Introduction
There is no curative treatment for advanced renal
cancer. Innovative treatments using tyrosine kinase inhibitors and
inhibitors of the mammalian target of rapamycin have been tested,
however, they were not curative (1–3). Development
of a novel treatment approach is urgently required.
Inducing endoplasmic reticulum (ER) stress and
ubiquitinated protein accumulation has emerged as a new approach
for the treatment of malignancies (4).
How effectively ER stress can be induced depends on how effectively
unfolded proteins can be accumulated in the cell. Combining a
proteasome inhibitor and histone deacetylase (HDAC) inhibitor is
one efficient way to induce ER stress. HDAC inhibitors generally
inhibit HDAC6, and ablation of HDAC6 induces hyperacetylation of
heat-shock protein (HSP) 90, disrupting its chaperone function
(5) and increasing unfolded proteins
in the cell. These unfolded proteins, however, are normally
degraded by the proteasome (6), so a
proteasome inhibitor in combination with an HDAC inhibitor is
expected to effectively accumulate unfolded proteins in the cell by
inhibiting degradation of the unfolded proteins increased by the
HDAC inhibitor. Our previous study revealed that the combinations
of the proteasome inhibitor bortezomib and the HDAC inhibitor
suberoylanilide hydroxamic acid (7) or
panobinostat (8) induced ER stress and
inhibited renal cancer growth synergistically. Combining bortezomib
and HDAC inibitors would thus be a promising approach to causing ER
stress in renal cancer cells.
Belinostat is a novel pandeacetylase inhibitor that
has been clinically tested in patients with solid tumors, such as
liver cancer (9), ovarian cancer
(10), malignant pleural mesothelioma
(11) and thymic epithelial tumors
(12), as well as in patients with
hematological malignancies (13,14).
Favorable antineoplastic effects of belinostat in combination with
bortezomib have been reported (15–18),
however, there has been no study evaluating the effect of the
combination on renal cancer cells and investigating ER stress
induction as a mechanism of action. Furthermore, belinostat is the
only HDAC inhibitor that can be administered in multiple ways
(i.e., intravenously or orally) (19)
and may thus be easier to use than other HDAC inhibitors.
In the present study, the efficacy of the
combination of bortezomib and belinostat was evaluated in renal
cancer cells and its underlying mechanism of action was clarified,
focusing on ER stress induction and ubiquitinated protein
accumulation.
Materials and methods
Cell lines
Renal cancer cell lines (769-P, 786-O and ACHN) were
purchased from American Type Culture Collection (Rockville, MD,
USA). Cells were grown in either RPMI or DMEM medium containing 10%
fetal bovine serum and 0.3% penicillin-streptomycin (Invitrogen,
Carlsbad, CA, USA).
Reagents
Belinostat and bortezomib were purchased from
Selleck Chemicals (Houston, TX, USA) and LC Laboratories (Woburn,
MA, USA), respectively. They were dissolved in dimethyl sulfoxide
and stored at −20°C until use. Z-VAD-FMK was obtained from Enzo
Life Sciences (Farmingdale, NY, USA).
Cell viability assay
In total, 5×103 cells were seeded in a
96-well culture plate one day before treatment for 48 h with 0–2 µM
belinostat and either 0, 5, or 10 nM bortezomib. Cell viability was
subsequently assayed using the MTS assay (CellTiter 96 AQueous kit;
Promega Madison, WI, USA) according to the manufacturer's
protocol.
Colony formation assay
A total of 100 cells were seeded in 6-well plates
one day before culture for 48 h in media containing 1 µM belinostat
and/or 10 nM bortezomib. The cells were subsequently provided fresh
media and allowed to grow for one week before being fixed with 100%
methanol, stained with Giemsa's solution and counted.
Cell cycle analysis
In total, 1.5×105 cells were plated in
6-well culture plates one day before treatment for 48 h with 2 µM
belinostat and/or 10 nM bortezomib, washed with phosphate-buffered
saline and harvested by trypsinization. The cells were resuspended
in citrate buffer and stained with propidium iodide according to
the procedure used by Vindeløv et al (20), and were analyzed by flow cytometry.
Annexin V assay
A total of 1.5×105 cells were plated in
6-well culture plates one day before culture for 48 h in media
containing 2 µM belinostat and 10 nM bortezomib with or without 40
µM of the pancaspase inhibitor Z-VAD-FMK. The cells were
subsequently stained with Annexin V according to the manufacturer's
protocol (Beckman Coulter, Marseille, France). In total, 10,000
cells were counted and analyzed by flow cytometry.
Western blotting
Equal amounts of protein were subjected to SDS-PAGE
and transferred to a nitrocellulose membrane. Following blocking
with 5% skimmed milk, the membrane was incubated with a rabbit
polyclonal antibody against glucose-regulated protein (GRP) 78
(1:200), a rabbit polyclonal antibody against ubiquitin (1:200)
(Santa Cruz Biotechnology, Dallas, TX, USA), a mouse monoclonal
antibody against actin (1:3,000) (Millipore, Billerica, MA, USA), a
rabbit polyclonal antibody against HSP70 (1:1,000) and a rabbit
polyclonal antibody against cleaved poly (ADP-ribose) polymerase
(PARP) (1:1,000) (Cell Signaling Technology, Danvers, MA, USA). The
samples were subsequently incubated with
horseradish-peroxidase-tagged secondary antibodies (Bio-Rad,
Hercules, CA, USA) and visualization was performed by
chemiluminescence (ECL, Amersham, Piscataway, NJ, USA).
Statistical analysis
Evaluation of synergism was carried out using
commercially available software (CalcuSyn, Biosoft, Cambridge, UK).
The statistical significance of the observed differences in the
colony formation assay was evaluated using the Mann-Whitney U test
(StatView software, SAS Institute, Cary, NC, USA). P<0.05 was
considered to indicate a statistically significant difference.
Results
Bortezomib and belinostat
synergistically inhibit renal cancer growth
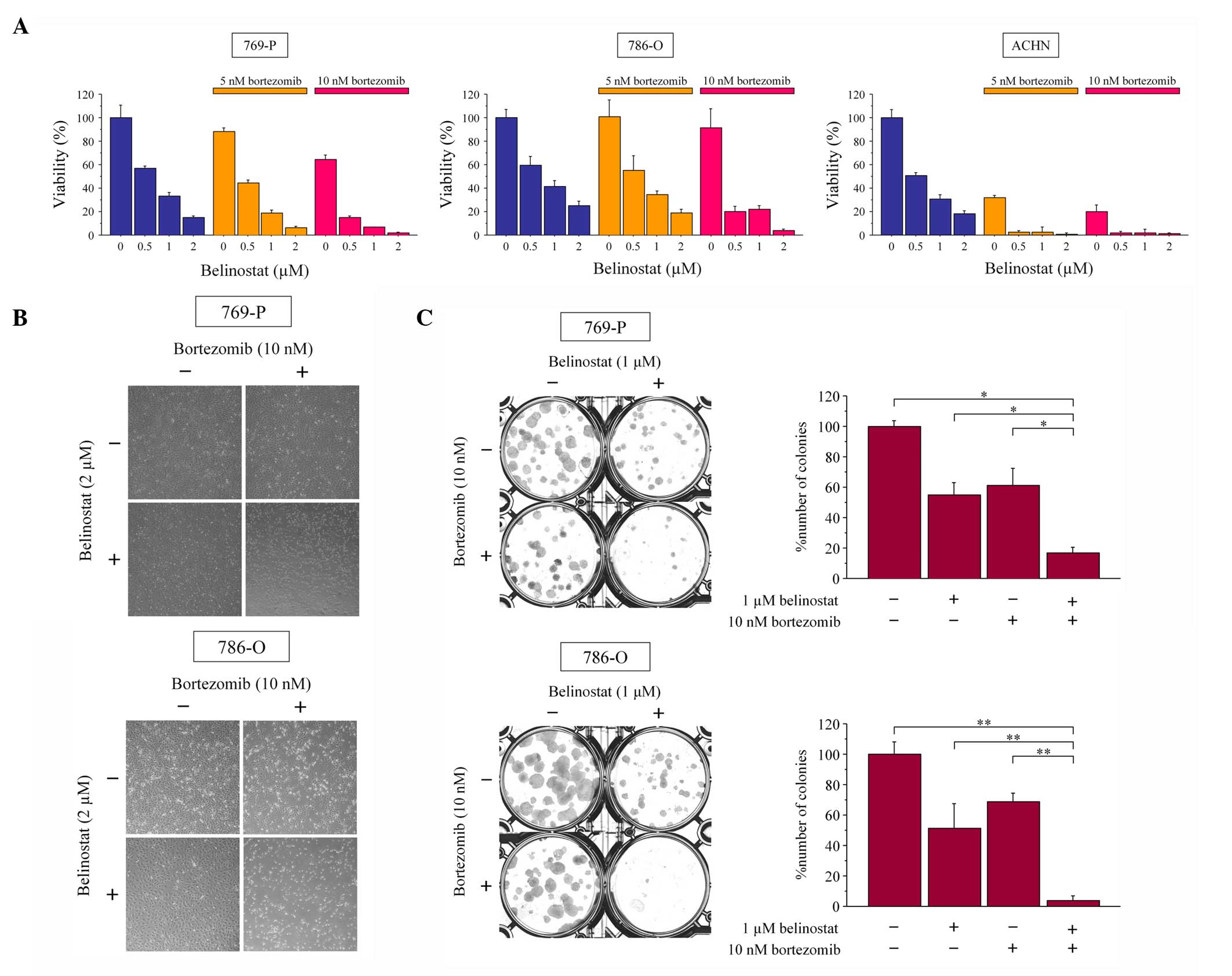
The combined effect of bortezomib and belinostat
were first examined on cell viability. Bortezomib and belinostat
each inhibited the growth of renal cancer cells in a dose-dependent
manner and the combination inhibited their growth more than
treatment with the agents individually (Fig. 1A and B). Subsequently, the interaction
of these two agents was evaluated using the Chou-Talalay method to
calculate the combination indexes. Indexes <1 showed that their
interaction was synergistic under a number of the treatment
conditions (Table I). Whether the
combination of bortezomib and belinostat interferes with colony
formation by renal cancer cells was subsequently investigated. The
combination suppressed colony formation significantly more than 10
nM bortezomib or 1 µM belinostat alone (Fig. 1C). Therefore, the combination of
bortezomib and belinostat was shown to inhibit renal cancer growth
effectively.
 | Table I.CI for the combination of bortezomib
and belinostat. |
Table I.
CI for the combination of bortezomib
and belinostat.
|
|
| Bortezomib, nM |
|---|
|
|
|
|
|---|
| Cell line | Belinostat, µM | 5 | 10 |
|---|
| 769-P | 0.5 | 1.029 | 0.529a |
|
| 1 | 0.685a | 0.405a |
|
| 2 | 0.579a | 0.286a |
| 786-O | 0.5 | 1.287 | 0.949a |
|
| 1 | 1.084 | 1.142 |
|
| 2 | 1.099 | 0.813a |
| ACHN | 0.5 | 0.075a | 0.093a |
|
| 1 | 0.115a | 0.127a |
|
| 2 | 0.073a | 0.129a |
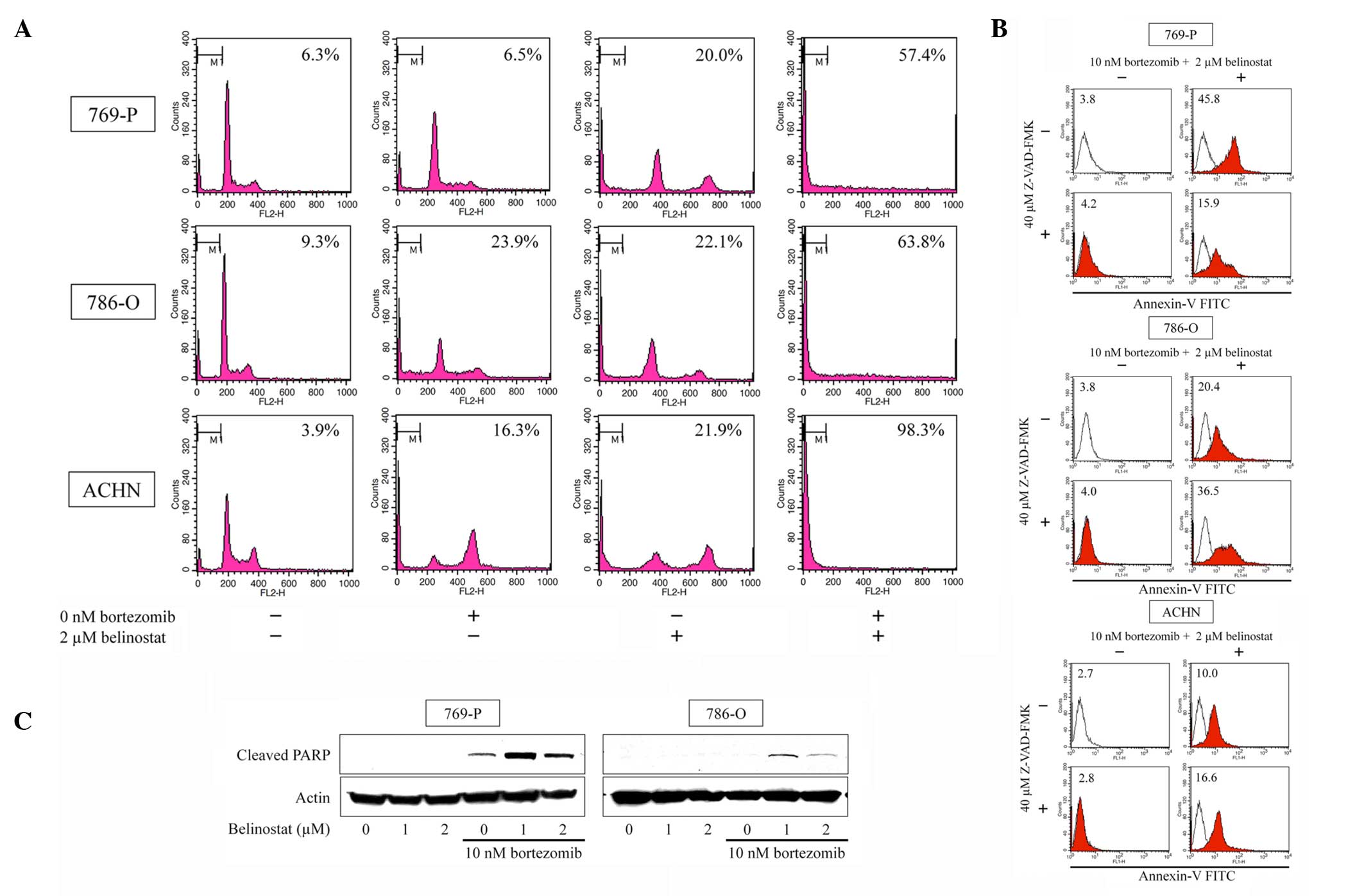
Bortezomib and belinostat induce
apoptosis
Cell cycle analysis and the Annexin V assay were
used to observe whether the combination of bortezomib and
belinostat induced apoptosis. Cell cycle analysis (Fig. 2A) showed that the combination of 10 nM
bortezomib and 2 µM belinostat synergistically increased the number
of the cells in the sub-G1 fraction (≤98.3%). The
combination also markedly increased Annexin V-FITC fluorescence
intensity (Fig. 2B) and increased the
expression of cleaved PARP (Fig. 2C).
Thus, the combination was shown to induce apoptosis. Whether the
combination-induced apoptosis is associated with caspase activation
was also examined. As co-treatment with a 40 µM concentration of
the pan-caspase inhibitor Z-VAD-FMK changed the number of Annexin
V-positive cells (reduced in 769-P cells and increased in 786-O and
ACHN cells), the apoptosis was considered, at least in part,
caspase dependent (Fig. 2B).
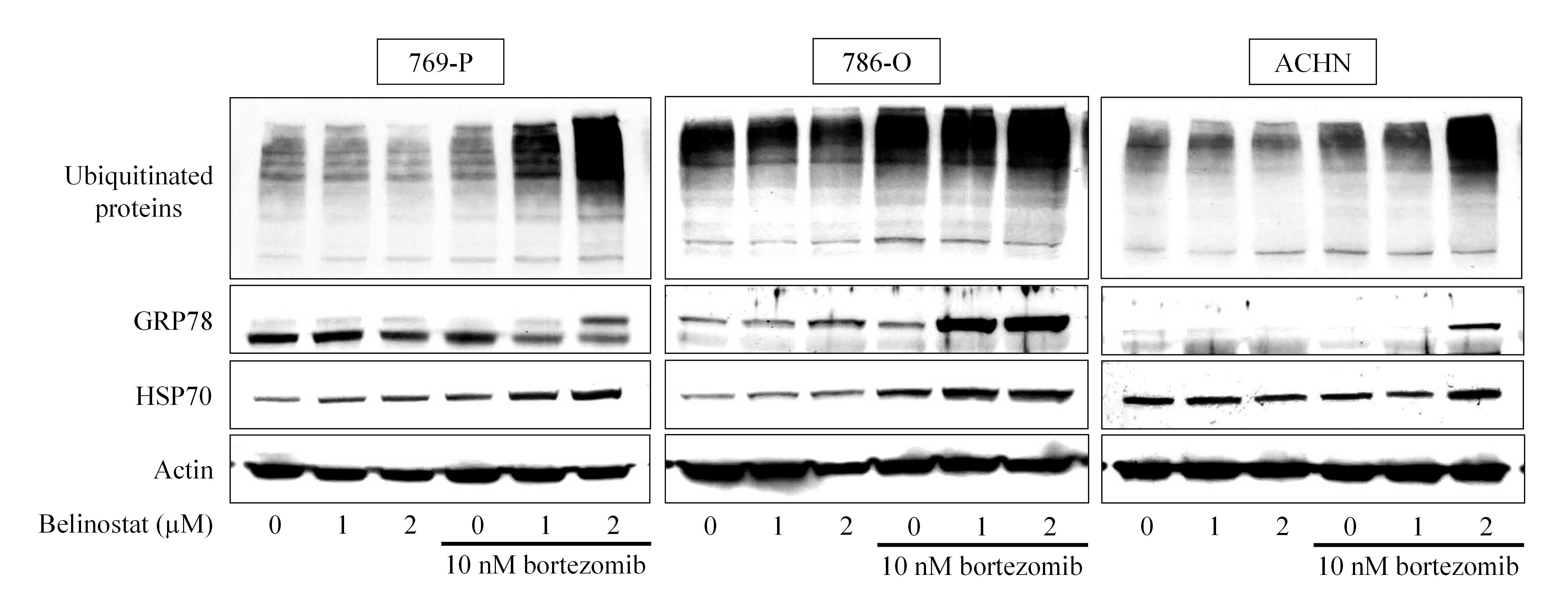
Bortezomib and belinostat cause
ubiquitinated protein accumulation and ER stress
Our hypothesis was that the combination of
bortezomib and belinostat would increase ubiquitinated proteins in
the cell and induce ER stress cooperatively, as belinostat would
increase the amount of unfolded proteins and bortezomib would
inhibit their degradation by inhibiting the proteasome. As
expected, the combination synergistically caused ubiquitinated
proteins to accumulate (Fig. 3). In
all three cell lines, no accumulation of ubiquitinated protein was
induced by 2 µM belinostat alone; however, in combination with 10
nM bortezomib, marked accumulation of ubiquitinated protein was
induced. By contrast, 10 nM bortezomib caused small (in 769-P and
ACHN cells) and moderate (in 786 cells) ubiquitinated protein
accumulation that was markedly enhanced by belinostat. The
combination also increased the expression of the ER stress markers
GRP78 and HSP70, confirming that the combination actually caused
ubiquitinated unfolded protein accumulation and thereby induced ER
stress.
Discussion
There is no curative therapy for advanced renal
cancer, and a novel treatment approach is urgently required.
Inducing ubiquitinated protein accumulation and ER stress is a new
approach to the treatment of malignancies (4) and our laboratory has been investigating
ways to kill urological cancer cells using clinically feasible
drugs to induce ER stress. To the best of our knowledge, for the
first time the favorable combined effect of the proteasome
inhibitor bortezomib and the HDAC inhibitor belinostat in renal
cancer was revealed in the present study.
Combining a proteasome inhibitor and HDAC inhibitor
is one effective way to induce ER stress, as HDAC inhibitors are
considered to increase unfolded proteins by suppressing HSP90
function (5) and proteasome inhibitors
inhibit the degradation of those unfolded proteins. This strategy
has been shown to be effective in killing renal cancer cells
(7,8).
The present analysis using the Chou-Talalay method
showed that the combination of bortezomib and belinostat inhibited
renal cancer growth synergistically. It caused ubiquitinated
protein accumulation and ER stress cooperatively: 2 µM belinostat
induced marked accumulation of ubiquitinated protein only when it
was combined with 10 nM bortezomib. Therefore, the increased amount
of unfolded proteins due to 2 µM belinostat alone did not overwhelm
the protein degrading ability of the proteasome; i.e., they were
rapidly degraded by the proteasome and therefore did not
accumulate. By contrast, in 769-P and ACHN cells, 10 nM bortezomib
alone caused only small ubiquitinated protein accumulation;
however, in combination with belinostat it caused marked
ubiquitinated protein accumulation. Thus, in these cell lines the
amount of unfolded protein must be increased for effective
ubiquitinated protein accumulation, even though the proteasome is
suppressed. Simultaneous inhibition of HSP90 and proteasome
function is therefore of importance in causing unfolded proteins to
accumulate and inducing enough ER stress to kill cancer cells. As
the proteasomes in renal cancer cells are overexpressed (21), this proteasome inhibition is considered
particularly important when killing renal cancer cells by inducing
ER stress.
As indicated by the cell cycle analysis, Annexin V
assay and western blot analysis detecting cleaved PARP, the
combination induced apoptosis effectively. This apoptosis is
considered to be associated with caspase activation as the
pancaspase inhibitor Z-VAD-FMK affected the number of Annexin
V-positive cells caused by the combination. Notably, the effect of
Z-VAD-FMK differed among the cell lines: Decreasing the number of
Annexin V-positive cells in 769-P cells and increasing it in 786-O
and ACHN cells. Therefore, in 786-O and ACHN cells the inhibition
of caspase paradoxically promoted apoptosis. A similar phenomenon
was reported by Liu et al (22)
who showed that Z-VAD-FMK sensitized neutrophils to tumor necrosis
factor-α-induced apoptosis using the Annexin V assay. The study
hypothesized that the caspase-dependent pathway normally suppresses
or supersedes caspase-independent mechanisms; however,
understanding the exact mechanism of this paradoxical phenomenon
will require further study.
One limitation of the present study is that
suppression of HSP90 function by belinostat was not shown.
Belinostat individually induced histone acetylation at the
indicated concentrations, so it is evident that belinostat
suppressed the function of HDACs, and we hypothesized that
belinostat would therefore cause HSP90 acetylation, leading to its
functional suppression. However, formal functional evaluation of
HSP90 under belinostat treatment would be the subsequent step in
clarifying the actual mechanism of action of belinostat in this
combination therapy.
In conclusion, the bortezomib-belinostat combination
induced ubiquitinated protein accumulation and ER stress, and
killed renal cancer cells synergistically. To the best of our
knowledge, this is the first study to demonstrate the favorable
combined effect of bortezomib and belinostat in renal cancer cells.
As this combination kills renal cancer cells in a way that is
completely different from the non-curative treatments currently
used, we hypothesize that the present study, along with our
previous studies, provides a framework for testing combinations of
proteasome inhibitors and HDAC inhibitors in patients with advanced
renal cancer.
References
|
1
|
Escudier B, Eisen T, Stadler WM, Szczylik
C, Oudard S, Siebels M, Negrier S, Chevreau C, Solska E, Desai AA,
et al: TARGET Study Group: Sorafenib in advanced clear-cell
renal-cell carcinoma. N Engl J Med. 356:125–134. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Motzer RJ, Rini BI, Bukowski RM, Curti BD,
George DJ, Hudes GR, Redman BG, Margolin KA, Merchan JR, Wilding G,
et al: Sunitinib in patients with metastatic renal cell carcinoma.
JAMA. 295:2516–2524. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Motzer RJ, Escudier B, Oudard S, Hutson
TE, Porta C, Bracarda S, Grünwald V, Thompson JA, Figlin RA,
Hollaender N, et al: RECORD-1 Study Group: Phase 3 trial of
everolimus for metastatic renal cell carcinoma: Final results and
analysis of prognostic factors. Cancer. 116:4256–4265. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Liu Y and Ye Y: Proteostasis regulation at
the endoplasmic reticulum: A new perturbation site for targeted
cancer therapy. Cell Res. 21:867–883. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bali P, Pranpat M, Bradner J, Balasis M,
Fiskus W, Guo F, Rocha K, Kumaraswamy S, Boyapalle S, Atadja P, et
al: Inhibition of histone deacetylase 6 acetylates and disrupts the
chaperone function of heat shock protein 90: A novel basis for
antileukemia activity of histone deacetylase inhibitors. J Biol
Chem. 280:26729–26734. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Naujokat C and Hoffmann S: Role and
function of the 26S proteasome in proliferation and apoptosis. Lab
Invest. 82:965–980. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sato A and Asano T, Ito K, Sumitomo M and
Asano T: Suberoylanilide hydroxamic acid (SAHA) combined with
bortezomib inhibits renal cancer growth by enhancing histone
acetylation and protein ubiquitination synergistically. BJU Int.
109:1258–1268. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sato A and Asano T, Isono M, Ito K and
Asano T: Panobinostat synergizes with bortezomib to induce
endoplasmic reticulum stress and ubiquitinated protein accumulation
in renal cancer cells. BMC Urol. 14:712014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yeo W, Chung HC, Chan SL, Wang LZ, Lim R,
Picus J, Boyer M, Mo FK, Koh J, Rha SY, et al: Epigenetic therapy
using belinostat for patients with unresectable hepatocellular
carcinoma: A multicenter phase I/II study with biomarker and
pharmacokinetic analysis of tumors from patients in the Mayo Phase
II Consortium and the Cancer Therapeutics Research Group. J Clin
Oncol. 30:3361–3367. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Mackay HJ, Hirte H, Colgan T, Covens A,
MacAlpine K, Grenci P, Wang L, Mason J, Pham PA, Tsao MS, et al:
Phase II trial of the histone deacetylase inhibitor belinostat in
women with platinum resistant epithelial ovarian cancer and
micropapillary (LMP) ovarian tumours. Eur J Cancer. 46:1573–1579.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ramalingam SS, Belani CP, Ruel C, Frankel
P, Gitlitz B, Koczywas M, Espinoza-Delgado I and Gandara D: Phase
II study of belinostat (PXD101), a histone deacetylase inhibitor,
for second line therapy of advanced malignant pleural mesothelioma.
J Thorac Oncol. 4:97–101. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Giaccone G, Rajan A, Berman A, Kelly RJ,
Szabo E, Lopez-Chavez A, Trepel J, Lee MJ, Cao L, Espinoza-Delgado
I, et al: Phase II study of belinostat in patients with recurrent
or refractory advanced thymic epithelial tumors. J Clin Oncol.
29:2052–2059. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Cashen A, Juckett M, Jumonville A, Litzow
M, Flynn PJ, Eckardt J, LaPlant B, Laumann K, Erlichman C and
DiPersio J: Phase II study of the histone deacetylase inhibitor
belinostat (PXD101) for the treatment of myelodysplastic syndrome
(MDS). Ann Hematol. 91:33–38. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Gimsing P, Hansen M, Knudsen LM, Knoblauch
P, Christensen IJ, Ooi CE and Buhl-Jensen P: A phase I clinical
trial of the histone deacetylase inhibitor belinostat in patients
with advanced hematological neoplasia. Eur J Haematol. 81:170–176.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Paoluzzi L, Scotto L, Marchi E, Zain J,
Seshan VE and O'Connor OA: Romidepsin and belinostat synergize the
antineoplastic effect of bortezomib in mantle cell lymphoma. Clin
Cancer Res. 16:554–565. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Spratlin JL, Pitts TM, Kulikowski GN,
Morelli MP, Tentler JJ, Serkova NJ and Eckhardt SG: Synergistic
activity of histone deacetylase and proteasome inhibition against
pancreatic and hepatocellular cancer cell lines. Anticancer Res.
31:1093–1103. 2011.PubMed/NCBI
|
|
17
|
Dai Y, Chen S, Wang L, Pei XY, Kramer LB,
Dent P and Grant S: Bortezomib interacts synergistically with
belinostat in human acute myeloid leukaemia and acute lymphoblastic
leukaemia cells in association with perturbations in NF-κB and Bim.
Br J Haematol. 153:222–235. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Feng R, Oton A, Mapara MY, Anderson G,
Belani C and Lentzsch S: The histone deacetylase inhibitor, PXD101,
potentiates bortezomib-induced anti-multiple myeloma effect by
induction of oxidative stress and DNA damage. Br J Haematol.
139:385–397. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lemoine M and Younes A: Histone
deacetylase inhibitors in the treatment of lymphoma. Discov Med.
10:462–470. 2010.PubMed/NCBI
|
|
20
|
Vindeløv LL, Christensen IJ and Nissen NI:
A detergent-trypsin method for the preparation of nuclei for flow
cytometric DNA analysis. Cytometry. 3:323–327. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kanayama H, Tanaka K, Aki M, Kagawa S,
Miyaji H, Satoh M, Okada F, Sato S, Shimbara N and Ichihara A:
Changes in expressions of proteasome and ubiquitin genes in human
renal cancer cells. Cancer Res. 51:6677–6685. 1991.PubMed/NCBI
|
|
22
|
Liu CY, Takemasa A, Liles WC, Goodman RB,
Jonas M, Rosen H, Chi E, Winn RK, Harlan JM and Chuang PI:
Broad-spectrum caspase inhibition paradoxically augments cell death
in TNF-alpha-stimulated neutrophils. Blood. 101:295–304. 2003.
View Article : Google Scholar : PubMed/NCBI
|