Introduction
Liver cancer remains the fifth most common
malignancy worldwide, and its incidence is on the rise due to
hepatitis virus infection (1,2). The
incidence of liver cancer is particularly high in China and
Southeast Asian countries. Current treatments for liver cancer
mainly include early surgical treatment, intervention, liver
transplantation and targeted therapy (2,3).
Since liver cancer originates in an occult fashion and progresses
rapidly, most patients are at the late stage at diagnosis and are
no longer good candidates for surgical removal (3,4). In
addition, the clinical benefits of intervention or targeted therapy
are limited in most patients (5,6).
Therefore, the development of new paradigms for liver cancer
treatment is crucial.
In China, traditional Chinese medicine has been
widely used in all aspects of liver cancer, including surgery,
interventional and targeted therapy, even as a single treatment for
advanced liver cancer, and has been confirmed to effectively
control cancer progression, improve quality of life and prolong
survival times, to some extent, in liver cancer patients (7–10).
Liver Yin deficiency is one of the common syndromes included in or
accompanying liver cancer (11,12).
Liver Yin tonifying herbs have been used for the treatment of liver
cancer with liver Yin deficiency and are crucial elements of
traditional Chinese medicine prescriptions (13,14).
However, the clinical application of liver Yin tonifying herbs is
empirical to some extent, and there is a lack of an efficient liver
Yin tonifying herbal formula for liver cancer, limiting the
improvement of its efficacy.
Based on the principles of traditional Chinese
medicine and progress in the pharmacological characterization of
anticancer herbs, we developed a liver Yin tonifying herbal formula
(LYTF) for liver cancer. Our LYTF consists of glossy privet fruit,
Carapax trionycis (processed with vinegar) and Polygonum
cuspidatum. Glossy privet fruit and Carapax trionycis
are classic liver Yin tonifying Chinese herbs that are frequently
used in the treatment of liver cancer and have demonstrated
anticancer effects to some extent (15–17).
Oleanolic acid, the main component of glossy privet fruit, has
recently been reported to induce the apoptosis of hepatocarcinoma
cells (18). Polygonum
cuspidatum has anti-inflammatory, anti-viral, anti-tumor and
anti-angiogenic effects, and is commonly employed in treatments for
viral hepatitis and liver cancer (10,19).
Resveratrol, the major component from Polygonum cuspidatum,
exerts desirable anticancer effects by inducing apoptosis and cell
senescence in numerous types of cancer cells (20–22).
In the present study, we observed that LYTF inhibited
proliferation, enhanced caspase activity and induced apoptosis in
Bel-7402 hepatocarcinoma cells. In addition, LYTF induced cell
senescence in Bel-7402 cells, accompanied by the up-regulation of
p16 and p21 expression and the down-regulation of retinoblastoma
(Rb) protein phosphorylation.
Materials and methods
Chemicals and reagents
RPMI-1640 medium and fetal bovine serum (FBS) were
obtained from Hyclone (Logan, UT, USA). The Cell Counting Kit-8
(CCK8) was purchased from Dojindo (Kumamoto, Japan). The FITC
Annexin V Apoptosis Detection kit was obtained from BD Biosciences
Pharmingen (San Diego, CA, USA). The Colorimetric CaspACE™ Assay
System was the product of Promega (Madison, WI, USA). Z-VAD-FMK,
Caspase-8 and Caspase-9 Colorimetric Assay kits were procured from
R&D Systems (Minneapolis, MN, USA). Antibodies against p16,
p21, Rb, phospho-Rb, poly(ADP-ribose)polymerase (PARP) and
α-tubulin were the product of Cell Signaling Technology (Danvers,
MA, USA). The Hoechst Staining kit was obtained from Beyotime
(Jiangsu, China). The Senescence β-Gal Staining kit was purchased
from Cell Signaling Technology.
Herbal preparation
LYTF was prepared as a lyophilized dry powder of hot
water extracts as described previously (23–25).
Authentic herb materials were from the Longhua Hospital herb store.
The herbs used were glossy privet fruit (24 g), Carapax
trionycis (processed with vinegar) (60 g) and Polygonum
cuspidatum (30 g). All herbs were soaked for 1 h and decocted
twice with an 8-fold volume of distilled water for 2 h. The
decoction was filtered and centrifuged twice at 12,000 rpm for 30
min to remove insoluble ingredients. The supernatants were mixed
with an equal volume of ethanol, maintained at 4°C overnight and
centrifuged at 12,000 rpm for 30 min to remove insoluble
ingredients. The resultant supernatants were lyophilized, weighed,
dissolved in RPMI-1640 medium and adjusted to a concentration of
400 mg/ml, and were sequentially passed through 0.45 and 0.22
μm sterilization filters.
Cell culture
The Bel-7402 human hepatocarcinoma cell line and
HL-7702 human hepatocyte cells were obtained from the Cell Bank of
Type Culture Collection of the Chinese Academy of Sciences.
Bel-7402 and HL-7702 cells were grown in RPMI-1640 medium with 10%
FBS and 1% Pen-Strep, and maintained at 37°C in a humidified
incubator with a 5% CO2 atmosphere.
Cell growth inhibition assay
Cells in the logarithmic growth phase were seeded
into 96-well plates (4×103 cells/well) and allowed to
attach for 24 h prior to treatment. The cells were exposed to
various doses of LYTF for 72 h, and cell viability was evaluated
every 24 h using the CCK-8 colorimetric assay, according to the
manufacturer’s instructions. The cell survival rate was calculated
as follows: cell survival rate (%) = experimental optical density
(OD) value/control OD value × 100%.
Detection of apoptosis
Bel-7402 cells were treated with various doses of
LYTF for 72 h, stained with 5 μg/ml Hoechst 33258 in the
dark for 5 min and assessed under fluorescence microscopy. For flow
cytometric analysis, LYTF-treated Bel-7402 cells were collected,
stained with Annexin V-FITC and PI as recommended by the
manufacturer, and detected in a FACScalibour flow cytometer
(Becton-Dickinson).
Caspase activity assay
Following treatment with varying concentrations of
LYTF, caspase-3, -8 and -9 activities were measured by the cleavage
of the specific chromogenic substrate according to the
manufacturer’s instructions. For caspase inhibition, cells
pre-treated with Z-VAD-FMK (50 μmol/l, 2 h) were incubated
with LYTF for a further 72 h.
Senescence-activated β-galactosidase
staining (SA-β-gal)
Bel-7402 cells (3×104) were plated in
35-mm diameter plates and treated with 200 μg/ml of LYTF for
5 days. SA-β-gal staining was performed according to the
manufacturer’s instructions (Cell Signaling Technology).
Western blot analysis
Western blot analysis was performed as described
previously (24). Briefly,
collected cells were lysed and subjected to 8–12% sodium dodecyl
sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and
transferred onto a nitrocellulose membrane (Amersham Biosciences,
Buckinghamshire, UK). The transferred membrane was blocked with 5%
nonfat milk, washed and probed with the indicated antibodies. Blots
were then washed and incubated with IRDye 700- and IRDye
800-conjugated secondary antibodies (Rockland Immunochemicals,
Gilbertsville, PA, USA), and visualized in Odyssey Infrared Imaging
System (LI-COR Biosciences, Lincoln, NE, USA).
Statistical analysis
Results are expressed as the means ± standard
deviation of at least two independent experiments, each conducted
in triplicate. Differences between the control and LYTF-treated
cells were analysed by one-way ANOVA. Differences were considered
significant at P<0.05.
Results
LYTF inhibits the proliferation of
Bel-7402 cells
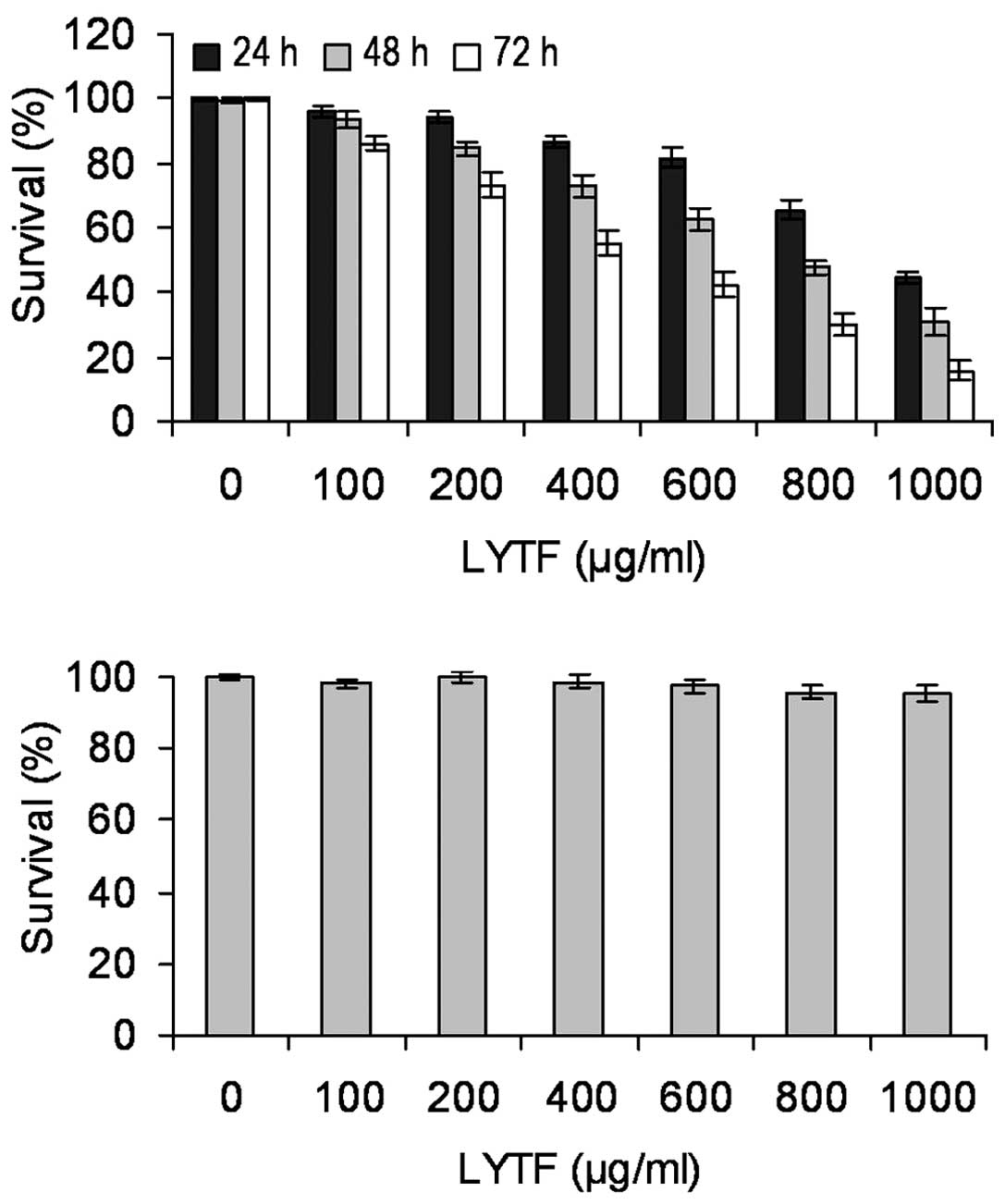
To determine the effects of LYTF on the growth of
Bel-7402 cells, the CCK-8 assay was performed. At a final
concentration of 100–1,000 μg/ml, LYTF inhibited the
proliferation of Bel-7402 cells in a dose- and time-dependent
manner (Fig. 1A) (P<0.05). By
contrast, LYTF had no significant effect on the proliferation of
human normal hepatocyte HL-7702 cells at these concentrations
(Fig. 1B).
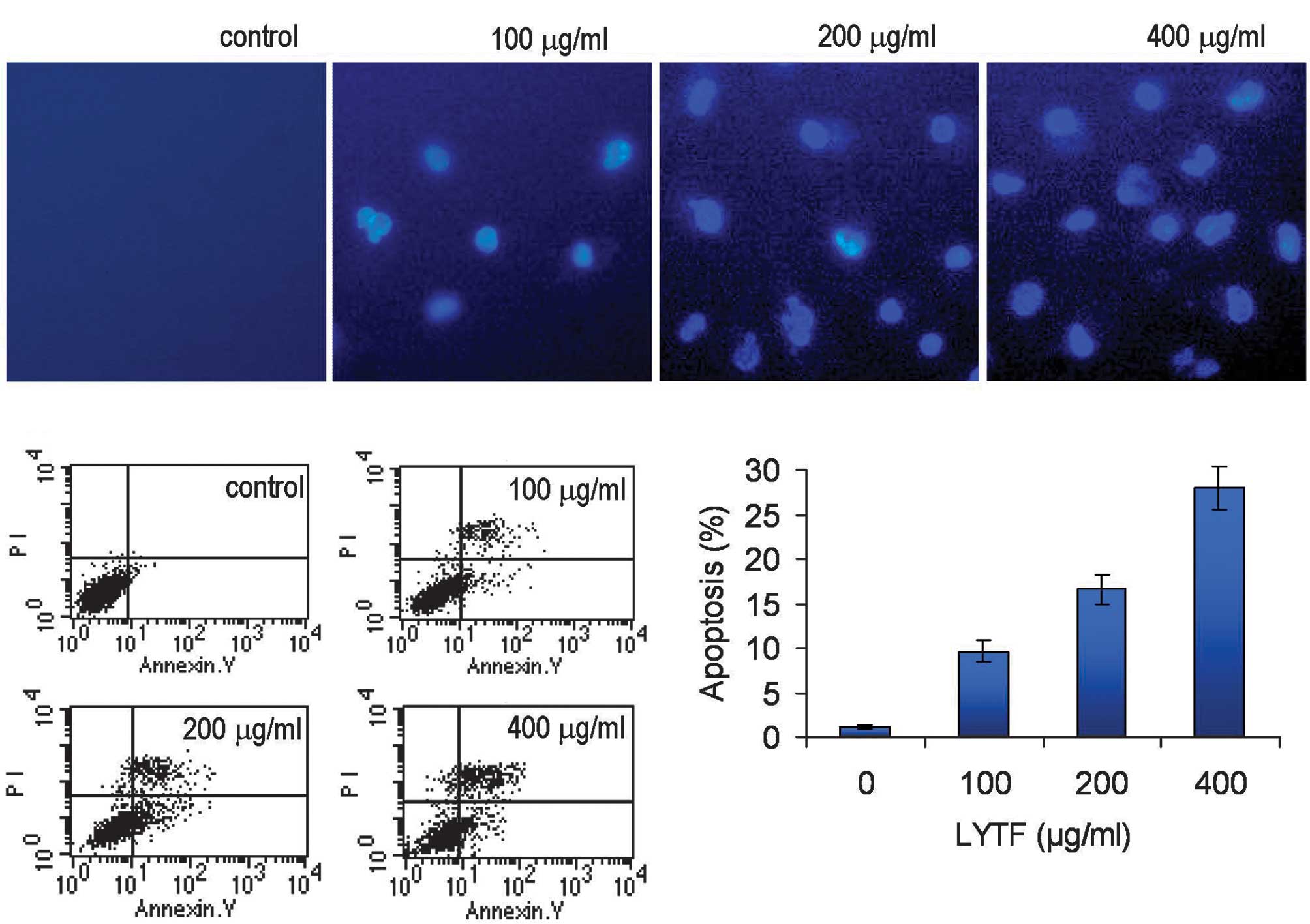
LYTF induces apoptosis of Bel-7402
cells
Following 72 h of incubation with 100–400
μg/ml of LYTF, Bel-7402 cells were stained with Hoechst
33258. We observed dense blue fluorescent particles within the
nucleus or cytoplasm in addition to significant nuclear
morphological changes, indicating the occurrence of apoptotic
processes (Fig. 2A). Flow
cytometric analysis further confirmed that incubation with 100–400
μg/ml of LYTF for 72 h induced considerable apoptosis of
Bel-7402 cells, compared to the control group (Fig. 2B and C) (P<0.01).
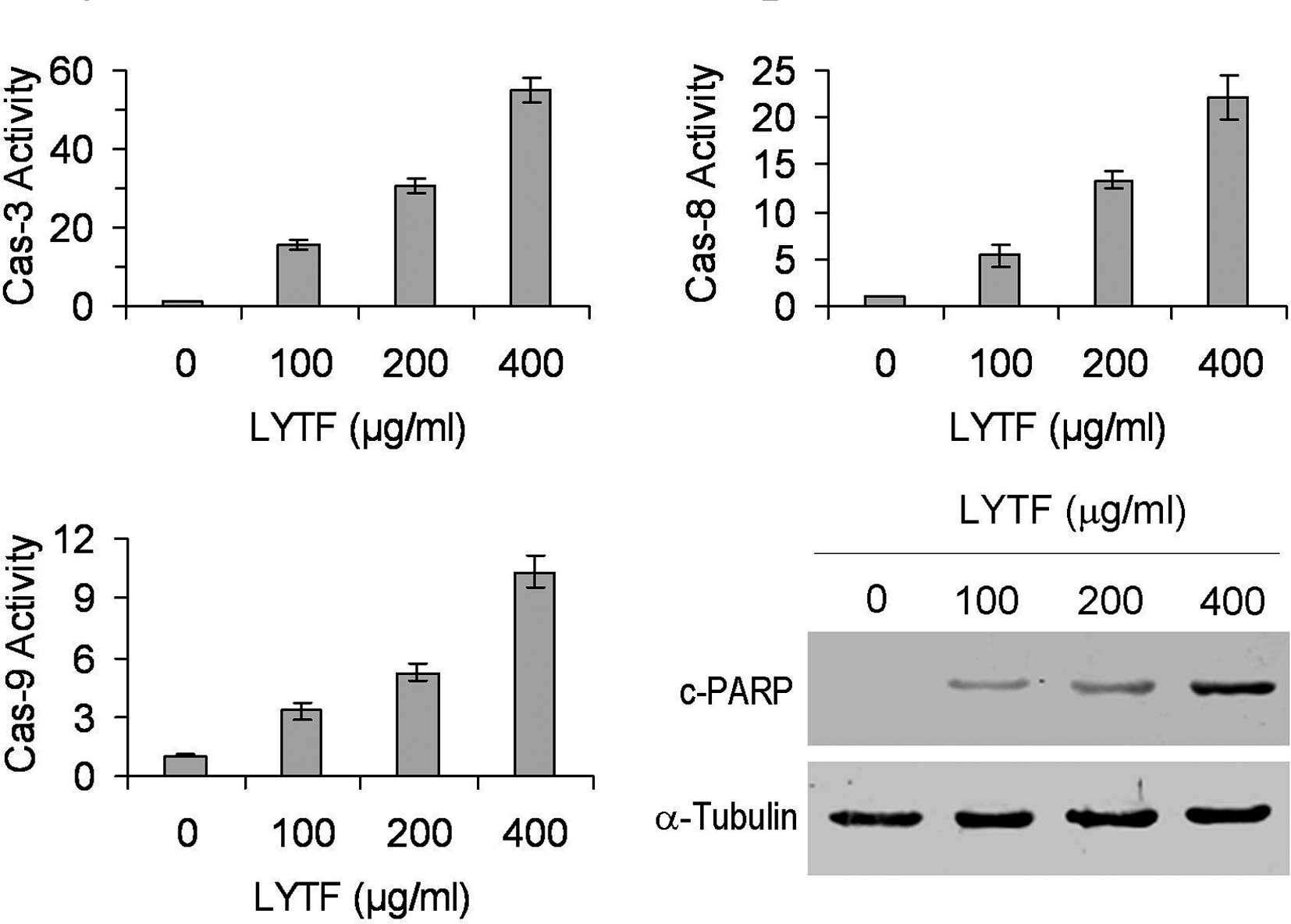
Role of caspases in LYTF-induced
anticancer effects
To determine whether caspases attributed to the
LYTF-induced apoptosis in Bel-7402 cells, the activities of
caspase-3, -8 and -9 were measured by the cleavage of the specific
substrate. Caspase activity assays revealed that LYTF was capable
of activating caspases-3, -8 and -9 in Bel-7402 cells to varying
degrees in a dose-dependent manner (Fig. 3A–C) (P<0.01). In addition, PARP,
one of the earliest substrates of caspase-3 during apoptosis
(26), was also cleaved following
LYTF treatment (Fig. 3D).
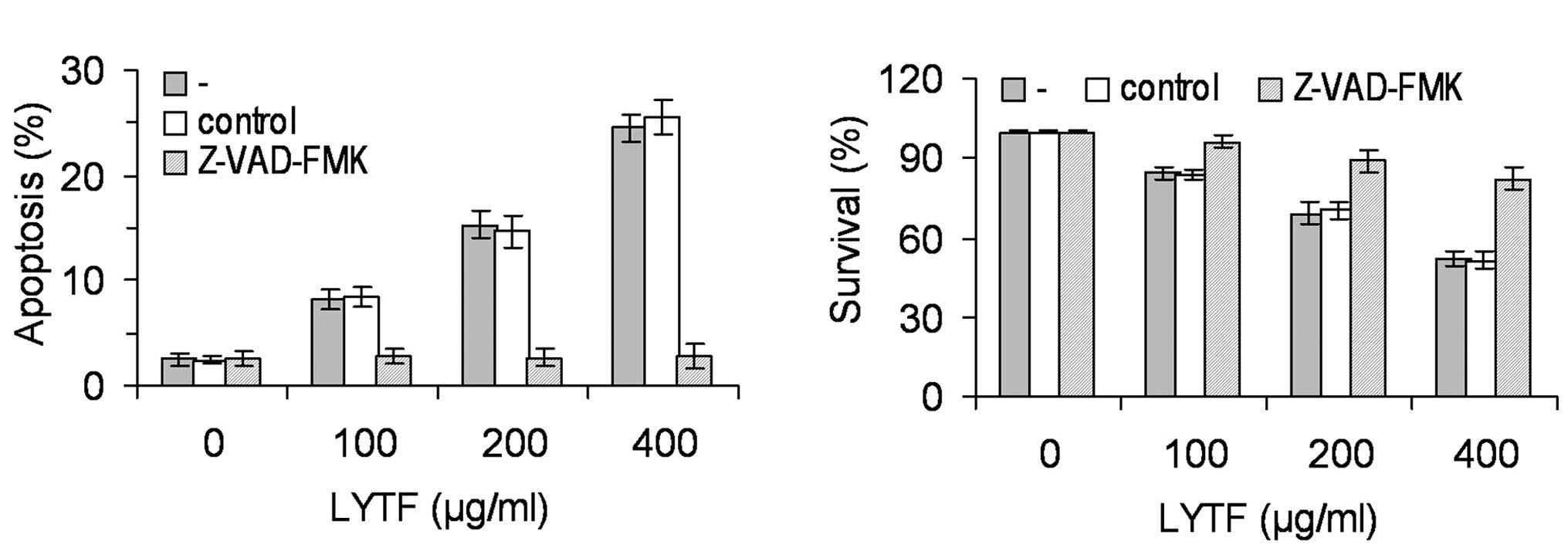
Furthermore, the LYTF-induced apoptosis of Bel-7402 cells was
completely abrogated by the pan-caspase inhibitor, Z-VAD-FMK
(Fig. 4A) (P<0.01), suggesting
that the LYTF-induced apoptosis was associated with caspase
signaling cascades. To our surprise, the inhibitory effects of LYTF
on the proliferation of Bel-7402 cells were partially abrogated by
the Z-VAD-FMK inhibitor (Fig. 4B)
(P<0.01), suggesting that there may be another mechanism
contributing to the LYTF-induced anticancer effects.
LYTF induces cell senescence in Bel-7402
cells
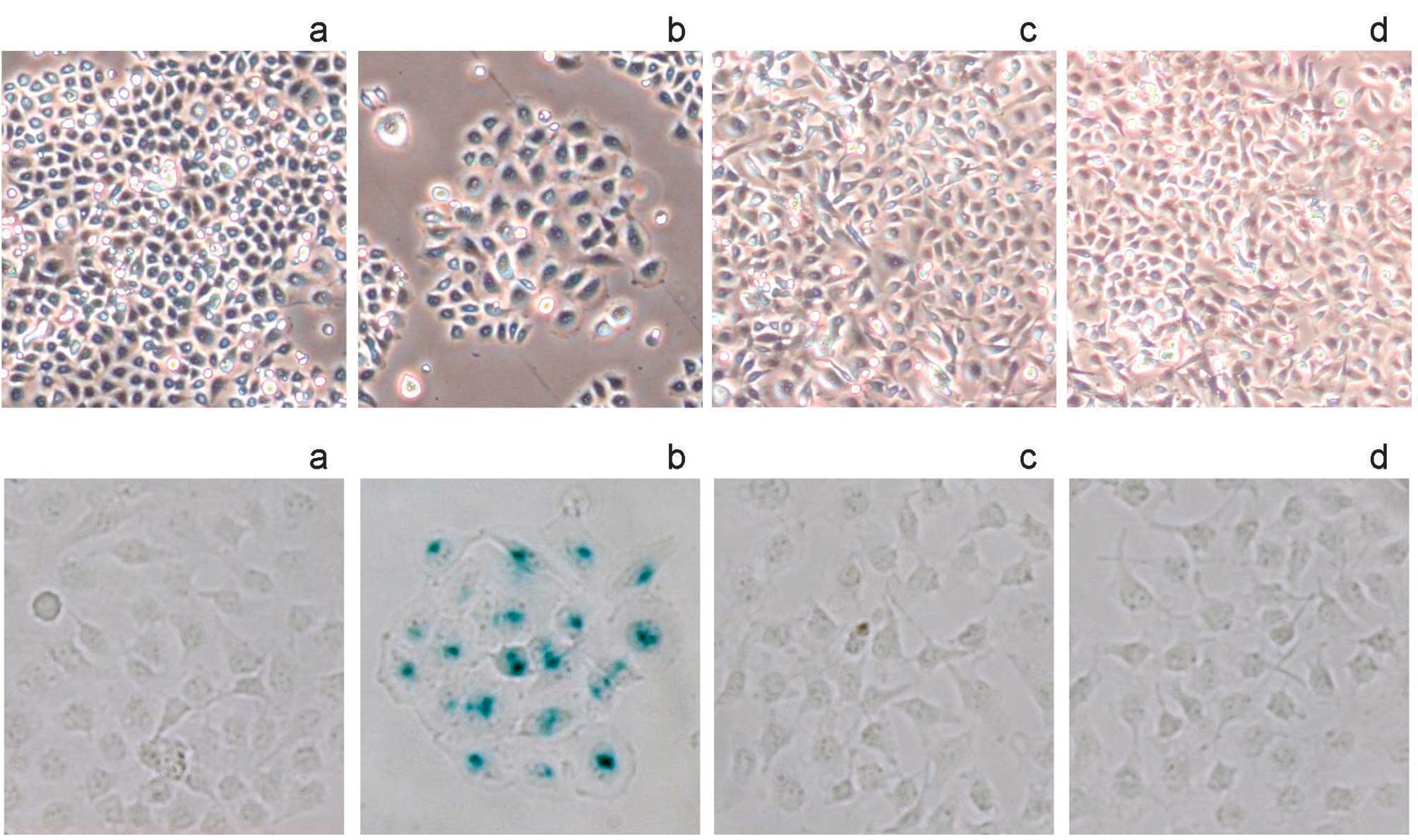
With low- dose LYTF treatment, partial Bel-7402
cells exhibited a large and flattened morphology, reminiscent of
cell senescence (Fig. 5A-b).
Therefore, SA-β-gal staining was performed. As shown in Fig. 5B-a and -b, LYTF treatment resulted
in SA-β-gal-positive staining, compared to the controls. By
contrast, neither morphology nor SA-β-gal activity was affected by
LYTF treatment in normal HL-7702 hepatocytes (Fig. 5B-c and -d). In addition, flow
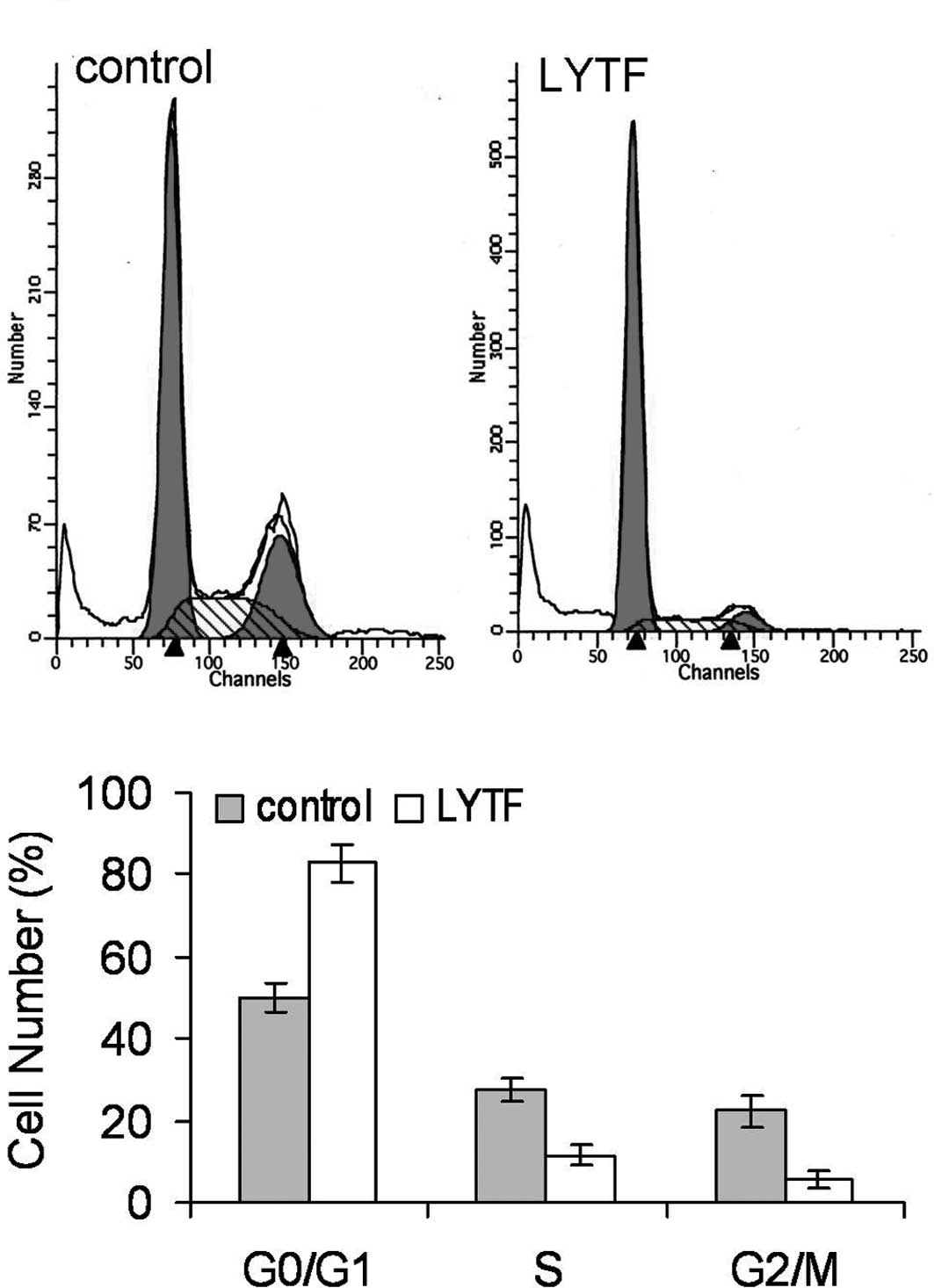
cytometric analysis revealed that the cell cycle of LYTF-treated
Bel-7402 cells was arrested in the G0/G1 phase (Fig. 6) (P<0.01). These observations
suggest that LYTF may induce senescence in Bel-7402 cells.
Effects of LYTF on the expression of
senescence-associated genes
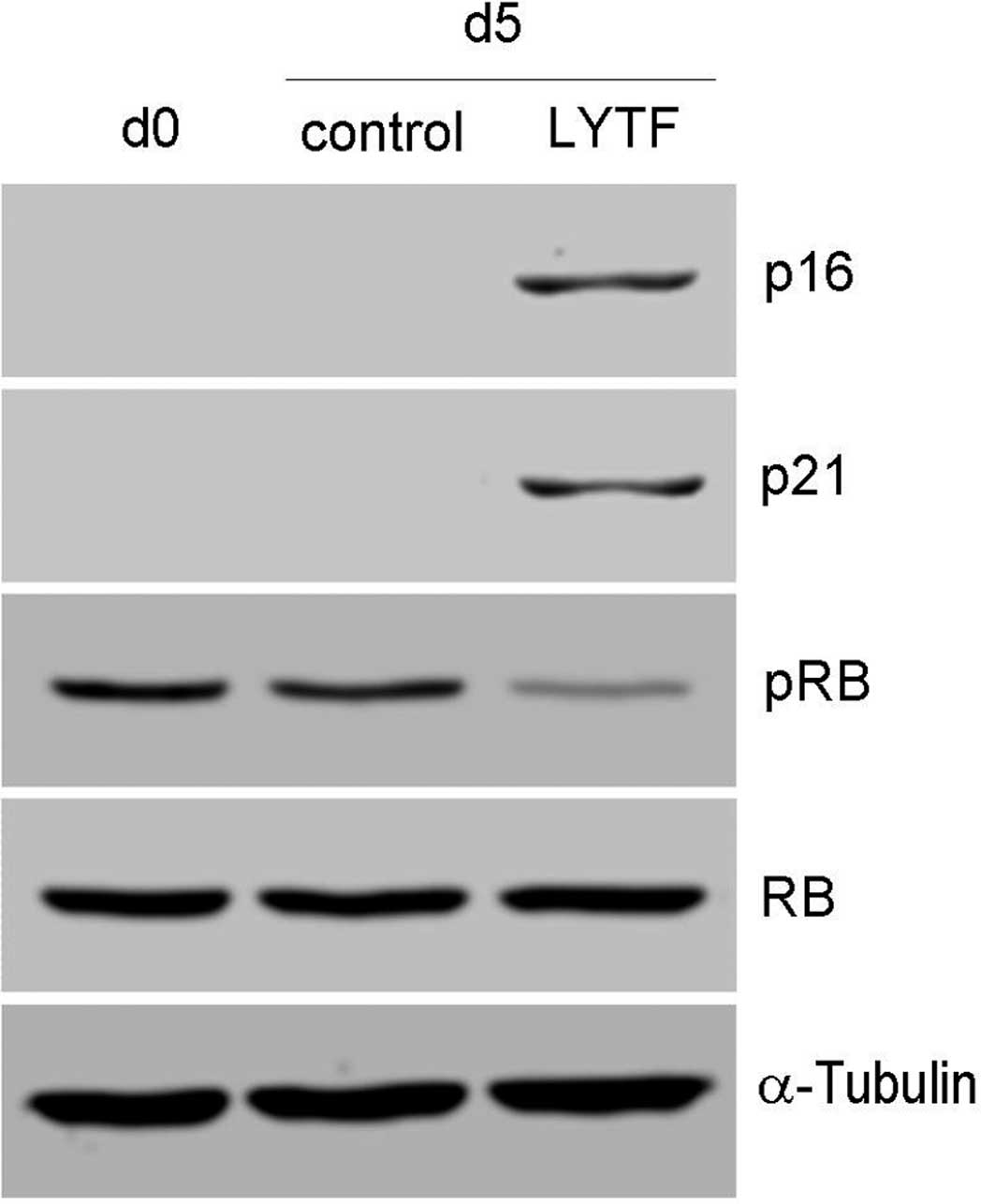
The effects of LYTF on the expression of
senescence-associated genes in Bel-7402 cells were detected by
Western blot analysis. As shown in Fig. 7, treatment with LYTF caused an
up-regulation in the expression of p21 and p16. Expression levels
of Rb did not change, but Rb phosphorylation was inhibited by LYTF
treatment. These results suggest that the LYTF-induced cell
senescence of Bel-7402 cells is related to elevated p21 and p16
expression and inhibition of Rb phosphorylation.
Discussion
Current cancer pharmacotherapy strategies, such as
chemotherapy and targeted therapy, act on cancer cells mainly
through apoptosis, cell senescence, autophagy and other mechanisms
to achieve therapeutic effects (27,28).
Apoptosis is an evolutionarily conserved cell suicide process
elicited by physiological, pharmacological and pathological
conditions. The initiation and execution of apoptosis depends on
the activation of the extrinsic and/or intrinsic death pathways
(28,29). Extrinsic or death receptor pathways
are associated with death receptors, such as FAS, the TRAIL
receptor (TRAIL-R) and TNF receptor (TNF-R). On binding to the
corresponding ligands, such as FasL, TRAIL or TNFα, death receptors
oligomerize and their death domains undergo conformational changes
to facilitate the interaction with the adaptor protein,
Fas-associated death domain (FADD), resulting in the recruitment of
caspase-8 to form the death-inducing signaling complex. Activated
caspase-8 then activates caspase-3, triggering apoptosis.
Conversely, intrinsic or mitochondrial pathways involve signals to
the mitochondria that lead to the release of cytochrome c and
Apaf-1, forming an apoptosome that activates the initiating
protease caspase-9, which, in turn, activates caspase-3, causing
the cell to undergo apoptosis. The activation of caspases-8 and -9
are the hallmark events of extrinsic and intrinsic apoptotic
pathways, respectively. In the present study, following LYTF
treatment, Bel-7402 cells exhibited apoptotic morphology. The
apoptotic cells were detected by flow cytometry. Furthermore, LYTF
activates caspases-3, -8 and -9, and the LYTF-induced apoptosis may
be blocked by a caspase inhibitor. These results indicate that the
LYTF-induced apoptosis of Bel-7402 cells is closely related to
caspase activation.
Cell senescence is a state of stable, irreversible
cell cycle arrest provoked by a variety of stimuli. Senescent cells
maintain some metabolic activity, but can no longer proliferate,
even if stimulated with mitogens. Cell senescence is usually
characterized by a large and flattened morphology, an increase in
intracellular granules and elevated SA-β-gal activity (30). Similar to apoptosis, cell
senescence plays a crucial role in suppressing tumorigenesis and
may contribute to the outcome of cancer therapy (29–32).
It has been shown that chemotherapeutic agents, such as cisplatin,
doxorubicin, SN-38 and camptothecin, inhibit the growth of tumors
via cell senescence (32–35). Cell senescence as a tumor
suppression mechanism has increasingly attracted attention in
recent times (36). We observed
that LYTF treatment caused Bel-7402 cells to assume a large and
flat morphology, test positive for SA-β-gal staining and become
arrested in the G0/G1 phase of the cell cycle, suggesting that LYTF
treatment induces senescence in these cells.
The occurrence of cell senescence is closely related
to the activation of the CDKN1a (p21WAF-1/Cip1)/pRB/E2F or CDKN2a
(p16INK4A)/pRB/E2F signaling pathway (37,38).
p21, a crucial cell cycle regulator, inhibits a variety of
cyclin/CDK complexes and induces the hypophosphorylation or
dephosphorylation of protein Rb (pRb). Hypophosphorylated pRb binds
to E2F and prevents it from activating target genes that are
essential in the cell cycle, usually leading to cell cycle arrest.
p21 plays a crucial role in cancer cell senescence, dependent or
independent of p53 (31,35,39).
The CDK4 and CDK6 inhibitor, p16, also participates in the
regulation of Rb phosphorylation, induces cell cycle arrest and may
contribute to the induction of cell senescence (25,34,40,41).
An increase in p16 and p21 expression and the inhibition of Rb
phosphorylation was observed in Bel-7402 cells following LYTF
treatment, suggesting that the LYTF-induced cell senescence of
Bel-7402 is associated with these processes. Moreover, the
inhibition of Rb phosphorylation by LYTF may be due to the
collaboration of p21 and p16.
In conclusion, our study demonstrates that LYTF
inhibits proliferation and activates caspase cascades to induce
apoptosis in Bel-7402 hepatocarcinoma cells. Furthermore, LYTF also
induces senescence in Bel-7402 cells, which may correlate with the
increased expression of p16 and p21, and the inhibition of Rb
phosphorylation. The present study provides new insights into
traditional Chinese medicine approaches to liver cancer treatment
that are worth pursuing.
Acknowledgements
This study was partially supported by
the Key Basic Research Program from the Science and Technology
Commission of Shanghai Municipality (09JC1413600), the Program for
Long-Yi Scholars and Research Team in State Clinical Research
Center of TCM (LYTD-04), the Key Discipline of State Administration
of Traditional Chinese Medicine (Traditional Chinese Medicine in
Oncology) and the Shanghai Shen Kang Platform Grant (SHDC12007206).
The authors would also like to thank the Shanghai Key Laboratory of
Tissue Engineering for providing technical assistance.
References
|
1.
|
A JemalF BrayMM CenterJ FerlayE WardD
FormanGlobal cancer statisticsCA Cancer J
Clin616990201110.3322/caac.20107
|
|
2.
|
K HagymásiZ TulassayEpidemiology risk
factors and molecular pathogenesis of primary liver cancerOrv
Hetil149541548200818343770
|
|
3.
|
B RamponeB SchiavoneG ConfuortoCurrent
management of hepatocellular cancerCurr Oncol
Rep12186192201010.1007/s11912-010-0094-3
|
|
4.
|
R LordA SuddlePJ RossEmerging strategies
in the treatment of advanced hepatocellular carcinoma: the role of
targeted therapiesInt J Clin
Pract65182188201110.1111/j.1742-1241.2010.02545.x21235699
|
|
5.
|
RS OliveriJ WetterslevC GluudTransarterial
(chemo) embolisation for unresectable hepatocellular
carcinomaCochrane Database Syst Rev16CD004787201121412886
|
|
6.
|
PJ WysockiTargeted therapy of
hepatocellular cancerExpert Opin Investig
Drugs19265274201010.1517/1354378090351411020074016
|
|
7.
|
Y YuQ LangZ ChenB LiC YuD ZhuX ZhaiC
LingThe efficacy for unresectable hepatocellular carcinoma may be
improved by transcatheter arterial chemoembolization in combination
with a traditional Chinese herbal medicine formula: a retrospective
studyCancer11551325138200910.1002/cncr.24567
|
|
8.
|
MB MengYL CuiYS GuanZ YingMH ZhengCK
YuanRM ZhangTraditional Chinese medicine plus transcatheter
arterial chemoembolization for unresectable hepatocellular
carcinomaJ Altern Complement
Med1410271042200810.1089/acm.2008.006018990050
|
|
9.
|
X ShuM McCullochH XiaoM BroffmanJ
GaoChinese herbal medicine and chemotherapy in the treatment of
hepatocellular carcinoma: a meta-analysis of randomized controlled
trialsIntegr Cancer
Ther4219229200510.1177/153473540527992716113029
|
|
10.
|
Q DuB HuKP ShenHM AnProgress in TCM
pathogenesis and treatment of liver cancerWorld J Integr Tradit
West Med58148172010
|
|
11.
|
FG HouCQ LingG ZhaoXM HeInvestigation of
clinical distribution of basic TCM Syndrome in primary hepatic
carcinomaShanghai J Tradit Chin Med3922232005
|
|
12.
|
YF WuSP WangJM SunJN WeiClinical
distribution and standards of TCM Syndrome of primary hepatic
carcinomaJ Shanxi Coll Tradit Chin Med821232007
|
|
13.
|
MQ PanPH ZengB PanRegularity of
traditional Chinese medicine in middle and advanced primary hepatic
carcinomaChinese Arch Tradit Chin Med21164116422003
|
|
14.
|
Q LiuYB ZhangCH MaXQ YueCQ LingAnalysis of
literature on therapeutic methods and medicines of traditional
Chinese medicine for primary liver cancerJ Chin Integr
Med3260262200510.3736/jcim2005040316009099
|
|
15.
|
M XiangZL GuWX ZhouCY GuoAnti-cancer
effects of extract of glossy privet fruit in vivoJiangsu Pharm Clin
Res1013152002
|
|
16.
|
L LiRL QiuG ChengY ShiY HanY GaoY LuThe
study on the anti-tumor effect of polysaccharides from glossy
privet fruit in vitro and in vivoChin Pharm Bull24161916222008
|
|
17.
|
Q DuB HuKP ShenA Survey of anti-cancer
effects of Chinese tonifying herbs in primary hepatic carcinomaJ
Chin Med Mater33151215162010
|
|
18.
|
SL YanCY HuangST WuMC YinOleanolic acid
and ursolic acid induce apoptosis in four human liver cancer cell
linesToxicol In
Vitro24842848201010.1016/j.tiv.2009.12.00820005942
|
|
19.
|
JM YangBB YangA ZhangJQ WangB ZhangThe
research and development of Polygounm cuspidatumActa Agric
Boreali-occidentalis Sinica131561592004
|
|
20.
|
L FengLF ZhangT YanJ JinWY TaoStudies on
active substance of anticancer effect in Polygonum
cuspidatumJ Chin Med Mater29689691200617059010
|
|
21.
|
D DelmasE SolaryN LatruffeResveratrol, a
phytochemical inducer of multiple cell death pathways: apoptosis,
autophagy and mitotic catastropheCurr Med
Chem1811001121201110.2174/09298671179502970821291372
|
|
22.
|
Z GaoMS XuTL BarnettCW XuResveratrol
induces cellular senescence with attenuated mono-ubiquitination of
histone H2B in glioma cellsBiochem Biophys Res
Commun407271276201110.1016/j.bbrc.2011.02.00821481687
|
|
23.
|
YL HsuPL KuoTF TzengSC SungMH YenLT LinCC
LinHuang-lian-jie-du-tang, a traditional Chinese medicine
prescription, induces cell-cycle arrest and apoptosis in human
liver cancer cells in vitro and in vivoJ Gastroenterol
Hepatol23e290e299200810.1111/j.1440-1746.2008.05390.x18522681
|
|
24.
|
B HuKP ShenHM AnY WuQ DuAqueous extract of
curcuma aromatica induces apoptosis and G2/M arrest in human colon
carcinoma LS-174-T cells independent of p53Cancer Biother
Radiopharm2697104201110.1089/cbr.2010.085321348775
|
|
25.
|
B HuHM AnKP ShenQ DuSenescence-inducing
effects of Chinese herbal medicine Tenglong Buzhong Decoction on
human colon carcinoma LS-174-T cells and the mechanismJ Chin Integr
Med810481052201010.3736/jcim2010110821078269
|
|
26.
|
YA LazebnikSH KaufmannS DesnoyersGG
PoirierWC EarnshawCleavage of poly(ADP-ribose)polymerase by a
proteinase with properties like
ICENature371346347199410.1038/371346a08090205
|
|
27.
|
JM BrownLD AttardiThe role of apoptosis in
cancer development and treatment responseNat Rev
Cancer5231237200515738985
|
|
28.
|
P MeierKH VousdenLucifer’s labyrinth - ten
years of path finding in cell deathMol Cell287467542007
|
|
29.
|
MV ChiantoreS VannucchiG ManginoZA
PercarioE AffabrisG FiorucciG RomeoSenescence and cell death
pathways and their role in cancer therapeutic outcomeCurr Med
Chem16287300200910.2174/09298670978700269119149578
|
|
30.
|
CA SchmittCellular senescence and cancer
treatmentBiochim Biophys Acta1775520200717027159
|
|
31.
|
JA EwaldJA DesotelleG WildingDF
JarrardTherapy-induced senescence in cancerJ Natl Cancer
Inst10215361546201010.1093/jnci/djq36420858887
|
|
32.
|
DA GewirtzSE HoltLW ElmoreAccelerated
senescence: an emerging role in tumor cell response to chemotherapy
and radiationBiochem
Pharmacol76947957200810.1016/j.bcp.2008.06.02418657518
|
|
33.
|
X DiRP ShiuIF NewshamDA GewirtzApoptosis,
autophagy, accelerated senescence and reactive oxygen in the
response of human breast tumor cells to adriamycinBiochem
Pharmacol7711391150200910.1016/j.bcp.2008.12.01619185564
|
|
34.
|
RH Te PoeleAL OkorokovL JardineJ
CummingsSP JoelDNA damage is able to induce senescence in tumor
cells in vitro and in vivoCancer Res6218761883200211912168
|
|
35.
|
Z HanW WeiS DunawayJW DarnowskiP
CalabresiJ SedivyEA HendricksonKV BalanP PantazisJH WycheRole of
p21 in apoptosis and senescence of human colon cancer cells treated
with camptothecinJ Biol
Chem2771715417160200210.1074/jbc.M11240120011877436
|
|
36.
|
C NardellaJG ClohessyA AlimontiPP
PandolfiPro-senescence therapy for cancer treatmentNat Rev
Cancer11503511201110.1038/nrc305721701512
|
|
37.
|
IB RoninsonTumor cell senescence in cancer
treatmentCancer Res6327052715200312782571
|
|
38.
|
GP DimriWhat has senescence got to do with
cancerCancer Cell7505512200510.1016/j.ccr.2005.05.02515950900
|
|
39.
|
D LodyginA MenssenH HermekingInduction of
the Cdk inhibitor p21 by LY83583 inhibits tumor cell proliferation
in a p53-independent mannerJ Clin
Invest11017171727200210.1172/JCI021658812464677
|
|
40.
|
UM ChangCH LiLI LinCP HuangLS KanSB
LinGanoderiol F, a ganoderma triterpene, induces senescence in
hepatoma HepG2 cellsLife
Sci7911291139200610.1016/j.lfs.2006.03.02716635496
|
|
41.
|
R XingW LiJ CuiJ ZhangB KangY WangZ WangS
LiuY LuGastrokine 1 induces senescence through p16/Rb pathway
activation in gastric cancer cellsGutJune142011(E-pub ahead of
print).
|