Introduction
Stroke is a leading cause of mortality and
disability worldwide and 70–80% of all cases are ischemic stroke
(1). Post-ischemic inflammation
is an essential step in the progression of ischemic stroke
(2). Microglial cells constitute
the resident immune cell population of the mammalian central
nervous system (CNS), which play a pivotal role in
neuroinflammation by avidly surveying the brain parenchyma
(3,4). In this sense, microglial cells
become activated and migrate to the injury site in order to fully
develop a concerted immune response, involving the release of both
trophic and pro-inflammatory factors, such as tumor necrosis
factor-α (TNF-α), interleukin (IL)-6 and IL-1β and nitric oxide
(NO), which are believed to lead to severe neuronal death and brain
injury (5,6). Chemokines [monocyte chemoattractant
protein-1 (MCP-1), IL-8 and macrophage inflammatory protein-1α
(MIP-1α)] produced by microglial cells are also important enhancers
of post-ischemic inflammation (7,8).
Gua Lou Gui Zhi decoction (GLGZD) is a classical traditional
Chinese formula that was first prescribed in the Eastern Han
dynasty, around 210 AD and it consists of a combination of six
herbs, including Trichosanthis Radix, Ramulus Cinnamomi, Paeonia
lactiflora, Glycyrrhiza, Zingiber officinale Roscoe and
Fructus Jujubae. It has long been used in China in clinical
practice for the treatment of post-stroke disabilities, such as
muscular spasticity (9). We have
previously demonstrated that GLGZD exerts inhibitory effects on
LPS-induced inflammatory events in microglial cells (9). The anti-inflammatory effects of
GLGZD prompted us to examine the effects of GLGZD on microglial
migration. The signal transduction pathways in BV-2 microglial
cells induced by lipopolysaccharide (LPS), a bacterial endotoxin
widely used for studying experimental inflammation, may eventually
lead to the phosphorylation of the mitogen-activated protein kinase
(MAPK) signaling pathways, resulting in the production of
inflammatory mediators, including chemotactic cytokines. In the
present study, we attempted to evaluate the effects of GLGZD on
LPS-induced microglial motility and the regulation of chemotactic
cytokines by GLGZD in activated BV-2 microglial cells. We
demonstrate that GLGZD ultimately protects neurons from secondary
damage following ischemic stroke.
Materials and methods
Materials and reagents
LPS (from Escherichia coli 055:B5) and
3-[4,5-dimethyl-2-thiazolyl]-2,5-diphenyltetrazoliumbromide (MTT)
were obtained from Sigma-Aldrich (St. Louis, MO, USA). FBS, DMEM,
penicillin, streptomycin, 0.05% (w/v) trypsin/EDTA and
phosphate-buffered saline (PBS) were obtained from Gibco-BRL
(Gaithersburg, MD, USA). The cytokine (MCP-1, MIP-1α and IL-8)
ELISA kits were purchased from R&D Systems (Minneapolis, MN,
USA). The real-time PCR kit was purchased from (Takara, Shiga,
Japan). The antibodies used for western blot analysis in this study
included: mouse p38 monoclonal antibody (mAb), phospho-p38 mAb,
c-Jun N-terminal protein kinase (JNK) mAb, phospho-JNK mAb,
extracellular signal-regulated kinase 1/2 (ERK1/2) mAb,
phospho-ERK1/2 mAb and β-actin mAb (Santa Cruz Biotechnology, Inc.,
Santa Cruz, CA, USA) and horseradish peroxidase (HRP)-conjugated
secondary antibodies were from Cell Signaling Technology, Inc.
(Beverly, MA, USA).
Preparation of water extract of
GLGZD
Medicinal plants were supplied by the Guo Yi Tang
Chinese Herbal medicine store (Fuzhou, China). The preparation of
the GLGZD water extract was performed as described in our previous
study (9). The stock and working
concentrations of GLGZD were prepared by dissolving the extract in
culture medium to a concentration of 50 mg/ml and 100 μg/ml.
Cell line culture
BV-2 murine microglial cells were maintained in DMEM
supplemented with 10% fetal bovine serum, 100 U/ml penicillin and
100 μg/ml streptomycin at 37°C in a humidified incubator under 5%
CO2. Cells were plated at a density of 1×105
cells per well in 96-well trays (MTT assays), 8×104
cells per well in 24-well trays (ELISA) or plated at a density of
5×105 cells per well in 6-well trays (for the remaining
experiments). Cell treatments included the following incubation
setup: serum-free medium for 4 h prior to cell treatment, LPS (100
ng/ml, Sigma) and/or GLGZD water extract (100 μg/ml) for 12 h (mRNA
expression evaluation, cytokine release and migration analyses) or
24 h (cytotoxicity assays).
MTT assay
BV-2 microglial cell viability was assessed by MTT
assay. MTT is actively catalyzed by mitochondrial succinate
dehydrogenase to form dark blue formazan in live cells. The
formation of formazan is therefore used as an indicator of cell
viability. BV-2 cells cultured in 96-well plates were treated with
GLGZD at various concentrations and LPS for 24 h, followed by
incubation with MTT (0.5 mg/ml) for an additional 4 h. The formazan
formed in the cells was dissolved by dimethyl sulfoxide (DMSO;
Sigma-Aldrich) and read at 570 nm using a microplate absorbance
reader (EXL 808; BioTek Instruments, Bad Friedrichshall,
Baden-Wurttemberg, Germany).
Scratch wound assay
The BV-2 cells were seeded in 6-well plates in
monolayer until approximately 95% confluent prior to the migration
assay. Four hours before performing the sratch wound assay, the
medium was replaced with serum-free medium to ensure that no
proliferation occurred during the experiments. The wound was made
by a perpendicular scratch made with a pipette tip. The cells were
stimulated with 100 ng/ml LPS, 100 μg/ml GLGZD or 100 ng/ml LPS +
100 μg/ml GLGZD for 12 h. Following treatment, microphotographs of
the cells that had migrated into the open wound were taken.
Transwell migration assay
In this study, the Costar Transwell System (8-μm
pore size polycarbonate membrane, Costar, Cambridge, MA, USA) was
used to evaluate vertical cell migration. BV-2 cells in 0.2 ml
serum-free medium were seeded in the upper well and 0.6 ml 100
ng/ml LPS, 100 μg/ml GLGZD, 100 ng/ml LPS + 100 μg/ml GLGZD, or
DMEM as the negative control were added to the lower chamber. At
the end of a 24-h incubation period at 37°C, the cells on the upper
side of the filters were removed with cotton-tipped swabs and the
filters were fixed with 4% paraformaldehyde in PBS for 30 min and
stained with crystal violet in PBS for 15 min followed by obtaining
representative images using a microscope. The migrated cells on the
lower surface of the membrane were then solubilized in extraction
buffer. Optical density (OD) values at 540 nm were determined using
a microplate reader.
Enzyme-linked immunosorbent assay
(ELISA)
The BV-2 cells were plated in a 24-well cell culture
plate and incubated with GLGZD water extract (100 μg/ml) in the
presence or absence of LPS (100 ng/ml) for 12 h. Following
treatment with the stimuli, ELISA kits were used according to the
manufacturer’s instructions for mouse MCP-1, MIP-1α and IL-8. The
OD values were recorded at 450 nm.
Quantitative polymerase chain
reaction
For gene expression analysis, the medium was removed
following incubation and the BV-2 cells were washed with PBS twice.
Total RNA was isolated from the BV-2 murine microglial cell
cultures using TRIzol reagent (Invitrogen Corp., Carlsbad, CA,
USA). Precipitated RNA was resuspended in 20 μl of
diethylpyrocarbonate (DEPC)-treated water and quantified by
measuring the OD using a NanoDrop 2000 spectrophotometer (Thermo
Scientific, Waltham, MA, USA). cDNA was synthesized using avian
myeloblastosis virus reverse transcriptase (Takara), according to
the manufacturer’s instructions. Quantitative PCR was performed in
triplicate using an Applied Biosystems Prism 7500 Fast Sequence
Detection System (Applied Biosystems, Foster City, CA, USA) with
the SYBR-Green I quantitative PCR kit according to the instructions
of the manufacturer (Takara). The sequences of the primers were as
follows: MCP-1 forward, 5′-TCGGAACCAAATGAGATCAGAAC-3′ and reverse,
5′-GAGGTGGTTGTGGAAAAGGTAGTG-3′ (135 bp); MIP-1α forward,
5′-TCGTTGACTATTTTG AAACCAGCAG-3′ and reverse, 5′-TCAGGCATTCAGTT
CCAGGTCA-3′ (140 bp); IL-8 forward, 5′-ATGCTCCTGCT GGCTGTCCT-3′ and
reverse, 5′-TTGGGACTGCTATCAC TTCCTTTC-3′ (178 bp); and β-actin
forward, 5′-GAGAGGG AAATCGTGCGTGACA-3′ and reverse, 5′-ACCCAAG
AAGGAAGGCTGGAAA-3′ (192 bp) (Takara). Levels of RNA expression were
determined using the 7500 Fast System (Applied Biosystems). The
results were normalized to β-actin expression and analyzed relative
to their untreated counterparts. The relative quantification of the
gene of interest was determined using the comparative CT method
(2−ΔΔCt).
Western blot analysis
The cells were washed twice with cold PBS and lysed
in ice-cold radioimmunoprecipitation buffer containing protease
inhibitors [phenylmethylsulfonyl fluoride (PMSF)]. The lysate was
centrifuged for 10 min at 12,000 × g at 4°C and the supernatant
collected. Total protein concentration was quantified using the
bicinchoninic acid (BCA) protein assay kit and the samples were
stored at −80°C. Subsequently, protein (50 μg) samples was
separated by SDS-PAGE on 10% gels and then transferred onto PVDF
membranes (Millipore, Billerica, MA, USA). The membranes were
blocked in Tris-buffer for 1 h at room temperature and then
incubated overnight at 4°C with primary antibodies: p38 and
phospho-p38 mAb, JNK and phospho-JNK mAb, ERK1/2 and phospho-ERK1/2
mAb and β-actin mAb (1:1,000). After rinsing with TBS-T, membranes
were incubated for 1 h at room temperature with horseradish
peroxidase-conjugated secondary antibodies: anti-goat, anti-rabbit
or anti-mouse IgG (1:1,000). Protein immunoreactive bands were
developed using the enhanced chemiluminescence (ECL)-Plus kit
(RPN2132; GE Healthcare, Pittsburgh, PA, USA) for 1 min and
chemiluminescent bands were detected on Kodak autoradiography film
in a dark room, and their densities were measured using ImageJ
software.
Data analysis
Data are presented as the means ± standard error of
the mean (SEM). Statistical analysis was performed using SPSS 15.0
software. One-way analysis was used for a comparison among the
experimental conditions and a P-value <0.05 was considered to
indicate a statistically significant difference.
Results
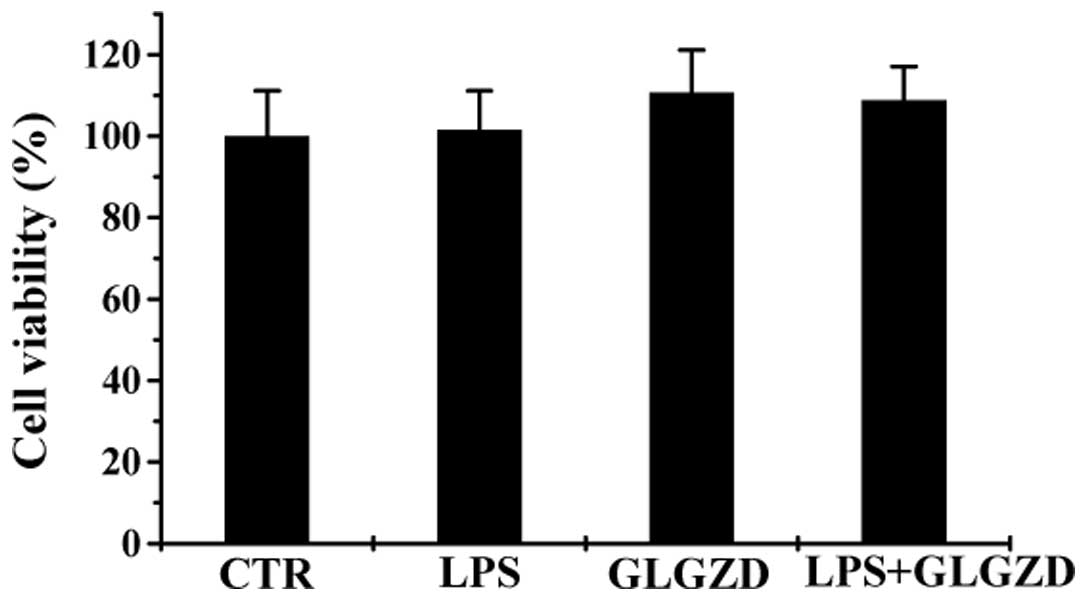
GLGZD does not affect BV-2 cell
viability
The cytotoxicity of GLGZD was evaluated in the
presence or absence of LPS by MTT assay. GLGZD at a concentration
of 100 μg/ml did not significantly affect the viability of the BV-2
cells when the cells were incubated with or without LPS (100 ng/ml)
for 24 h (Fig. 1). Hence, GLGZD
at 100 μg/ml alone and co-treatment with LPS exerted no cytotoxic
effects on the BV-2 microglial cells compared with the control
group.
GLGZD exerts an inhibitory effect on
LPS-induced microglial migration
To assess the effect of GLGZD on microglial
motility, we performed a scratch wound assay and Transwell
migration assay. As shown in Fig.
2A, representative microscopic images clearly illustrate that
the LPS-activated BV-2 cells exhibited an enhanced migratory
potential compared with the control; however, we observed that 100
μg/ml GLGZD significantly reduced cell motility in the co-culture
with LPS and GLGZD alone had no effect on microglial migration.
Similar to the scratch assays, the Transwell migration assay over a
period of 24 h indicated that the migratory capacity of the BV-2
cells was not altered by GLGZD alone; however, co-treatment with
LPS decreased the number of migrating cells (Fig. 2B). The statistical analysis of
three independent experiments revealed that GLGZD significantly
reduced the number of migrating cells in the presence of LPS in the
culture medium (Fig. 2C). In both
assays, GLGZD significantly attenuated microglial migration.
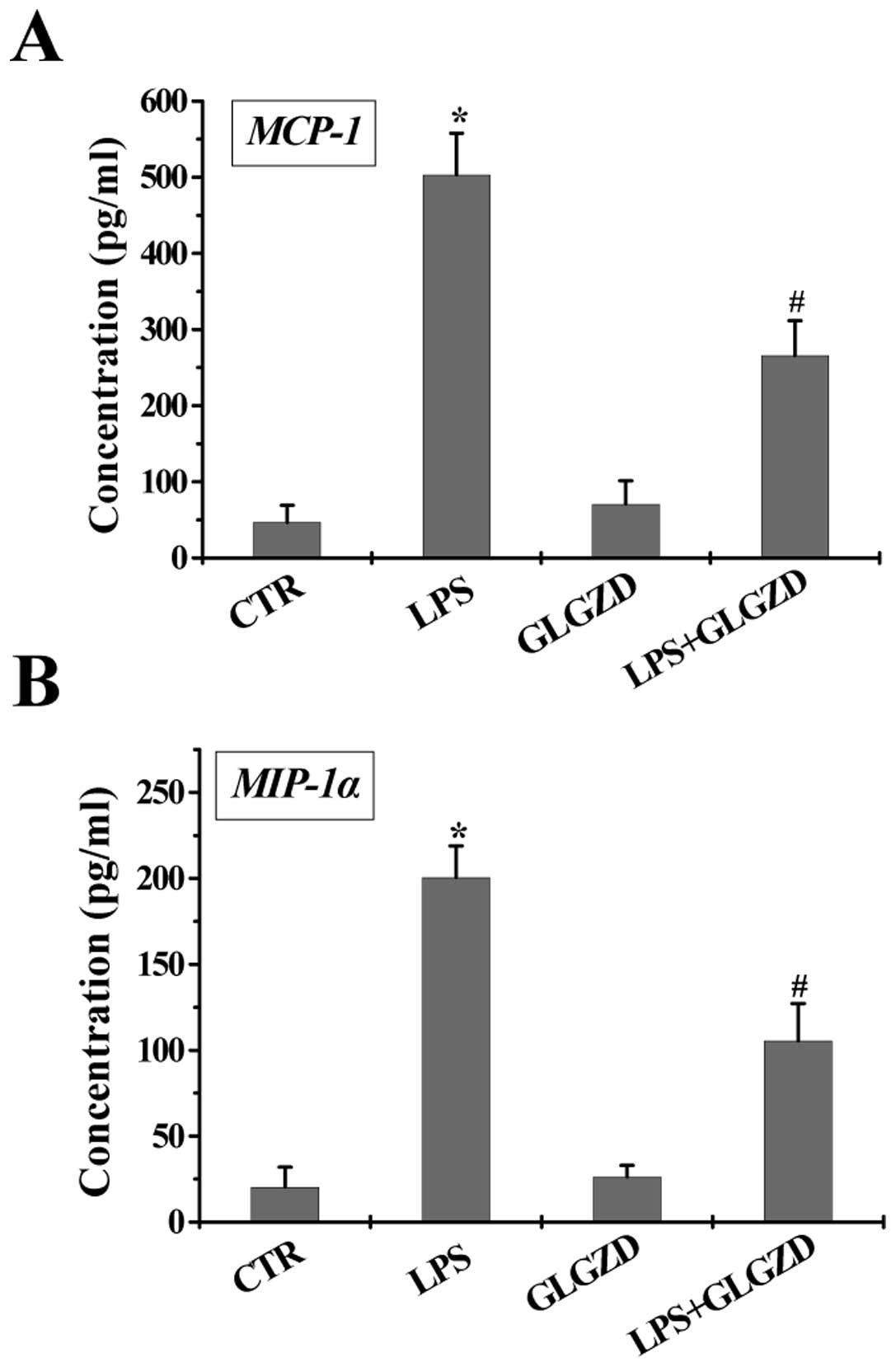
GLGZD attenuates the LPS-stimulated
release of chemotactic cytokines at the transcriptional and
translational levels in BV-2 cells
Activated microglial cells produce chemokines,
inducing immune cells to migrate to the injured brain site
following ischemic stroke. To explore the effects of GLGZD on the
production of several chemokines associated with microglial
motility, following incubation with GLGZD in the presence or
absence of LPS, cell supernatant was collected to examine the
release of chemokines in BV-2 cells. As indicated in Fig. 3A–C, the incubation of BV-2 cells
with LPS (100 ng/ml) for 12 h resulted in a significant increase in
MCP-1, MIP-1α and IL-8 production. The concentrations of these
chemokines were not significantly affected by GLGZD (100 μg/ml)
alone (levels were similar to the control). However, the induction
of these chemokines by LPS was reduced significantly by
co-treatment with GLGZD. In parallel to the protein secretion
levels, the LPS-induced upregulation of MCP-1, MIP-1α and IL-8 mRNA
was attenuated by GLGZD (100 μg/ml), as measured at 12 h (Fig. 3D).
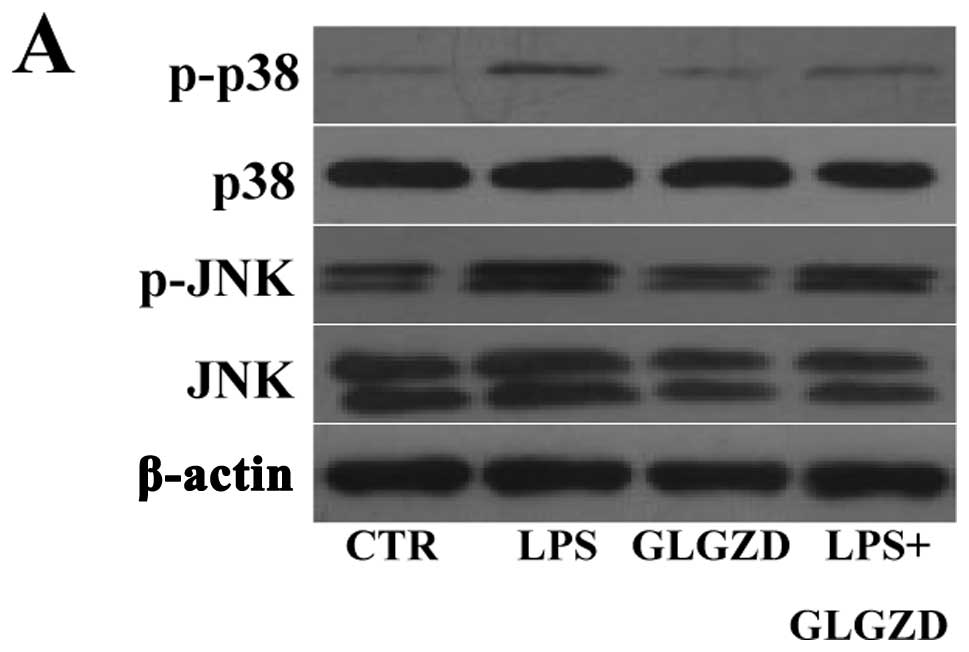
GLGZD supresses the LPS-induced
phosphorylation of MAPK in BV-2 cells
It is well recognized that the LPS-mediated
microglial migration is dependent on the phosphorylation of MAPK.
The members of MAPK are p38, JNK and ERK1/2, which may be involved
in the inhibitory effects of GLGZD on BV-2 microglial activation.
The inhibitory effects of GLGZD on cell motility are very likely
mediated by the MAPK signaling pathway. To futher delineate the
mechanisms of action of GLGZD in LPS-stimulated microglial
migration, we evaluated the phosphorylation of MAPK in BV-2 cells.
The phosphorylation of p38 and JNK induced by LPS (100 ng/ml)
and/or GLGZD (100 μg/ml) was observed. The level of phospho-p38 and
phospho-JNK significantly increased 1 h after the challenge with
LPS and GLGZD significantly reduced the phosphorylation of p38 and
JNK MAPK in the LPS-challenged BV-2 cells (Fig. 4), but did not have any effect on
the phosphorylation of ERK1/2 (data not shown). These results
suggested that JNK and p38 MAPK signaling was required for GLGZD to
suppress the induction of chemokines in microglial cells.
Discussion
Microglial cells are the intrinsic immune cells of
the brain and the main effectors of inflammatory response following
cerebral ischemia (10), playing
important roles in the physiological and pathological conditions of
the CNS. Following injury to the CNS, microglia are rapidly
activated, triggering inflammatory reactions at the sites of injury
(11,12). The migration of microglial cells
is a hallmark of pro-inflammatory and chronic activation during the
early phases of neurodegeneration. Chemokines released from
microglial cells are key components required for cell movement. The
overactivation of microglia, however, can be highly detrimental to
neuronal cells, due to the release of several pro-inflammatory
factors and chemotactic cytokines. Accordingly, the inhibition of
microglial activation and the production of chemotactic cytokines
may be beneficial in reversing microglial-mediated
neurodegeneration.
The recognition that cerebral stroke is a
manifestation of chronic progressive inflammation has had great
influence on the development of therapeutic and preventative
strategies (13,14) and as such, the Chinese formula may
be a promising therapeutic strategy for ameliorating stroke-induced
inflammatory pathological events. GLGZD has been reported to be an
effective treatment for stroke in clinical practice and in animal
experiments (15–17). In our previous study in
vitro, we demonstrated that GLGZD water extract exerts
anti-inflammatory effects in LPS-induced microglial cells mediated
by the nuclear factor-κB (NF-κB) sinaling pathway (9); however, the precise molecular
mechanisms responsible for its neuroprotective effects have not yet
been elucidated. In this regard, we hypothesized that GLGZD may
affect the motility of activated microglia. In the present study,
we used LPS, a bacterial endotoxin used to experimentally study
induced inflammation in vitro (18,19), to promote the migration of
microglial cells, thus creating a cell model of LPS-induced
microglial activation. LPS activates NF-κB and MAPK, which are
classified into at least three components: ERK1/2, JNK and p38 MAPK
(20). Certain studies have
demonstrated that LPS-induced microglial migration requires p38
phosphorylation (21) and that
the PI3K/Akt pathway correlates with microglial migration (22,23). Therefore, in this study, we aimed
to elucidate the mechanisms responsible for the effects of GLGZD on
LPS-induced microglial migration by evaluating the role of the MAPK
signaling pathways.
We demonstrated that LPS (100 ng/ml) and GLGZD (100
μg/ml) had no effect on microglial cell viability; thus, we
selected the concentration of 100 μg/ml GLGZD for the remaining
experiments. Our data demonstrated that GLGZD significantly
inhibited microglial motility in LPS-activated BV-2 cells, as shown
by scratch wound healing assays and Transwell migration assays. To
further assess the effects of GLGZD on LPS-induced microglial
migration, we examined the expression levels of several chemotactic
mediators in BV-2 cells. LPS stimulated microglial cells to secrete
representative chemokines, including MCP-1, MIP-1α and IL-8;
however, GLGZD markedly suppressed the LPS-induced production and
expression of such cytokines (assessed by ELISA and quantitative
RT-PCR). In addition, we sought to examine the modulatory effects
of GLGZD on the MAPK singnaling pathway associated with the
proliferation, survival and migration of microglial cells following
ischemic stroke. We found that within 1 h after the stimulation of
BV-2 cells with LPS, the phosphorylation of JNK and p38 MAPK was
significantly enhanced but was considerably reduced in the cells
co-cultured with LPS and GLGZD, indicating the involvement of JNK
and p38 MAPK in the inhibitory effects of GLGZD on LPS-induced cell
motility.
Based on previous reports and our findings, GLGZD
appears to be effective as an anti-inflammatory agent and exerts
potent protective effects on the activation of microglia in
multiple ways. Collectively, these findings further expand our
current understanding of the regulatory mechanisms of GLGZD under
inflammatory conditions following stroke in vitro, and
provide a new range of therapeutic applications for GLGZD as an
effective inflammatory modulator for the treatment of ischemic
stroke.
Acknowledgements
This study was sponsored by grants from the Guidance
Project of the Fujian Provincial Department of Science and
Technology (no. 2012D012), the Key Project of the Department of
Health of Fujian Province (no. zlckf01) and the Key Project of the
Fujian Provincial Department of Science and Technology (no.
2012Y0041).
Abbreviations:
|
GLGZD
|
Gua Lou Gui Zhi decoction
|
|
CNS
|
central nervous system
|
|
LPS
|
lipopolysaccharide
|
|
MCP-1
|
monocyte chemoattractant protein-1
|
|
MIP-1α
|
macrophage inflammatory protein-1α
|
|
IL-8
|
interleukin-8
|
|
MAPKs
|
mitogen-activated protein kinases
|
|
JNK
|
c-Jun N-terminal protein kinase
|
|
ERK1/2
|
extracellular signal-regulated kinase
1/2
|
References
|
1
|
Shichita T, Ago T, Kamouchi M, Kitazono T,
Yoshimura A and Ooboshi H: Novel therapeutic strategies targeting
innate immune responses and early inflammation after stroke. J
Neurochem. 123(Suppl 2): 29–38. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Shichita T, Sakaguchi R, Suzuki M and
Yoshimura A: Post-ischemic inflammation in the brain. Front
Immunol. 3:1322012. View Article : Google Scholar
|
|
3
|
Garden GA and Möller T: Microglia biology
in health and disease. J Neuroimmune Pharmacol. 1:127–137. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Block ML, Zecca L and Hong JS:
Microglia-mediated neurotoxicity: uncovering the molecular
mechanisms. Nat Rev Neurosci. 8:57–69. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Nam KN, Koketsu M and Lee EH:
5-Chloroacetyl-2-amino-1,3-selenazoles attenuate microglial
inflammatory responses through NF-kappaB inhibition. Eur J
Pharmacol. 589:53–57. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Choi Y, Lee MK, Lim SY, Sung SH and Kim
YC: Inhibition of inducible NO synthase, cyclooxygenase-2 and
interleukin-1beta by torilin is mediated by mitogen-activated
protein kinases in microglial BV2 cells. Br J Pharmacol.
156:933–940. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Choi IY, Lee JC, Ju C, Hwang S, Cho GS,
Lee HW, Choi WJ, Jeong LS and Kim WK: A3 adenosine receptor agonist
reduces brain ischemic injury and inhibits inflammatory cell
migration in rats. Am J Pathol. 179:2042–2052. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
McLarnon JG: Microglial chemotactic
signaling factors in Alzheimer’s disease. Am J Neurodegener Dis.
1:199–204. 2012.PubMed/NCBI
|
|
9
|
Hu H, Li Z, Zhu X, Lin R, Lin J, Peng J,
Tao J and Chen L: Gua Lou Gui Zhi decoction suppresses LPS-induced
activation of the TLR4/NF-κB pathway in BV-2 murine microglial
cells. Int J Mol Med. 31:1327–1332. 2013.PubMed/NCBI
|
|
10
|
Dirnagl U, Iadecola C and Moskowitz MA:
Pathobiology of ischaemic stroke: an integrated view. Trends
Neurosci. 22:391–397. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bethea JR and Dietrich WD: Targeting the
host inflammatory response in traumatic spinal cord injury. Curr
Opin Neurol. 15:355–360. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bareyre FM and Schwab ME: Inflammation,
degeneration and regeneration in the injured spinal cord: insights
from DNA microarrays. Trends Neurosci. 26:555–563. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Valtysson J, Hillered L, Andiné P, Hagberg
H and Persson L: Neuropathological endpoints in experimental stroke
pharmacotherapy: the importance of both early and late evaluation.
Acta Neurochir (Wien). 129:58–63. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Amantea D, Nappi G, Bernardi G, Bagetta G
and Corasaniti MT: Post-ischemic brain damage: pathophysiology and
role of inflammatory mediators. FEBS J. 276:13–26. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang L and Ai H: Effects of Gua Lou Gui
Zhi decoction on c-fos and c-jun on epileptic rats. J Sichuan
Tradit Chin Med. 23:21–22. 2005.
|
|
16
|
Yang C, Chen L and Tao J: New usage of a
classical formula - Gua Lou Gui Zhi decoction. Liaoning J Tradit
Chin Med. 8:166–167. 2012.
|
|
17
|
Huang J, Tao J, Xue X, Yang S, Han P, Lin
Z, Xu W, Lin J, Peng J and Chen L: Gua Lou Gui Zhi decoction exerts
neuroprotective effects on post-stroke spasticity via the
modulation of glutamate levels and AMPA receptor expression. Int J
Mol Med. 31:841–848. 2013.PubMed/NCBI
|
|
18
|
Rivest S: Molecular insights on the
cerebral innate immune system. Brain Behav Immun. 17:13–19. 2003.
View Article : Google Scholar
|
|
19
|
Cohen J: The immunopathogenesis of sepsis.
Nature. 420:885–891. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ajmone-Cat MA, De Simone R, Nicolini A and
Minghetti L: Effects of phosphatidylserine on p38 mitogen activated
protein kinase, cyclic AMP responding element binding protein and
nuclear factor-kappaB activation in resting and activated
microglial cells. J Neurochem. 84:413–416. 2003. View Article : Google Scholar
|
|
21
|
Ferreira R, Santos T, Cortes L, Cochaud S,
Agasse F, Silva AP, Xapelli S and Malva JO: Neuropeptide Y inhibits
interleukin-1 beta-induced microglia motility. J Neurochem.
120:93–105. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Horvath RJ and DeLeo JA: Morphine enhances
microglial migration through modulation of P2X4 receptor signaling.
J Neurosci. 29:998–1005. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Martin S, Vincent JP and Mazella J:
Involvement of the neurotensin receptor-3 in the
neurotensin-induced migration of human microglia. J Neurosci.
23:1198–1205. 2003.PubMed/NCBI
|