Introduction
Epidemiological and clinical evidence has indicated
a substantially higher risk of urinary bladder cancer in males yet
there is a tendency showing more aggressive behavior in tumors from
female patients (1,2). Recent experimental data suggest that
urothelial carcinoma, like prostate and breast cancers, is an
endocrine-related neoplasm (reviewed in ref. 3). In particular, the androgen receptor
(AR) and estrogen receptor (ER) signaling pathways have been shown
to contribute to bladder tumorigenesis and cancer progression
(3–13), which may explain some of the
differences in male versus female bladder cancer.
Activation of the epidermal growth factor (EGF)
receptor (EGFR) family is known to involve the growth and
progression of a variety of malignancies. In bladder cancer,
EGFR/ERBB2 is frequently overexpressed, which correlates with
higher tumor grade/stage and poorer prognosis (14–16).
Experimental evidence in bladder cancer has also suggested that the
EGFR pathway plays a critical role in cell proliferation,
apoptosis, differentiation, migration and angiogenesis (17–19).
Consequently, the efficacy of targeted therapy directed at EGFR
signals has been assessed in bladder cancer.
The crosstalk between nuclear hormone receptors and
growth factors leads to activation of nuclear receptor-mediated
transcription. Specifically, in prostate cancer cells, AR signals
upregulate EGFR and ERBB2 gene expression, whereas
activation of EGFR and ERBB2 modulates AR functions (20–24).
It has also been shown that the assembly of the EGFR/AR/ER/Src
signaling complex is crucial for proliferation of prostate and
breast cancer cells triggered by androgens, estrogens and/or EGF
(25). In contrast, the
relationship between the AR and EGFR pathways in bladder cancer
remains poorly understood. We have recently shown that AR
activation results in upregulation of EGFR and ERBB2 expression in
bladder cancer cells, which may play an important role in
androgen-mediated tumor progression (26). In the present study, we
investigated whether EGF could alter AR activity in bladder cancer
cells.
Materials and methods
Cell culture and chemicals
Human bladder cancer cell lines, UMUC3, 5637 and
J82, obtained from the American Type Culture Collection (Manassas,
VA, USA) were maintained in Dulbecco’s modified Eagle’s medium
(Mediatech, Manassas, VA, USA) supplemented with 10% fetal bovine
serum (FBS) at 37°C in a humidified atmosphere of 5%
CO2. Cells were cultured in phenol-red free medium
supplemented with 5% charcoal-stripped FBS at least 18 h before
experimental treatment. We obtained dihydrotestosterone (DHT) and
EGF from Sigma (St. Louis, MO, USA); hydroxyflutamide (HF) from
Schering (Kenilworth, NJ, USA); and PD168393 from Calbiochem (San
Diego, CA, USA).
Stable cell lines with AR and AR-short
hairpin RNA (shRNA)
Cell lines stably expressing a full-length wild-type
human AR (5637-AR and J82-AR) or vector only (5637-V and J82-V)
were established, using a lentivirus vector (pWPI-AR or
pWPI-control) with psPAX2 envelope and pMD2.G packaging plasmids,
as we described previously (11,26).
Similarly, stable AR knockdown/control cell lines
(UMUC3-AR-shRNA/UMUC3-control-shRNA) were established with a
retrovirus vector pMSCV/U6-AR-shRNA or pMSCV/U6-control-shRNA
(5,26).
Reporter gene assay
Bladder cancer cells at a density of 50–60%
confluence in 24-well plates were co-transfected with 250 ng of
MMTV-luc reporter plasmid DNA and 2.5 ng of pRL-TK-luc plasmid DNA,
using GeneJuice transfection reagent (Novagen, Gibbstown, NJ, USA).
Six hours after transfection, the medium was replaced with one
supplemented with 5% charcoal-stripped FBS containing ethanol or
ligands (DHT, HF, EGF and/or PD168393) for 24 h. Cells were
harvested, lysed and assayed for luciferase activity determined
using a dual-luciferase reporter assay kit (Promega, Madison, WI,
USA) and luminometer (TD-20/20; Turner BioSystems, Sunnyvale, CA,
USA).
Cell proliferation assay
We used the MTT (methyl thiazolyl diphenyl
tetrazolium bromide) assay to assess cell viability, as described
previously (26,27). Briefly, cells (3×103)
seeded in 96-well tissue culture plates were incubated with medium
supplemented with charcoal-stripped FBS in the presence or absence
of ligands (DHT, HF and EGF). The media were refreshed every 24 h.
After 96 h of treatment, 10 μl MTT (Sigma) stock solution (5
mg/ml) was added to each well with 0.1 ml of medium for 4 h at
37°C. The medium was replaced with 100 μl DMSO followed by
incubation for 5 min at room temperature. The absorbance was then
measured at a wavelength of 570 nm with background subtraction at
655 nm.
Western blotting
Protein extraction and western blotting were
performed, as described previously (27) with minor modifications. Briefly,
equal amounts of protein (20 μg) obtained from cell extracts
were separated in a 10% sodium dodecylsulfate (SDS)-polyacrylamide
gel electrophoresis (PAGE) and transferred to polyvinylidene
difluoride membrane (Millipore, Billerica, MA, USA) by
electroblotting using a standard protocol. Specific antibody
binding was detected, using an anti-AR antibody (clone N20; diluted
1:2,000; Santa Cruz Biotechnology, Santa Cruz, CA, USA), an
anti-transcriptional intermediary factor 2 (TIF2) antibody (clone
29/TIF2; diluted 1:1,000; BD Bioscience, Franklin Lakes, NJ, USA),
or an anti-GAPDH antibody (clone 6C5; diluted 1:1,000; Santa Cruz
Biotechnology), with horseradish peroxidase detection system
(SuperSignal West Pico Chemiluminescent Substrate; Thermo
Scientific, Rockford, IL, USA).
Co-immunoprecipitation
UMUC3 cells were treated with mock (ethanol) or EGF
for 24 h and protein (500 μg) from the cell lysates was
incubated with 2 μg of anti-AR antibody (N20) or normal
rabbit IgG (Santa Cruz Biotechnology) for 16 h at 4°C with
agitation. To each sample we added 20 μl of protein
A/G-agarose beads (Santa Cruz Biotechnology), incubated for 1 h and
washed thrice with radio-immunoprecipitation assay buffer. Then,
the complex was resolved on a 10% SDS-PAGE, transferred to the
membrane and blotted with an anti-ERβ antibody (clone 14C8; diluted
1:500; Abcam, Cambridge, MA, USA) or an anti-v-Src antibody (clone
327; diluted 1:1,000; Calbiochem).
Statistical analysis
Student’s t-test was used to analyze differences in
relative luciferase activity and relative cell number between the
two groups. P<0.05 was considered statistically significant.
Results
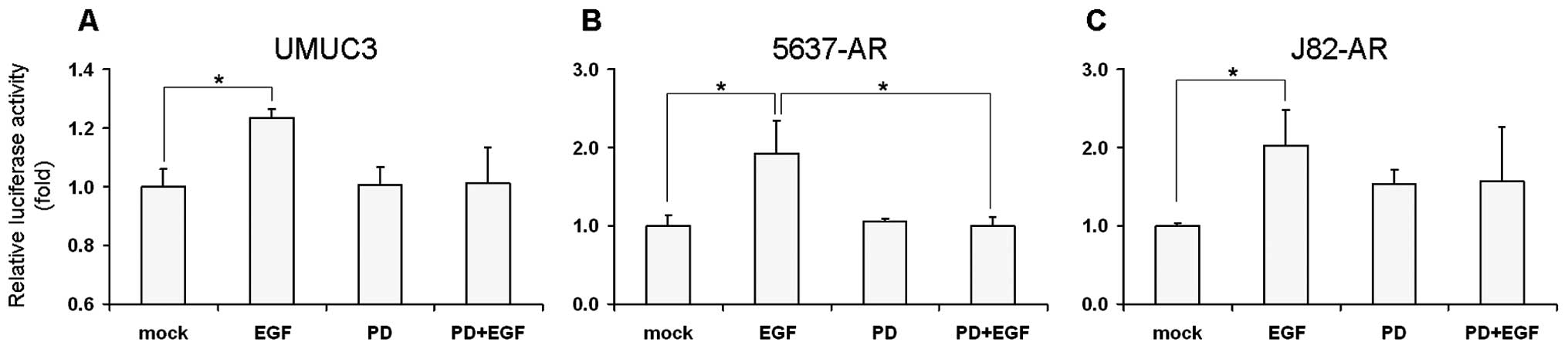
EGF mediates AR transactivation via
EGFR
Because previous studies showed ligand-independent
activation of AR transcription by EGF in prostate cancer cells
(20–22), we first assessed the effects of EGF
and a specific EGFR inhibitor PD168393 on AR transactivation in
bladder cancer lines. In AR-positive UMUC3 and AR-negative 5637 and
J82 with a full-length AR stably expressed by lentivirus,
luciferase activity was determined in the cell extracts with
transfection of a plasmid (MMTV-luc) containing an androgen
response element (ARE) as a reporter of AR-mediated transcriptional
activity. As shown in Fig. 1, EGF
treatment increased luciferase activity by 1.2-, 1.9- and 2.0-fold
in UMUC3 (p=0.013), 5637-AR (p=0.036) and J82-AR (p=0.050),
respectively, over mock treatment. PD168393 showing only marginal
activity (in UMUC3 and 5637-AR) or some agonist effect (1.5-fold in
J82-AR) could antagonize the EGF effect on AR transcription. In
AR-knockdown UMUC3-AR-shRNA and AR-negative lines (5637, 5637-V,
J82 and J82-V), EGF and/or PD168393 showed marginal effects on AR
transcriptional activity (data not shown). These results suggest
that EGF induces AR transactivation via EGFR in an
androgen-independent manner.
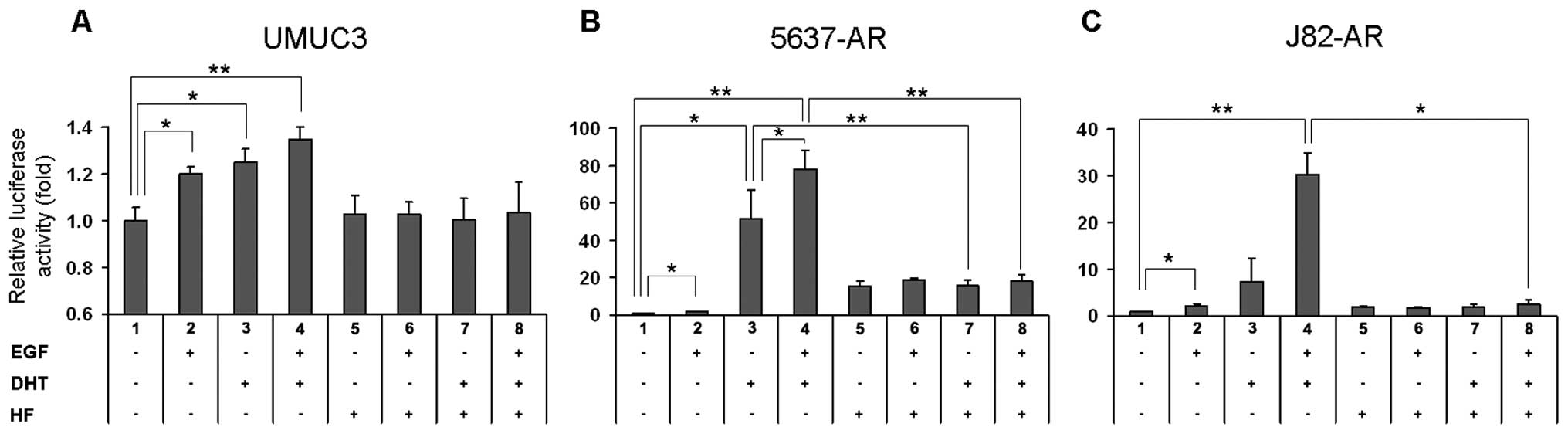
Antiandrogen blocks EGF-induced AR
transactivation
We next assessed the effect of EGF, in conjunction
with androgen and/or antiandrogen, on AR transcriptional activity
in bladder cancer cells. As shown in Fig. 2A, DHT treatment increased AR
transcription by 25% (lanes 1 vs. 3, p=0.032) and addition of EGF
further induced it by 35% (lanes 1 vs. 4, p=0.001; lanes 3 vs. 4,
p=0.103) in UMUC3. Interestingly, HF showing only marginal activity
(lanes 1 vs. 5) abolished the effects of not only DHT (lanes 3 vs.
7, p=0.077) but also EGF (lanes 2 vs. 6, p=0.061) and EGF+DHT
(lanes 4 vs. 8, p=0.082). Similarly, in 5637-AR (Fig. 2B) and J82-AR (Fig. 2C), DHT (lane 3) induced AR
transcription to 52- and 7.4-fold, respectively and EGF in the
presence of DHT (lanes 4 vs. 3) enhanced it to 78- (p= 0.035) and
30-fold (p=0.054), respectively. HF showing some agonist activities
(lanes 1 vs. 5) in 5637-AR (15-fold)/J82-AR (1.8-fold), which were
much higher (vs. 1.7-fold)/similar (vs. 2.1-fold) compared to EGF
stimulations (lane 2), could block the effects of DHT (lanes 3 vs.
7, p=0.005/p=0.164) and EGF+DHT (lanes 4 vs. 8, p=0.009/p=0.013).
Again, in UMUC3-AR-shRNA, 5637(-V) and J82(-V) cells, EGF, DHT
and/or HF showed marginal effects on AR transcription (data not
shown). These findings suggest that EGF and androgen cooperatively
induce AR transactivation that is sufficiently inhibited by an
anti-androgen.
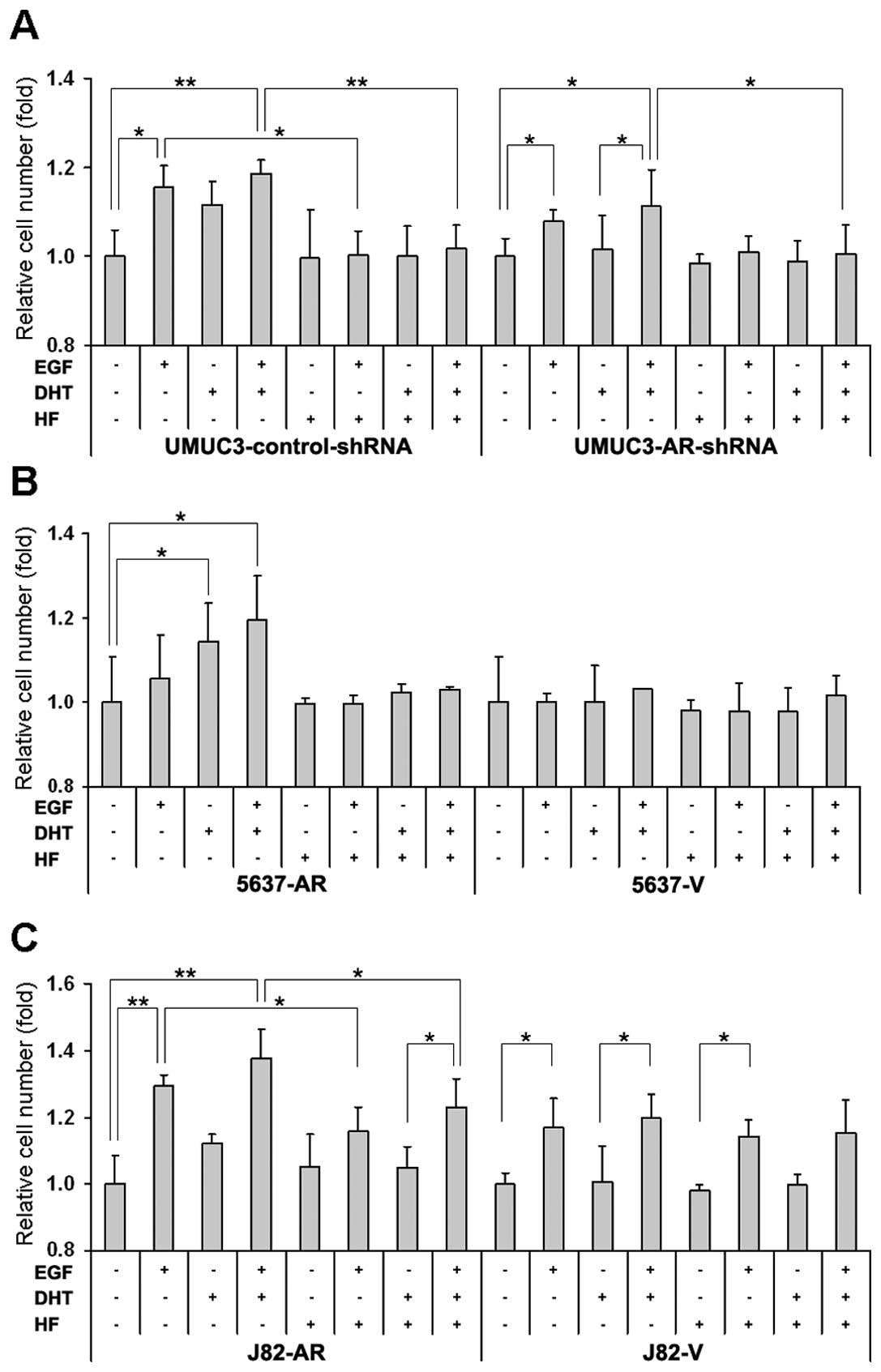
EGF stimulates cell growth via AR
signaling
We then performed the MTT assay to investigate the
effects of EGF androgen and antiandrogen on cell proliferation of
bladder cancer lines with vs. without AR (i.e., UMUC3-control-shRNA
vs. UMUC3-AR-shRNA, 5637-AR vs. 5637-V and J82-AR vs. J82-V). As
shown in Fig. 3A, in
UMUC3-control-shRNA, treatment of EGF, DHT and EGF+DHT increased
cell growth by 16% (p=0.020), 12% (p=0.195) and 19% (p=0.009),
respectively, over mock treatment and HF treatment appeared to
restore the growth to the basal levels. In UMUC3-AR-shRNA, DHT
effect was marginal (2%) and the effects of EGF (8%, p=0.039) and
EGF+DHT (11%, p=0.040) were less significant compared to those in
UMUC3-control-shRNA. In 5637-AR, treatment of EGF, DHT and EGF+DHT
induced cell growth by 6% (p= 0.558), 14% (p=0.016) and 19%
(p=0.050), respectively and HF almost completely abolished the
stimulation (Fig. 3B). In 5637-V,
only marginal effects of EGF, DHT and/or HF on cell numbers were
seen. In J82-AR, treatment of EGF, DHT and EGF+DHT induced cell
growth by 30% (p=0.001), 12% (p=0.179) and 38% (p<0.001),
respectively (Fig. 3C).
Interestingly, HF was able to antagonize the DHT effect but only
partially blocked the EGF effect. As expected, in J82-V, DHT did
not increase cell growth, while EGF and EGF+DHT, although less
significant, induced it by 17% (p=0.010) and 20% (p=0.043),
respectively. Additionally, HF failed to block the EGF effect in
J82-V cells. These results suggest that EGF promotes bladder cancer
cell proliferation at least partially through the AR pathway.
EGF increases AR and TIF2 expression
To further investigate how EGF influences AR
signals, we examined AR expression by western blotting. In UMUC3,
AR expression was increased by DHT (4.4-fold) and further enhanced
by addition of EGF (6.4-fold), whereas no significant effect of EGF
or HF was seen in the absence of DHT (Fig. 4A). HF clearly antagonized the
effects of DHT with or without EGF. In J82-AR, EGF appeared to
increase AR expression both in the presence (2.8-fold) and absence
(1.5-fold) of DHT and HF abolished these effects (Fig. 4C). In contrast, only marginal
effects of EGF and/or DHT on AR expression were observed in 5637-AR
(Fig. 4B).
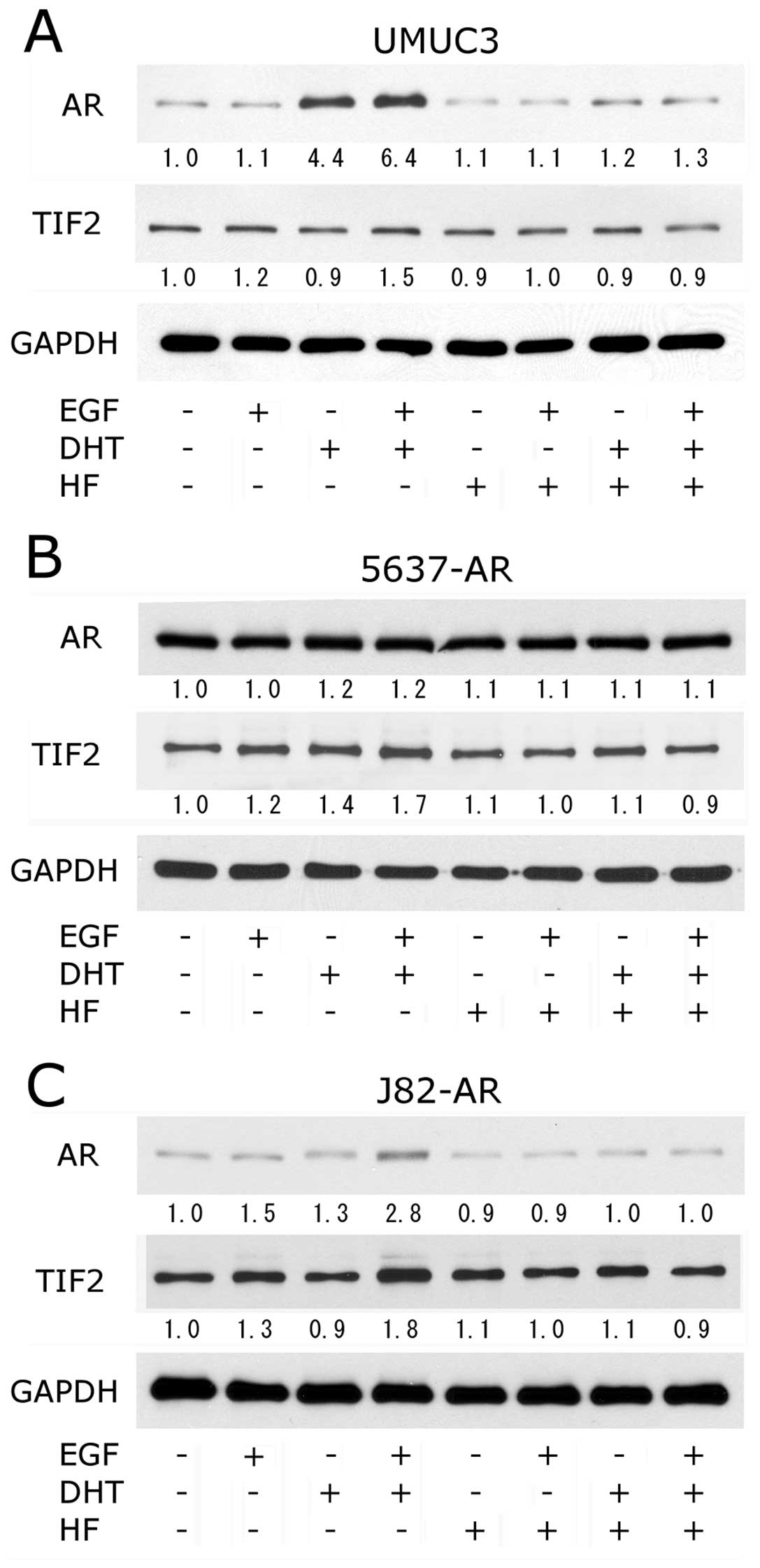 | Figure 4.Effects of EGF on AR and TIF2 protein
expression. Bladder cancer cells (A, UMUC3; B, 5637-AR; C, J82-AR)
were cultured for 24 h in the presence of ethanol (mock), 100 ng/ml
EGF, 10 nM DHT and/or 10 μM HF, as indicated. Equal amounts
of protein extracted from each cell line were immunoblotted for AR
(110 kDa, upper), TIF2 (160 kDa, middle), or GAPDH (37 kDa, lower)
as indicated. Densitometry values for specific bands standardized
by GAPDH that are relative to those of mock treatment (first lanes;
set as 1-fold) are included below the lanes. |
Because EGF was shown to induce AR transcription by
upregulating TIF2 expression in prostate cancer cells (21), we then determined the levels of
TIF2 expression in bladder cancer cell lines upon treatment with
EGF, androgen and/or antiandrogen. As shown in middle panels of
Fig. 4, EGF increased TIF2
expression in the presence (1.5- to 1.8-fold) and absence (1.2- to
1.3-fold) of DHT. DHT alone increased TIF2 expression only in
5637-AR (1.4-fold) and showed marginal effects in UMUC3 and J82-AR.
In addition, HF abrogated EGF- and/or DHT-enhanced TIF2 expression
in all these three lines.
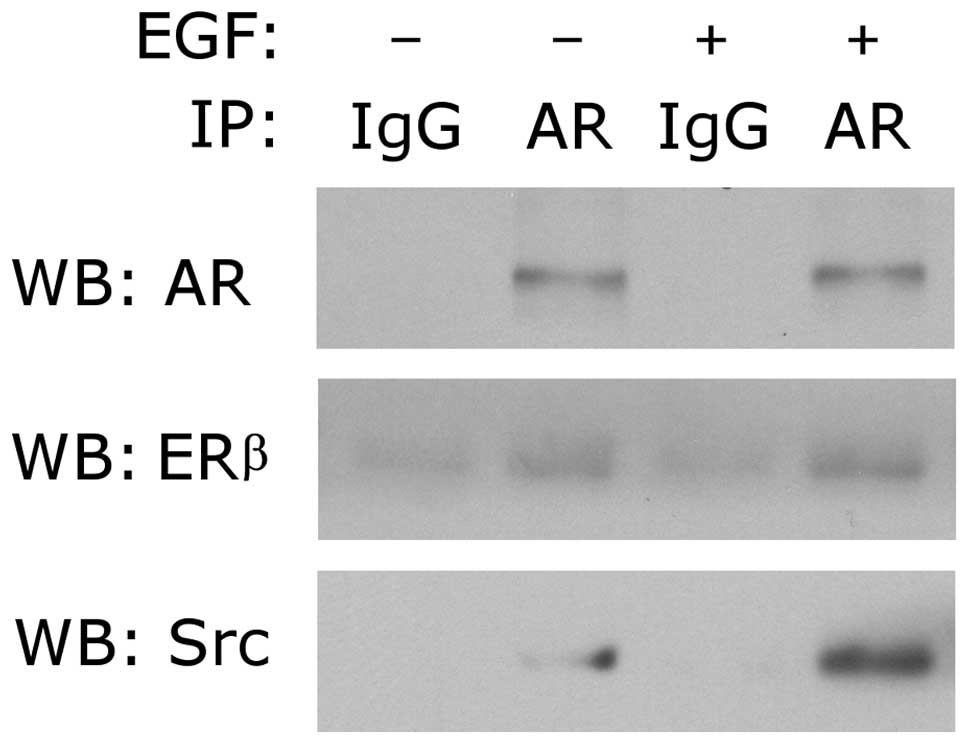
EGF induces AR association with ER and
Src
Previous studies in prostate and breast cancers
demonstrated that EGF induced AR/ER/Src association, resulting in
activation of Src signaling (25,28)
and that Src signals phosphorylated tyrosine residue of AR,
provoking its transactivation and cell proliferation (29). We therefore investigated whether
EGF induced AR/ER/Src complex formation in UMUC3 which is
ERα-negative/ERβ-positive (figure not shown). As shown in Fig. 5, both Src and ERβ were
co-immunoprecipitated with AR in bladder cancer cells. Furthermore,
EGF treatment facilitated the association of AR with ERβ or
Src.
Discussion
Dysregulation of the EGFR family is well known to
associate with bladder cancer (14–16).
AR signals have also been implicated in bladder carcinogenesis and
tumor progression (3,5,7,9–13).
Nonetheless, crosstalk between the AR and EGFR pathways remains
unclear in bladder cancer, although it has been widely studied in
prostate cancer (20–24). We have recently shown that AR
signals increase EGFR and ERBB2 expression and activity, suggesting
androgen-mediated bladder cancer progression via the regulation of
the EGFR/ERBB2 pathways (26). In
the present study, we provided evidence suggesting that EGF could
regulate cell proliferation by activating AR signals in bladder
cancer.
In prostate cancer, accumulating evidence has
indicated that EGFR/ERBB2 signals induce AR transactivation in an
androgen-dependent and -independent manner (20–22).
In bladder cancer cells, we here showed that EGF could activate AR
transcription and PD168393, a specific inhibitor of EGFR, restored
this EGF effect. These data suggest that EGF androgen-independently
induces EGFR-mediated ARE reporter activity in bladder cancer.
However, it was shown that the effect of EGF on AR transcription
might be almost negligible compared to the induction by androgens
in prostate cancer (20,21). Similarly, in bladder cancer lines
5637-AR and J82-AR where a wild-type AR was stably overexpressed,
the effect of EGF was less significant than that of DHT. On the
other hand, in UMUC3 cells that possess endogenous AR, EGF effect
(20% increase) is similar to the relatively insignificant effect of
DHT (25% increase). In addition, PD168393 displayed agonist effects
[1.5-fold (vs. 2.0-fold by EGF or 7.4-fold by DHT)] on AR
transcription in J82-AR via unknown mechanisms. It was described in
prostate cancer cells that PD168393 upregulated AR target gene
expression in the presence of androgen, possibly via blocking basal
activity of EGFR or ERBB2 (30).
Importantly, as shown in prostate cancer (21), a combination of EGF and androgen
further induced AR transcriptional activity in all the three
bladder cancer lines tested and the AR antagonist HF completely
abolished AR transactivation induced by EGF, androgen, or both at
least in UMUC3. We could not evaluate antagonistic effects of HF on
EGF-induced AR transcription due to the considerable agonist
activity of HF which was even higher than that of EGF in 5637-AR.
Thus, our results support the possibility that EGF mediates AR
transcriptional activity through the EGFR and AR pathways in
bladder cancer cells.
Consistent with previous findings shown by others
and us (5,7,9,26)
androgens promoted AR-positive bladder cancer cell proliferation
that was blocked by antiandrogens. These effects of androgens were
suggested to be at least partially mediated through the EGFR
pathway (26). In the present
study, as expected, EGF increased the growth of AR-positive cells
and, less significantly, that of AR-knockdown/negative cells. In
AR-positive lines, combined treatment with EGF and androgen further
induced cell proliferation. Of note were inhibitory effects of the
AR antagonist on EGF- and EGF+androgen-increased cell growth.
Specifically, on the growth of 5637-derived lines, EGF and/or DHT
showed only marginal effects (5637-V) and HF almost completely
abolished EGF-mediated effects (5637-AR). These findings indicate
that EGF-induced cell proliferation involves the AR pathway in
bladder cancer. Nonetheless, in J82-derived lines, EGF retained its
effect on cell growth without AR (J82-V) and HF failed to
completely inhibit EGF-increased cell proliferation (J82-AR),
suggesting the involvement of those other than the AR pathway.
It has been reported that EGF is capable of inducing
AR transcription and protein expression in androgen-independent
prostate cancer cells (21).
Others also described negative regulation of AR expression and
activity by EGFR signaling in prostate cancer (30,31).
In bladder cancer cells, we previously showed increases in the
expression of endogenous AR by androgen treatment (26), which was inconsistent with the
results demonstrated by Boorjian et al(9). We also showed no significant
increases in exogenously overexpressed AR (5637-AR) by DHT or in
endogenous and exogenous ARs by EGF (26). We confirmed our previous findings
in the three lines tested and further showed EGF-enhanced AR
overexpression in the presence of androgen in UMUC3 and J82-AR, but
not in 5637-AR. The mechanism underlying this discrepancy in the
response to the treatment of EGF+DHT between levels of exogenous AR
expression in 5637 versus J82 remains uncertain. Repeatedly, the AR
expression increased by androgen with or without EGF in bladder
cancer cells was abolished by an AR antagonist.
EGF has been shown to enhance the expression or
phosphorylation of TIF2, one of the p160 nuclear receptor
coactivators, leading to an increase in AR transactivation in
prostate cancer cells (21).
Indeed, the expression of major AR coactivators, including TIF2,
was detected in bladder cancer cell lines as well as in AR-positive
and even AR-negative bladder tumor specimens and TIF2 knockdown
resulted in a decrease in androgen-mediated cell proliferation
(9). We here found that TIF2 was
considerably (e.g., ≥1.5-fold) augmented in the presence of EGF and
DHT in bladder cancer cells, while EGF or DHT alone could lead to
marginal/only slight increases in TIF2 expression. Interestingly,
like our results in AR expression/activity and cell proliferation,
EGF-induced TIF2 upregulation was abolished by the antiandrogen.
Although detailed mechanisms need to be clarified, these results
may imply that elevated levels of TIF2 contribute to
EGF/androgen-enhanced AR trans-activation in bladder cancer
cells.
In hormone-responsive cells expressing both AR and
ER (α and/or β), such as prostate and breast cancers, AR/ER/Src
association plays a crucial role in activation of Src signals
triggered by EGF and/or sex hormones (25,28).
It was noteworthy that either AR or ER antagonist sufficiently
inhibited this EGF-mediated association and subsequent stimulatory
effects (28). It has also been
shown that Src mediates EGF-induced AR tyrosine phosphorylation in
prostate cancer cells, which leads to an increase in AR
transcriptional activity (29).
Indeed, in many bladder cancer tissue specimens, AR and ER(s) were
found to be co-expressed (3,10,12).
In this study, we showed associations of AR with ERβ and Src in
UMUC3 which were enhanced by EGF treatment. These findings suggest
that EGF activates Src via assembling the AR/ER/Src complex,
resulting in AR transactivation and cell proliferation in bladder
cancer. This may also justify the drastic inhibition of EGF-induced
effects accomplished by antiandrogen treatment.
In conclusion, EGF could increase AR transcriptional
activity and cell proliferation in bladder cancer. These EGF
effects were likely mediated through the AR pathway involving
upregulation of TIF2 expression as well as activation of Src
signals due to forming an AR/ER/Src complex. These results,
together with our previous findings, not only shed light on
crosstalk between the AR and EGFR pathways in bladder cancer but
also enhance the feasibility of androgen deprivation interfering
with this crosstalk as a potential therapeutic approach.
Abbreviations:
|
AR
|
androgen receptor;
|
|
ER
|
estrogen receptor;
|
|
EGF
|
epidermal growth factor;
|
|
EGFR
|
epidermal growth factor receptor;
|
|
FBS
|
fetal bovine serum;
|
|
DHT
|
dihydrotestosterone;
|
|
HF
|
hydroxyflutamide;
|
|
shRNA
|
short hairpin RNA;
|
|
SDS
|
sodium dodecylsulfate;
|
|
PAGE
|
polyacrylamide gel
electrophoresis;
|
|
TIF2
|
transcriptional intermediary factor
2;
|
|
ARE
|
androgen response element
|
Acknowledgements
H.M. was supported by the Department
of Defense Prostate Cancer Research Program (W81XWH-09-1-0305).
References
|
1.
|
A JemalF BrayMM CenterJ FerlayE WardD
FormanGlobal cancer statisticsCA Cancer J
Clin616990201110.3322/caac.20107
|
|
2.
|
E ScosyrevK NoyesC FengE MessingSex and
racial differences in bladder cancer presentation and mortality in
the USCancer1156874200910.1002/cncr.2398619072984
|
|
3.
|
H MiyamotoY ZhengK IzumiNuclear hormone
receptor signals as new therapeutic targets for urothelial
carcinomaCurr Cancer Drug
Targets121422201210.2174/15680091279888896522111835
|
|
4.
|
SS ShenCL SmithJ-T HsiehExpression of
estrogen receptors-α and -β in bladder cancer cell lines and human
bladder tumor tissueCancer106261026162006
|
|
5.
|
H MiyamotoZ YangY-T ChenPromotion of
bladder cancer development and progression by androgen receptor
signalsJ Natl Cancer
Inst99558568200710.1093/jnci/djk11317406000
|
|
6.
|
G SonpavdeN OkunoH WeissEfficacy of
selective estrogen receptor modulators in nude mice bearing human
transitional cell
carcinomaUrology6912211226200710.1016/j.urology.2007.02.04117572228
|
|
7.
|
AM JohnsonMJ O’ConnellH MiyamotoJ HuangJL
YaoEM MessingJE ReederAndrogenic dependence of exophytic tumor
growth in a transgenic mouse model of bladder cancer: a role for
thrombospodin-1BMC Urol87200810.1186/1471-2490-8-718433501
|
|
8.
|
J TengZ-Y WangDF JarrardDE BjorlingRoles
of estrogen receptor α and β in modulating urothelial cell
proliferationEndocr Relat Cancer153513642008
|
|
9.
|
SA BoorjianHV HeemersI FrankSA FarmerLJ
SchmidtTJ SeboDJ TindallExpression and significance of androgen
receptor coactivators in urothelial carcinoma of the bladderEndocr
Relat Cancer16123137200910.1677/ERC-08-012418845648
|
|
10.
|
C TuygunD KankayaA ImamogluA SertcelikK
ZenginM OktayN SertcelikSex-specific hormone receptors in
urothelial carcinomas of the human urinary bladder: a comparative
analysis of clinicopathological features and survival outcomes
according to receptor expressionUrol
Oncol294351201110.1016/j.urolonc.2009.01.033
|
|
11.
|
K IzumiY ZhengJ-W HsuC ChangH
MiyamotoAndrogen receptor signals regulate
UDP-glucuronosyltransferases in the urinary bladder: a potential
mechanism of androgen-induced bladder carcinogenesisMol
CarcinogenNov152011(Epub ahead of print)10.1002/mc.21833
|
|
12.
|
H MiyamotoJL YaoA ChauxExpression of
androgen and oestrogen receptors and its prognostic significance in
urothelial neoplasm of the urinary bladderBJU
Int10917161726201210.1111/j.1464-410X.2011.10706.x22221549
|
|
13.
|
Y LiK IzumiH MiyamotoThe role of the
androgen receptor in the development and progression of bladder
cancerJpn J Clin Oncol42569577201210.1093/jjco/hys07222593639
|
|
14.
|
DE NealL SharplesK SmithJ FennellyRR
HallAL HarrisThe epidermal growth factor receptor and the prognosis
of bladder
cancerCancer6516191625199010.1002/1097-0142(19900401)65:7%3C1619::AID-CNCR2820650728%3E3.0.CO;2-Q2311071
|
|
15.
|
H MiyamotoY KubotaS Noguchic-erbB-2
gene amplification as a prognostic marker in human bladder
cancerUrology556796832000
|
|
16.
|
Z LatifHER2/neu gene amplification and
protein overexpression in G3 pT2 transitional cell carcinoma of the
bladder: a role for anti-HER2 therapy?Eur J
Cancer405663200410.1016/j.ejca.2003.08.02714687790
|
|
17.
|
J BellmuntM HussainCP DinneyNovel
approaches with targeted therapies in bladder cancerCrit Rev Oncol
Hematol4685104200310.1016/S1040-8428(03)00067-2
|
|
18.
|
PC BlackPK AgarwalCPN DinneyTargeted
therapies in bladder cancer - an updateUrol
Oncol25433438200710.1016/j.urolonc.2007.05.01117826665
|
|
19.
|
NJ MacLaineMD WoodJC HolderRW ReesJ
SouthgateSensitivity of normal, paramalignant and malignant human
urothelial cells to inhibitors of the epidermal growth factor
receptor signaling pathwayMol Cancer
Res65363200810.1158/1541-7786.MCR-07-013418234962
|
|
20.
|
Z CuligA HobischMV CronauerAndrogen
receptor activation in prostatic tumor cell lines by insulin-like
growth factor-I, keratinocyte growth factor and epidermal growth
factorCancer Res54547454781994
|
|
21.
|
CW GregoryX FeiLA PongutaB HeHM BillFS
FrenchEM WilsonEpidermal growth factor increases coactivation of
the androgen receptor in recurrent prostate cancerJ Biol
Chem27971197130200410.1074/jbc.M30764920014662770
|
|
22.
|
IK MellinghoffI VivancoA KwonC TranJ
WongvipatCL SawyersHER2/neu kinase-dependent modulation of androgen
receptor function through effects on DNA binding and
stabilityCancer
Cell6517527200410.1016/j.ccr.2004.09.03115542435
|
|
23.
|
B MukherjeeD MayerDihydrotestosterone
interacts with EGFR/MAPK signaling and modulates EGFR levels in
androgen receptor-positive LNCaP prostate cancer cellsInt J
Oncol33623629200818695894
|
|
24.
|
J-C PignonB KoopmanschG NolensL DelacroixD
WaltregnyR WinklerAndrogen receptor controls EGFR and ERBB2 gene
expression at different levels in prostate cancer cell linesCancer
Res6929412949200910.1158/0008-5472.CAN-08-376019318561
|
|
25.
|
A MigliaccioG CastoriaMP GiovannelliF
AuricchioCross talk between epidermal growth factor (EGF) receptor
and extra nuclear steroid receptors in cell linesMol Cell
Endocrinol3271924201010.1016/j.mce.2010.06.01420603181
|
|
26.
|
Y ZhengK IzumiJL YaoH
MiyamotoDihydrotestosterone upregulates the expression of epidermal
growth factor receptor and ERBB2 in androgen receptor-positive
bladder cancer cellsEndocr Relat
Cancer18451464201110.1530/ERC-11-0010
|
|
27.
|
AM CanacciK IzumiY ZhengJ GordetskyJL YaoH
MiyamotoExpression of semenogelins I and II and its prognostic
significance in human prostate
cancerProstate7111081114201110.1002/pros.2132321557275
|
|
28.
|
A MigliaccioM Di DomenicoG CastoriaSteroid
receptor regulation of epidermal growth factor signaling through
Src in breast and prostate cancer cells: steroid antagonist
actionCancer Res651058510593200510.1158/0008-5472.CAN-05-0912
|
|
29.
|
Z GuoB DaiT JiangRegulation of androgen
receptor activity by tyrosine phosphorylationCancer
Cell10309319200610.1016/j.ccr.2006.08.02117045208
|
|
30.
|
C CaiDC PortnoyH WangX JiangS ChenSP
BalkAndrogen receptor expression in prostate cancer cells is
suppressed by activation of epidermal growth factor receptor and
ErbB2Cancer
Res6952025209200910.1158/0008-5472.CAN-09-002619491261
|
|
31.
|
P HenttuP VihkoGrowth factor regulation of
gene expression in the human prostatic carcinoma cell line
LNCaPCancer Res531051105819937679946
|