Introduction
Psoriasis is considered as a genetic, immunological
skin disease, estimated to affect 2–4% of the population worldwide
(1,2). Histological changes of psoriasis are
characterized by hyperproliferation and poor differentiation of
epidermal keratinocytes, increased skin vascularization and
leukocyte infiltration, including T cells, macrophages, dendritic
cells (DCs) and neutrophils (3).
As a central pathogenic player that activates T cells and produces
cytokines and chemokines in psoriasis (4), DCs in the dermis may be responsible
for inflammatory infiltrates and development of psoriasis (5).
DCs that serve as surveillance cells in the body,
are the most important professional antigen-presenting cells
(APCs), and migrate towards immune organs to present processed
antigens to T cells, initiating primary immune responses (6,7).
Inflammatory DCs in developing psoriasis are the important source
of pro-inflammatory cytokines including interleukin (IL)-23, IL-12,
IL-6, IL-1β and tumor necrosis factor-α (TNF-α) (8,9).
As known, it is through IL-23 and IL-12 that DCs drive
IL-17-producing T cells, such as Th1 and Th22 cells, the activation
of which leads to excessive production of psoriatic cytokines such
as IL-17 and interferon (IFN)-γ mediating effects on keratinocytes
to amplify inflammation (10–14). Controlling these pro-inflammatory
cytokines in DCs would be a breakthrough for psoriasis
treatment.
It is well-known that innate immune cells, such as
DCs, recognize invading pathogens by Toll-like receptors (TLRs) and
respond appropriately to resolve infections (15). TLRs play important roles in the
development of psoriasis, but targeting TLR signaling in DCs
remains a challenge for treating psoriasis. Imiquimod (IMQ) is a
ligand of TLR7/8 and a potent immune activator, which causes
activation and maturation of DCs when applied to the skin of mice
(16,17). TLR signaling involves the
recruitment of adaptor proteins, such as myeloid differentiation
factor 88 (MyD88) (18), and
controlling the MyD88-dependent signaling pathway should throw new
light on the treatment of inflammatory diseases closely related to
DCs, such as psoriasis.
Paeonol is a small-molecule compound derived from a
Chinese herbal plant Cortex Moutan, widely used as a traditional
Chinese medicine (TCM) for alleviation of inflammatory disorders
and it has been proved to treat inflammation, allergies and cancer
(19). However, its effects on
the development and final outcome of these diseases remain unclear.
Numerous drugs containing paeonol are applied as prescribed
medicines for the treatment of eczema, dermatitis, psoriasis and
other skin diseases in China (20). Herein, for the first time, we
showed that paeonol alleviated IMQ-induced psoriasis in mice by
inhibiting the production of pro-inflammatory cytokines such as
IL-23. Bone marrow-derived dendritic cells (BMDCs) stimulated with
R848 and treated with paeonol were also investigated. Therefore,
control of maturation and activation of DCs by decreasing MyD88 and
TLR8 proteins in the TLR7/8 signaling pathway by paeonol could be a
novel strategy to treat psoriasis.
Materials and methods
Psoriatic model in mice
The 8- to 10-week-old-male BALB/c mice (18–20 g)
were supplied by Beijing HFK Bioscience Co., Ltd. (Beijing, China)
(certification no. SCXK Jing 2014-0004), and maintained with free
access to food and water under specific pathogen-free
conditions.
The mice were divided into six groups of eight mice
each. Five groups were administered a daily topical dose of 42 mg
of a cream preparation containing 5% IMQ (Mingxinlidi Laboratory,
China) on their hair-free backs to establish a model of IMQ-induced
psoriasis. The control group (Con) received appropriate vaseline.
Paeonol (National Institutes for Food and Drug Control, Beijing,
China), was dissolved in normal saline (NS) to achieve different
concentrations for oral administration. The model group (IMQ)
received saline and the methotrexate (MTX) group received 1 mg/kg
MTX, a drug used for psoriasis treatment. The paeonol-high (PH)
group received 100 mg/kg paeonol, the paeonol-medium (PM) group 50
mg/kg paeonol, and the paeonol-low (PL) group 25 mg/kg paeonol. All
treatments were administered (0.4 ml/day) from the day IMQ was
applied, once a day for a week. After 7 days, the mice were
sacrificed by cervical dislocation under sodium pentobarbital
anesthesia, while skin lesions and serum samples were collected.
All animal experiments were performed in accordance with the Guide
for the Care and Use of Laboratory Animals, formulated by the
National Institutes of Health (Bethesda, MA, USA), and approved by
the Office of the Experimental Animal Management Committee
(Beijing, China) and the local animal ethics committee.
Isolation of bone marrow cells and in
vitro induction and culture of BMDCs
BMDCs were induced from bone marrow cells of C57/BL6
mice by flushing the femurs and tibiae of 6- to 10-week-old mice
with phosphate-buffered saline (PBS). Pooled cells from four limbs
were washed in RPMI-1640 medium after lysis of red blood cells, and
then plated in 6-well Petri dishes (1×107/well) with
RPMI-1640 medium supplemented with 10% fetal bovine serum (FBS)
(Gibco Life Technologies, Grand Island, NY, USA), 1%
penicillin-streptomycin, 20 ng/ml IL-4 and 20 ng/ml GM-CSF (both
from PeproTech, Rocky Hill, NJ, USA), cultured for 7 days to induce
DCs and the supplemented medium was replaced every day.
CD11c+ DCs were sorted by magnetic beads and seeded in
6-well plates on day 8. BMDCs were then induced to maturation by
R848 (1 μg/ml). Different concentrations of paeonol (75,
35.5 and 17.75 μg/ml) were added into the medium, and we
collected the cells after 24 h.
Severity scoring of skin
inflammation
The severity of skin inflammation was monitored and
graded using a modified human scoring system Psoriasis Area
Severity Index (PASI). Scaling, thickness and erythema were scored
separately on a scale from 0 to 4: 0, none; 1, slight; 2, moderate;
3, marked; and 4, very marked. The total score denotes severity of
inflammation.
Histopathological examination and
immunochemical, immunofluorescence studies
Mice were sacrificed by cervical dislocation under
sodium pentobarbital anesthesia after one week. The skin lesions
were removed, fixed in 10% formalin and embedded in paraffin.
Sections (5-μm) were stained with hematoxylin
and eosin (H&E). The staining was assessed by light microscope
(Olympus, Tokyo, Japan) and epidermal thickness was measured by
Image-Pro Plus 6.0 software (Media Cybernetics, Inc., Rockville,
MD, USA).
For immunostaining, skin sample sections from the
back lesions were stained with anti-rabbit proliferating cell
nuclear antigen (PCNA) (Cat. no. ab15497; diluted 1:100), CD3 (Cat.
no. ab16669; diluted 1:100) and CD11c (Cat. no. ab33483; diluted
1:500) antibodies (all from Abcam, Cambridge, UK) and staining was
assessed using light and fluorescence microscopes (Olympus).
Cell viability assay
The effects of paeonol on cell viability were
assessed using the Cell Counting Kit-8 (CCK-8) assay (Dojindo
Laboratories, Kumamoto, Japan) according to the manufacturer's
instructions. Cells were seeded in 96-well plates and treated with
different paeonol concentrations for 24 h. The plates were
incubated at 37°C in 5% CO2 for 2 h. The mean optical
density (OD) of the cells in each group was used to identify the
non-toxic concentration of paeonol.
Real-time polymerase chain reaction
(RT-PCR)
Total RNA was extracted from skin lesions and BMDCs
using TRIzol (Invitrogen Life Technologies, Carlsbad, CA, USA) and
purified using a NucleoSpin RNA clean-up kit (Macherey-Nagel,
Germany). Following the generation of complementary DNA using an
AffinityScript multiple temperature cDNA synthesis kit (Agilent
Technologies, Inc., Santa Clara, CA, USA), the relative expression
levels of genes were determined with an ABI 7500 Fast Real-Time PCR
system using real-time PCR Master Mix (Roche Diagnostics,
Indianapolis, IN, USA). The gene-specific primers are listed in
Table I. Cycle parameters were as
follows: 95°C for 10 min; 50 cycles of 95°C for 15 sec, and 60°C
for 60 sec. The β-actin gene was used as a reference to normalize
the data that was quantitatively analyzed using the
2−ΔΔCq method.
 | Table IPrimers used for RT-PCR. |
Table I
Primers used for RT-PCR.
| Primer
sequences |
|---|
| IL-23 | F:
5′-ACTCCCCATTCCTACTTCTCCCT-3′ |
| R:
5′-CACTTGCTGCATGAGGAATTGTA-3′ |
| IL-1β | F:
5′-TGCCACCTTTTGACAGTGATGA-3′ |
| R:
5′-TGTGCTGCTGCGAGATTTGA-3′ |
| IL-12 | F:
5′-TCAACGCAGCACTTCAGAATCACAA-3′ |
| R:
5′-GAAGGCGTGAAGCAGGATGCAGAGC-3′ |
| β-actin | F:
5′-CGTTGACATCCGTAAAGACCTC-3′ |
| R:
5′-ACAGAGTACTTGCGCTCAGGAG-3′ |
Western blotting
Skin samples were lysed and the protein was resolved
by 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis
(SDS-PAGE). The membrane fractions were incubated with MyD88 (4283;
Cat. no. 4283; Cell Signaling Technology, Inc., Danvers, MA, USA),
rabbit-TLR8 (Cat. no. ab180610; Abcam) and mouse anti-β-tubulin
(Cat. no. YM3030; Immunoway) antibodies, followed by IRDye 700DX-
or 800DX-conjugated secondary antibodies (Rockland Immunochemicals
Inc., Gilbertsville, PA, USA). Immunofluorescence was assessed by
Odyssey infrared imaging system (LI-COR Biosciences, Lincoln, NE,
USA).
Flow cytometric analysis
Spleen samples from each group were harvested to
determine whether the frequency of DCs was altered in the spleens
of the IMQ-induced mouse model following treatment with paeonol.
Following, the samples were minced through a 70-μm mesh to
obtain single-cell suspensions, and 1×106 cells were
stained with fluorescein isothiocyanate (FITC)-conjugated mouse
anti-CD11c. Samples were analyzed using flow cytometry (FCM).
To determine whether paeonol altered the ratio of
mature and activated DCs induced by R848 in vitro, BMDCs
from different groups were suspended in PBS, and then stained with
anti-CD11c-PE (Cat. no. ab155349), anti-MHCII-FITC (Cat. no.
ab93561), anti-CD80-FITC (Cat. no. ab24860) and anti-CD86-FITC
(Cat. no. ab24862) antibodies at 4°C for 30 min in the dark. Cells
were washed once with PBS and analyzed with FCM.
ELISA
BMDCs were seeded in 12-well culture plates at a
density of 1×106 cells/ml and pretreated with various
concentrations (75, 35.5 and 17.75 μg/ml) of paeonol for 24
h, followed by R848 (Cat. no. SML0196; Sigma, St. Louis, MO, USA)
(1 μg/ml) for 24 h. The levels of IL-12p40 and IL-12p70 in
the supernatants were determined using mouse ELISA kits according
to the manufacturer's instructions, with standard curves made from
purified recombinant IL-12p40 and IL-12p70 at various
dilutions.
Statistical analysis
Results are expressed as mean ± SD. Differences
between groups were evaluated with Student's t-test (for
comparisons between two samples) or by a one-way analysis of
variance (ANOVA) (for comparisons of multiple samples) using the
SPSS 15.0 software (SPSS, Inc., Chicago, IL, USA). A p-value
<0.05 was considered to indicate a statistically significant
difference.
Results
Paeonol improves psoriatic lesions in a
mouse model by inhibiting proliferation and differentiation of
keratinocytes
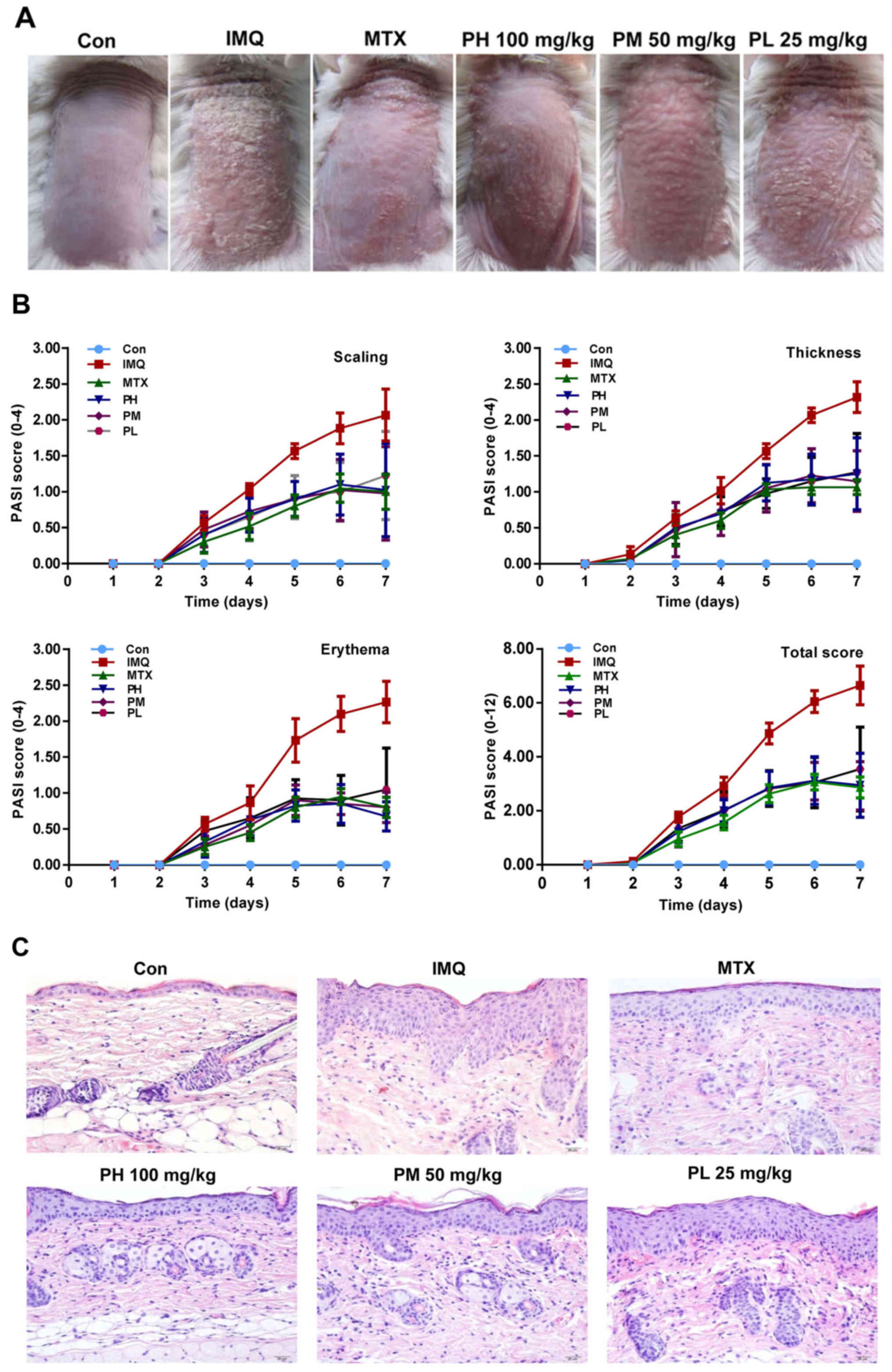
We initially investigated the effect of paeonol
(Fig. 1) on the psoriatic mouse
model induced by IMQ through morphological observations and
pathological slices. Typical erythema, scaling and thickening were
observed in the IMQ-induced skin lesions as compared to the control
group, while paeonol significantly inhibited these pathological
changes in a dose-dependent manner (Fig. 2A and B).
Skin treated with IMQ demonstrated pathological
changes of the epidermal cuticle, including significant
parakeratosis, acanthosis and perivascular infiltration of
inflammatory cells in the upper dermis, a phenotype typical of
human psoriatic skin. Paeonol significantly reduced the thickness
of the epidermis, and attenuated IMQ-induced psoriasis (Fig. 2C and E).
PCNA is expressed in proliferative cells, especially
basal cells. Expression of PCNA was reduced in the skin lesions of
the paeonol-treated mice, suggesting that paeonol reduces
IMQ-induced proliferation of keratinocytes (Fig. 2D and F) and effectively
ameliorates IMQ-induced keratinocyte differentiation.
Paeonol downregulates inflammatory
infiltration especially mature and activated DCs along with
pro-inflammatory cytokines in a mouse model of psoriasis
CD3 is mainly expressed in T lymphocytes and appears
as brown particles after immunohistochemistry with the DAB
chromogenic agent. Infiltration of CD3+ cells was
observed in IMQ-induced lesions but was reduced in the
paeonol-treated mice. Fig. 3B
shows the expression of CD3+ cells, not IL-23 mRNA in
the skin lesions, and it is the statistical data for Fig. 3A.
The numbers of CD11c+ DCs in the dorsal
skin and spleens of IMQ-induced mice were detected by
immunofluorescence assay (IFA), which showed a major increase in
these cells in the dorsal skin (Fig.
3C and D) as well as in the spleens, which were markedly
increased (Fig. 3E and F).
Compared with the control group, the mRNA expression
level of IL-23 was markedly increased in the IMQ-induced psoriatic
skins (Fig. 3G). As expected,
expression in the group treated with paeonol was significantly
decreased.
Paeonol decreases MyD88 and TLR8 proteins
in a mouse model of psoriasis
We further explored how paeonol inhibits the TLR7/8
signaling pathway, and found lower protein expression of MyD88 and
TLR8 in skin lesions of the paeonol-treated group as compared to
these levels in the IMQ-induced model (Fig. 4A and B).
Paeonol inhibits the maturation and
activation of BMDCs
We stimulated BMDCs with 1 μg/ml R848.
Surface markers of DCs including CD11c, MHCII, CD80 and CD86 were
detected by flow cytometry. MHCII, CD80 and CD86 were downregulated
in the paeonol-treated mice (Fig.
5).
Paeonol decreases the expression of
pro-inflammatory cytokines secreted by mature BMDCs
The mRNA levels of IL-23, IL-12 and IL-1β were
measured by RT-PCR and related cytokines were assessed by ELISA.
RT-PCR showed that IL-23 was significantly suppressed, while IL-12
and IL-1β were enhanced after paeonol treatment in a dose-dependent
manner, while cytokine levels were decreased by paeonol (Fig. 6A and B).
Discussion
Psoriasis is a chronic autoimmune disease mediated
by immune cells and molecules, with persistent inflammation and
hyperplasia (21). Suppression of
T cell responsiveness by influencing the phenotype and function of
DCs, the most powerful APCs, remains one of the most effective
treatment approaches (22). As
bridges that link innate and adaptive immunity, activated and
mature DCs lead to the next phase of immune reactions under
physiological circumstances, removing pathogens from the body
(23). However, inappropriate
activation and maturation of DCs are closely related to the
development of immune disorders and multiple pro-inflammatory
cytokines such as IL-23, IL-12 and IL-1β in the psoriatic skin are
secreted by DCs (24). Symptoms
could be relieved if signaling pathways of DCs are inhibited.
TLRs play a central role in the recognition and
response to invading pathogens by immune cells, leading to the
production of pro-inflammatory cytokines and chemokines (25). The TLR7/8, signaling pathway,
which includes recruitment of MyD88, could be an important target
for treating psoriasis. The role of MyD88 protein in the TLR7/8
signaling pathway has been established (26,27). Blocking of the MyD88-dependent
signaling pathway aids in the treatment of psoriasis.
IMQ is a TLR7/8 agonist, and the IMQ-induced mouse
model is a classic model of psoriasis (28). Mice administered paeonol showed
lower scores for PASI, thinner epidermis and decreased keratinocyte
proliferation and differentiation, which were positively correlated
with the paeonol concentration (Fig.
2). We believe that paeonol can be potentially used for
treating psoriasis in a dose-dependent manner.
The role of the IL-23/Th17 axis in the development
of psoriasis is a current focus of research (29). However, we are more interested in
the upstream pathway. Research has shown that CD11c is regarded as
the correct marker of myeloid DCs (30) and the number of CD11c+
cells was lower in the spleens of the paeonol-treated mice
(Fig. 3), consistent with the
data from the immunofluorescence assay of skin lesions indicating
that paeonol decreased the number of DCs. IL-23, mostly secreted by
mature and activated DCs, is responsible for the local infla
mmation of psoriatic skin, leading us to focus on DCs (31). Other studies have shown that
pro-inflammatory factors such as TNF-α, MCP-1, IL-1β and IL-6 were
dose-dependently reduced by paeonol treatment in vivo
(32–35). Results from skin lesions showed
that expression of IL-23 mRNA decreased in a dose-dependent manner
(Fig. 3G), as previously
reported. We further detected the expression of MyD88 and TLR8
proteins (Fig. 4), which were
significantly reduced in the paeonol-treated group.
BMDCs were collected for further study concerning
the frequency and the phenotype of DCs. MHCII, CD80 and CD86 are
all specific markers for mature DCs (36). BMDCs cultured for 1 week were
stimulated with R848, a TLR8 agonist that induces DC maturation for
24 h, and were then treated with paeonol. The number of
marker-positive cells following paeonol therapy was significantly
reduced as compared to the model group (Fig. 5). Results from the in vitro
experiments indicated that paeonol improved the clinical symptoms
of psoriasis by suppressing the maturation and activation of DCs.
CD80 and CD86 are induced in an MyD88-dependent manner in the TLR7
signaling pathway, and the IMQ-induced mouse model is characterized
by activated TLR7. Moreover, R848 used in our in vitro
experiment is a TLR8 agonist. Meanwhile, expression of MyD88 and
TLR8 proteins was significantly reduced in the paeonol-treated
group. Thus, paeonol plays a role in anti-inflammation of psoriasis
by inhibiting maturation and activation of DCs through the TLR7/8
signaling pathway in DCs.
In order to validate the suppressive effects on
inflammation in BMDCs, we also detected the expression of IL-23,
IL-12 and IL-1β, cytokines that cause specific T cell responses
(36). Results from the BMDCs
showed that expression of IL-23 mRNA decreased in a dose-dependent
manner, consistent with our results in the skin lesions (Fig. 6). Meanwhile, ELISA for IL-12p40 in
BMDCs indicated that the levels of IL-23 were decreased. IL-23,
composed of two chains, the unique p19 chain and the p40 chain
shared with IL-12 is required for T cells to produce IL-17.
IL-12p40 represented IL-23 in the ELISA kit we used. Conversely,
IL-12 and IL-1β were opposite of what we expected, which was in
disagreement with a previous study (37). Strangely, groups treated with
paeonol showed higher expression of IL-12 and IL-1β genes than the
model, which was dose-dependent. Perhaps, paeonol specifically acts
on the TLR7/8 signaling pathway, a MyD88-dependent pathway in DCs,
and inhibits the maturation and activation of DCs to reduce the
secretion of IL-23. Also, we suspect that the role of paeonol
varies over time.
These findings imply that paeonol may be used to
treat inflammatory and hyperplastic diseases. We explored a novel
strategy by which to treat psoriasis by decreasing MyD88, a key TLR
adaptor protein, blocking the TLR7/8-mediated signaling pathway in
DCs and contributing to the improvement of IMQ-induced psoriasis in
mice.
Recent research progress on the inhibitory activity
of paeonol on the inflammatory reaction is significant (38,39). The mechanism by which paeonol
decreases MyD88 and TLR proteins, either by inducing degradation of
these proteins or blocking their synthesis, remains unknown. We
will continue to explore the precise mechanisms underlying the
regulation of DCs in psoriasis.
Acknowledgments
The present study was supported by the National
Natural Science Foundation of China (nos. 81403410 and
81573974).
References
|
1
|
Parisi R, Symmons DPM, Griffiths CEM and
Ashcroft DM; Identification and Management of Psoriasis and
Associated ComorbidiTy (IMPACT) project team: Global epidemiology
of psoriasis: a systematic review of incidence and prevalence. J
Invest Dermatol. 133:377–385. 2013. View Article : Google Scholar
|
|
2
|
Ben Salem C, Hmouda H and Bouraoui K:
Psoriasis. N Engl J Med. 361:17102009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lima Ede A and Lima Mde A: Reviewing
concepts in the immunopathogenesis of psoriasis. An Bras Dermatol.
86:1151–1158. 2011.
|
|
4
|
Zaba LC, Fuentes-Duculan J, Eungdamrong
NJ, Abello MV, Novitskaya I, Pierson KC, Gonzalez J, Krueger JG and
Lowes MA: Psoriasis is characterized by accumulation of
immunostimulatory and Th1/Th17 cell-polarizing myeloid dendritic
cells. J Invest Dermatol. 129:79–88. 2009. View Article : Google Scholar
|
|
5
|
Kim J and Krueger JG: The
immunopathogenesis of psoriasis. Dermatol Clin. 33:13–23. 2015.
View Article : Google Scholar
|
|
6
|
Mahil SK, Capon F and Barker JN: Update on
psoriasis immunopathogenesis and targeted immunotherapy. Semin
Immunopathol. 38:11–27. 2016. View Article : Google Scholar :
|
|
7
|
Nestle FO, Turka LA and Nickoloff BJ:
Characterization of dermal dendritic cells in psoriasis.
Autostimulation of T lymphocytes and induction of Th1 type
cytokines. J Clin Invest. 94:202–209. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Nestle FO, Di Meglio P, Qin JZ and
Nickoloff BJ: Skin immune sentinels in health and disease. Nat Rev
Immunol. 9:679–691. 2009.PubMed/NCBI
|
|
9
|
Krueger JG and Bowcock A: Psoriasis
pathophysiology: current concepts of pathogenesis. Ann Rheum Dis.
64(Suppl 2): 30–36. 2005. View Article : Google Scholar
|
|
10
|
Lowes MA, Suárez-Fariñas M and Krueger JG:
Immunology of psoriasis. Annu Rev Immunol. 32:227–255. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Di Cesare A, Di Meglio P and Nestle FO:
The IL-23/Th17 axis in the immunopathogenesis of psoriasis. J
Invest Dermatol. 129:1339–1350. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bettelli E, Oukka M and Kuchroo VK:
T(H)-17 cells in the circle of immunity and autoimmunity. Nat
Immunol. 8:345–350. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Arican O, Aral M, Sasmaz S and Ciragil P:
Serum levels of TNF-alpha, IFN-gamma, IL-6, IL-8, IL-12, IL-17, and
IL-18 in patients with active psoriasis and correlation with
disease severity. Mediators Inflamm. 2005:273–279. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Mitra A, Fallen RS and Lima HC:
Cytokine-based therapy in psoriasis. Clin Rev Allergy Immunol.
44:173–182. 2013. View Article : Google Scholar
|
|
15
|
Akira S, Uematsu S and Takeuchi O:
Pathogen recognition and innate immunity. Cell. 124:783–801. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Dika E, Varotti C, Bardazzi F and Maibach
HI: Drug-induced psoriasis: an evidence-based overview and the
introduction of psoriatic drug eruption probability score. Cutan
Ocul Toxicol. 25:1–11. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Stary G, Bangert C, Tauber M, Strohal R,
Kopp T and Stingl G: Tumoricidal activity of TLR7/8-activated
inflammatory dendritic cells. J Exp Med. 204:1441–1451. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
O'Neill LA, Golenbock D and Bowie AG: The
history of toll-like receptors - redefining innate immunity. Nat
Rev Immunol. 13:453–460. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
19
|
Liu MH, Lin AH, Lee HF, Ko HK, Lee TS and
Kou YR: Paeonol attenuates cigarette smoke-induced lung
inflammation by inhibiting ROS-sensitive inflammatory signaling.
Mediators Inflamm. 2014:6518902014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chou TC: Anti-inflammatory and analgesic
effects of paeonol in carrageenan-evoked thermal hyperalgesia. Br J
Pharmacol. 139:1146–1152. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Elder JT, Bruce AT, Gudjonsson JE,
Johnston A, Stuart PE, Tejasvi T, Voorhees JJ, Abecasis GR and Nair
RP: Molecular dissection of psoriasis: integrating genetics and
biology. J Invest Dermatol. 130:1213–1226. 2010. View Article : Google Scholar
|
|
22
|
Kim ME, Kim HK, Park HY, Kim DH, Chung HY
and Lee JS: Baicalin from Scutellaria baicalensis impairs Th1
polarization through inhibition of dendritic cell maturation. J
Pharmacol Sci. 121:148–156. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bieber K and Autenrieth SE: Insights how
monocytes and dendritic cells contribute and regulate immune
defense against microbial pathogens. Immunobiology. 220:215–226.
2015. View Article : Google Scholar
|
|
24
|
Domingues R, de Carvalho GC, Aoki V, da
Silva Duarte AJ and Sato MN: Activation of myeloid dendritic cells,
effector cells and regulatory T cells in lichen planus. J Transl
Med. 14:1712016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Nahid MA, Benso LM, Shin JD, Mehmet H,
Hicks A and Ramadas RA: TLR4, TLR7/8 agonist-induced miR-146a
promotes macrophage tolerance to MyD88-dependent TLR agonists. J
Leukoc Biol. 100:339–349. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Shao F, Tan T, Tan Y, Sun Y, Wu X and Xu
Q: Andrographolide alleviates imiquimod-induced psoriasis in mice
via inducing autophagic proteolysis of MyD88. Biochem Pharmacol.
115:94–103. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yu D, Shi M, Bao J, Yu X, Li Y and Liu W:
Genipin ameliorates hypertension-induced renal damage via the
angiotensin II-TLR/MyD88/MAPK pathway. Fitoterapia. 112:244–253.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Singh M, Khong H, Dai Z, Huang XF, Wargo
JA, Cooper ZA, Vasilakos JP, Hwu P and Overwijk WW: Effective
innate and adaptive antimelanoma immunity through localized TLR7/8
activation. J Immunol. 193:4722–4731. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Singh TP, Schön MP, Wallbrecht K,
Michaelis K, Rinner B, Mayer G, Schmidbauer U, Strohmaier H, Wang
XJ and Wolf P: 8-Methoxypsoralen plus ultraviolet A therapy acts
via inhibition of the IL-23/Th17 axis and induction of
Foxp3+ regulatory T cells involving CTLA4 signaling in a
psoriasis-like skin disorder. J Immunol. 184:7257–7267. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zaba LC, Fuentes-Duculan J, Steinman RM,
Krueger JG and Lowes MA: Normal human dermis contains distinct
populations of CD11c+BDCA-1+ dendritic cells
and CD163+FXIIIA+ macrophages. J Clin Invest.
117:2517–2525. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kollipara R, Downing C, Gordon R and
Tyring S: Interleukin-23 in the pathogenesis and treatment of
psoriasis. Skin Therapy Lett. 20:1–4. 2015.PubMed/NCBI
|
|
32
|
Nam KN, Woo BC, Moon SK, Park SU, Park JY,
Hwang JW, Bae HS, Ko CN and Lee EH: Paeonol attenuates
inflammation-mediated neurotoxicity and microglial activation.
Neural Regen Res. 8:1637–1643. 2013.PubMed/NCBI
|
|
33
|
Hsieh CL, Cheng CY, Tsai TH, Lin IH, Liu
CH, Chiang SY, Lin JG, Lao CJ and Tang NY: Paeonol reduced cerebral
infarction involving the superoxide anion and microglia activation
in ischemia-reperfusion injured rats. J Ethnopharmacol.
106:208–215. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Tao W, Wang H, Su Q, Chen Y, Xue W, Xia B,
Duan J and Chen G: Paeonol attenuates lipopolysaccharide-induced
depressive-like behavior in mice. Psychiatry Res. 238:116–121.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Meng L, Xu W, Guo L, Ning W and Zeng X:
Paeonol Inhibits the proliferation, invasion, and inflammatory
reaction induced by TNF-α in vascular smooth muscle cells. Cell
Biochem Biophys. 73:495–503. 2015. View Article : Google Scholar
|
|
36
|
Lin C, Lin HY, Chen JH, Tseng WP, Ko PY,
Liu YS, Yeh WL and Lu DY: Effects of paeonol on
anti-neuroinflammatory responses in microglial cells. Int J Mol
Sci. 16:8844–8860. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kim HW, Cho SI, Bae S, Kim H, Kim Y, Hwang
YI, Kang JS and Lee WJ: Vitamin C up-regulates expression of CD80,
CD86 and MHC class II on dendritic cell line, DC-1 via the
activation of p38 MAPK. Immune Netw. 12:27–283. 2012. View Article : Google Scholar
|
|
38
|
Fu PK, Wu CL, Tsai TH and Hsieh CL:
Anti-inflammatory and anticoagulative effects of paeonol on
LPS-induced acute lung injury in rats. Evid Based Complement
Alternat Med. 2012:8375132012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Jin X, Wang J, Xia ZM, Shang CH, Chao QL,
Liu YR, Fan HY, Chen DQ, Qiu F and Zhao F: Anti-inflammatory and
anti-oxidative activities of paeonol and its metabolites through
blocking MAPK/ERK/p38 signaling pathway. Inflammation. 39:434–446.
2016. View Article : Google Scholar
|