genome
Biol. 3:RESEARCH0034. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Bachmann HS, Heukamp LC, Schmitz KJ, et
al: Regulatory BCL2 promoter polymorphism (−938C>A) is
associated with adverse outcome in patients with prostate
carcinoma. International journal of cancer. Int J Cancer.
129:2390–2399. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sadée W, Yu VC, Richards ML, Preis PN,
Schwab MR, Brodsky FM and Biedler JL: Expression of
neurotransmitter receptors and myc protooncogenes in subclones of a
human neuroblastoma cell line. Cancer Res. 47:5207–5212.
1987.PubMed/NCBI
|
|
21
|
Barnes EN, Biedler JL, Spengler BA and
Lyser KM: The fine structure of continuous human neuroblastoma
lines SK-N-SH, SK-N-BE(2) and SK-N-MC. In Vitro. 17:619–631. 1981.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Biedler JL, Helson L and Spengler BA:
Morphology and growth, tumorigenicity and cytogenetics of human
neuroblastoma cells in continuous culture. Cancer Res.
33:2643–2652. 1973.PubMed/NCBI
|
|
23
|
Killela PJ, Reitman ZJ, Jiao Y, Bettegowda
C, Agrawal N, Diaz LA Jr, Friedman AH, Friedman H, Gallia GL,
Giovanella BC, et al: TERT promoter mutations occur frequently in
gliomas and a subset of tumors derived from cells with low rates of
self-renewal. Proc Natl Acad Sci USA. 110:6021–6026. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lindsey JC, Schwalbe EC, Potluri S, Bailey
S, Williamson D and Clifford SC: TERT promoter mutation and
aberrant hypermethylation are associated with elevated expression
in medulloblastoma and characterise the majority of non-infant SHH
subgroup tumours. Acta Neuropathol. 127:307–309. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Papathomas TG, Oudijk L, Zwarthoff EC,
Post E, Duijkers FA, van Noesel MM, Hofland LJ, Pollard PJ, Maher
ER, Restuccia DF, et al: Telomerase reverse transcriptase promoter
mutations in tumors originating from the adrenal gland and
extra-adrenal paraganglia. Endocr Relat Cancer. 21:653–661. 2014.
View Article : Google Scholar : PubMed/NCBI
|