Introduction
Collapsin response mediator proteins (CRMPs) include
five family members (CRMP1-5) and all family members have 50–70%
sequence homology with each other (1). CRMP is a cytosolic phosphoprotein and
none of CRMP isoforms demonstrate any enzymatic activity (2,3).
Previous studies have shown that CRMP in Sema3A (collapsin-1)
signaling, mediate axonal guidance and neuronal differentiation
(3,4). The downstream effectors of CRMP
signaling is still not clear (1),
research showed CRMP mediates microtubule dynamics and endocytosis
(5–7). CRMP is mainly regulated by
semaphorins and it has been shown that CRMP also combine with a
variety of other signaling molecules (8–11)
and this imply CRMP plays a role in numerous cellular
processes.
Most studies of CRMP focus on its role in axonal
guidance in the developing nervous system and CRMP has been show
highly expressed in the developing and injured nervous system
(1), however, recent study
indicated that CRMP is involved in several malignant diseases
(12). Shih et al reported
that CRMP1 negatively correlated with lung cancer invasiveness
(13), further study indicated
LCRMP1, a long form isoform of CRMP1, promotes lung cancer cell
invasiveness and this effect may relate to LCRMP1-WAVE1 combination
and GSK-3β phosphorylation of LCRMP1 (14,15).
Both GSK-3β and WAVE-1 were involved in CRMP mediated axon
outgrowth, indicating CRMP has diverse functions in different
cellular processes. Other CRMP family members involved in cancer
include CRMP2, CRMP4 and CRMP5, all were reported associated with
different cancer cell migration, invasion, differentiation and
clinical outcome in certain tumor types (16–19).
In our previous study (20), we
first identified CRMP4 as a prostate cancer metastasis suppressor
factor by proteomics approach and verified that CRMP4 suppress
prostate cancer cell line proliferation and invasion in
vitro and the down-regulation of CRMP4 in metastatic tumor may
be due to the methylation of the CpG island within the promoter
region of the CRMP4 gene.
Cancer metastasis is the end product of tumor
genesis and development. Disseminated tumor cells arrest at target
organ though vasculature, finally forming macroscopic neoplastic
growth (21). Tumor metastasis is
a highly inefficient process, extremely small percentage of tumor
cells that enter into the systemic circulation ultimately develop
into macroscopic metastasis (22,23).
Unlike lung cancer and breast cancer, which also have a high
incidence of bone metastasis and often form osteolytic damage, most
prostate cancer formed osteoblastic bone metastasis (24). Inefficient growth at the secondary
site and high incidence of osteoblastic bone metastasis of prostate
cancer indicate the specific tumor cell-organ interactions with
host organ have a significant influence on the development of
metastasis. During the process of bone metastasis, cytokines play
an important role in the complex reciprocal interactions between
the tumor cells and the bone microenvironment, factors secreted by
tumor cells and bone stromal cells, which may contribute to the
progress of metastatic tumor growth (25).
In our present study, we sought to investigate the
role of CRMP4 in prostate cancer bone metastasis and the
relationship of CRMP4 and the cytokines and proteins which
correlate with prostate cancer bone metastasis.
Materials and methods
Cell culture
PC3 and DU145 cell lines were obtained from the ATCC
(Rockville, MD, USA) and maintained in culture PRMI-1640 medium
(Gibco, USA) supplemented with 10% fetal bovine serum (FBS, Gibco,
USA). Cells were cultured in a humidified atmosphere at 5%
CO2 and 37°C.
Lentiviral transduction
GFP-CRMP4+ lentivirus and a GFP
lentivirus (Junhui Biology, China) were transfected into PC3 and
DU145 cells in the presence of 5 μg/ml polybrene (Sigma, USA).
Transduced cells were selected by FACS (fluorescence-activated cell
sorting, BD influx, USA) by GFP expression after transduced
prostate cancer cells reached a total score of 107.
Western blot analysis, FCM (flow cytometer, BD LSRII, USA) and
quantitative real-time PCR (qRT-PCR) were used to determine the
effects of CRMP4 gene overexpression.
In vivo tumor models (animal
experiments)
The 4–5-week old male athymic nude mice (Vital
River®, Beijing, China) were studied to evaluate the
in vivo metastatic behavior of tumor cells. All the
experiments were approved by the Institutional Animal Care and Use
Committee (approval no: XYXK-2012-0081) of Sun Yat-sen
University.
Orthotopic implantation
Two groups of 7 animals each were used. Mice were
anesthetized with xylazine (FaMu Chemical Plant, Nanjing, China)
and ketamine (Fujian GuTian Pharma Co., China) though
intraperitoneal injection. A lower middle abdominal incision was
made and seminal vesicle and bladder were exposed to identify the
mouse prostate. CRMP4+PC3 cells (104/10 μl)
(CRMP4 overexpressing PC3 cells) and control PC3 were injected into
the capsule of the prostate of the animals. The incision in the
abdominal wall was closed with a 4-0 surgical suture. Baytril (2.5
μg) (Bayer, Germany) was subcutaneously injected three days after
operation to prevent infection. Small animal micro-PET/CT scan
(Inveon, Siemens, Germany) was used to detect the skeletal
metastasis 40 days after infection. fluorine-18 fluorodeoxyglucose
(18F-FDG) was used as a probe, 70 μCi 18F-FDG
for each mouse and injected intravenously in conscious animals via
the tail vein. Twenty-five minutes later, the mice were
anesthetized and the scanning was performed 30 min after injection.
The animals were placed on a heating pad to maintain body
temperature throughout the procedure and visually monitored for
breathing and any other signs of distress throughout the entire
imaging period. Imagines were analysis by Inevon Research Workplace
4.1 software (Siemens, Germany). After the examination the animals
were sacrificed and dissected to identify metastasis tumor tissue.
Suspicious bone metastatic was fixed in 10% formalin for
histological examination.
Intracardiac injection
Two groups of 7 animals each were used, The
CRMP4+PC3 and PC3 cells were injected into the left
ventricle at a concentration of 105/50 μl of
phosphate-buffered saline (PBS). Injection methods as previously
described (26). Small animal
micro-PET/CT scan and tissue processing was as indicated above.
Intratibial injection
Two groups of 7 animals each were used, the
CRMP4+PC3 and PC3 cells were injected into the left
tibia medullary cavity of the two groups at a concentration of
104/10 μl of 5 μl PBS and 5 μl Matrigel (BD, USA). A
21-G syringe was used to drill a hole though the tuberosity of
tibia and cells were injected though 29-G insulin syringe.
Injection was done very slowly to prevent cells entering the soft
tissue. No incision was made. The mice were sacrificed 40 days
after injection and the left legs were harvested, 10%
formalin-fixed and a high-resolution micro-CT (Inveon, Siemens,
Germany) were used to measure bone destruction. CT imagines were
scored as follows: 0, normal; 1, lytic lesion present within the
medullary canal only; 2, obliteration of one cortex; 3,
obliteration of two cortices. Two experienced orthopedists in a
blinded manner identified the scores.
Quantitative real-time PCR (qRT-PCR)
analysis
The total RNA was isolated using TRIzol Reagent
(Invitrogen, USA), cDNA synthesized with a Revert Aid first-strand
cDNA synthesis kit (Takara, China). SYBR Premix Ex Taq (Life
Technologies, USA) was used for qRT-PCR. Primers are listed in
Table I. qRT-PCR analysis was
performed on ABI 7500 Sequence Detection System (Applied
Biosystems, USA). Gene expression relative to housekeeping gene
GAPDH was calculated using the 2−ΔΔCt method.
2−ΔΔCt >2 or <1/2 was considered statistically
significant.
 | Table IPrimer sequences. |
Table I
Primer sequences.
| Gene | Forward primer | Reverse primer |
|---|
| Noggin |
CCATGCCGAGCGAGATCAAA |
TCGGAAATGATGGGGTACTGG |
| BMP2 |
TGCGTACTCACGGTGGAATC |
GTAAAACCCGTCTGTAGCTTCTT |
| BMP4 |
ATGATTCCTGGTAACCGAATGC |
CCCCGTCTCAGGTATCAAACT |
| BMP6 |
AGCGACACCACAAAGAGTTCA |
GCTGATGCTCCTGTAAGACTTGA |
| BMP7 |
TCGGCACCCATGTTCATGC |
GAGGAAATGGCTATCTTGCAGG |
| Sema3A |
ACACCAGAAGAGATGAATGC |
GCGTACAAGTGAGTCTGATT |
| NRP1 |
GGCGCTTTTCGCAACGATAAA |
TCGCATTTTTCACTTGGGTGAT |
| NRP2 |
GCTGGCTATA-TCACCTCTCCC |
TCTCGATTTCAAAGTGAGGGTTG |
| VEGFA |
CTTTTCTCTGCCTCCACAATG |
GAGTGGTTGACCTTCCTCCA |
| VEGFB |
CCATCTCTTTTATCAGGGTTGG |
CTCTGTGCAAGTAAGCATCTTACA |
| VEGFC |
CCACGGGAGGTGTGTATAGA |
CAGGAAGTGTGATTGGCAAA |
| VEGFD |
ACTCAGTGCAGCCCTAGAGAA |
GAACACGTTCACACAAGGGG |
| OPG |
GCTAACCTCACCTTCGAG |
TGATTGGACCTGGTTACC |
| RANKL |
TTAAGCCAGTGCTTCACGGG |
ACGTAGACCACGATGATGTCGC |
| SDF-1 |
ATTCTCAACACTCCAAACTGTGC |
ACTTTAGCTTCGGGTCAATGC |
| CXCR4 |
ACTACACCGAGGAAATGGGCT |
CCCACAATGCCAGTTAAGAAGA |
Western blot analysis
Total cellular proteins were extracted with RIPA
lysis buffer kit (KeyGen Biotech, China). Samples were separated by
10% sodium dodecylsulfate polyacrylamide gel electrophoresis
(SDS-PAGE) and then transferred to a polyvinylidene fluoride (PVDF)
membrane. The CRMP4 and Noggin was assessed using primary rabbit
anti-CRMP4 and rabbit anti-Noggin monoclonal antibody (Abcam, USA).
NRP1 was assessed using primary rabbit anti-NRP1 monoclonal
antibody (ABclonal, Cambridge). The membranes were incubated with
primary antibodies and then horseradish peroxidase (HRP) secondary
antibodies. An enhanced chemiluminescence reagent (ECL) kit
(Millipore, USA) was used to detect the labeled proteins. GAPDH
(rabbit anti-GAPDH monoclonal antibody, Jetway, China) was used to
control equal loading quantity.
Histological experiments
Tissue specimens were decalcificated in 14% EDTA and
embedded in paraffin, 5-μm sections of tissue were used.
Hematoxylin and eosin (H&E) staining was used to define tumor
tissues of the intracardiac injection and orthotopic implantation
models. H&E staining, tartrate-resistant acid phosphatase
(TRAP) staining (Sigma, USA) and immunohistochemical (IHC) staining
were applied on the left tibia tissue of the intratibial injection
models. Osteoclast number was assessed as TRAP-positive cells along
the tumor-bone interface on TRAP-stained sections, and expressed as
the osteoblast number per ×200 field. For IHC staining, antigen
retrieval was carried out by pepsin antigen retrieval solutions
(ZSGB-Bio, China). Primary rabbit anti-Noggin monoclonal antibody
(Abcam) and primary rabbit anti-NRP1 monoclonal antibody (ABclonal,
USA) were used, respectively. The images were observed under a
fluorescence microscope (Leica DM 400B, Germany) and analyzed by
Leica Application suite Version 3.8.0 software (Leica, Germany).
The IHC result was classified as 0 [equal to negative (−)]; 1–4
[weakly positive (+)]; 5–8 [moderately positive (++)] and 9–12
[strongly positive (+++)]. Two experienced pathologists in a
blinded manner identified the scores.
Statistical analysis
Osteoclast data are expressed as the mean ± SD.
Statistical significance was determined by the unpaired Student’s
t-test. CT score and IHC score were analyzed using Kruskal-Wallis
test. Statistical analysis was performed using SPSS 13.0 software.
The results with P-values <0.05 were considered as statistically
significant, and indicated by an asterisk in the figures.
Results
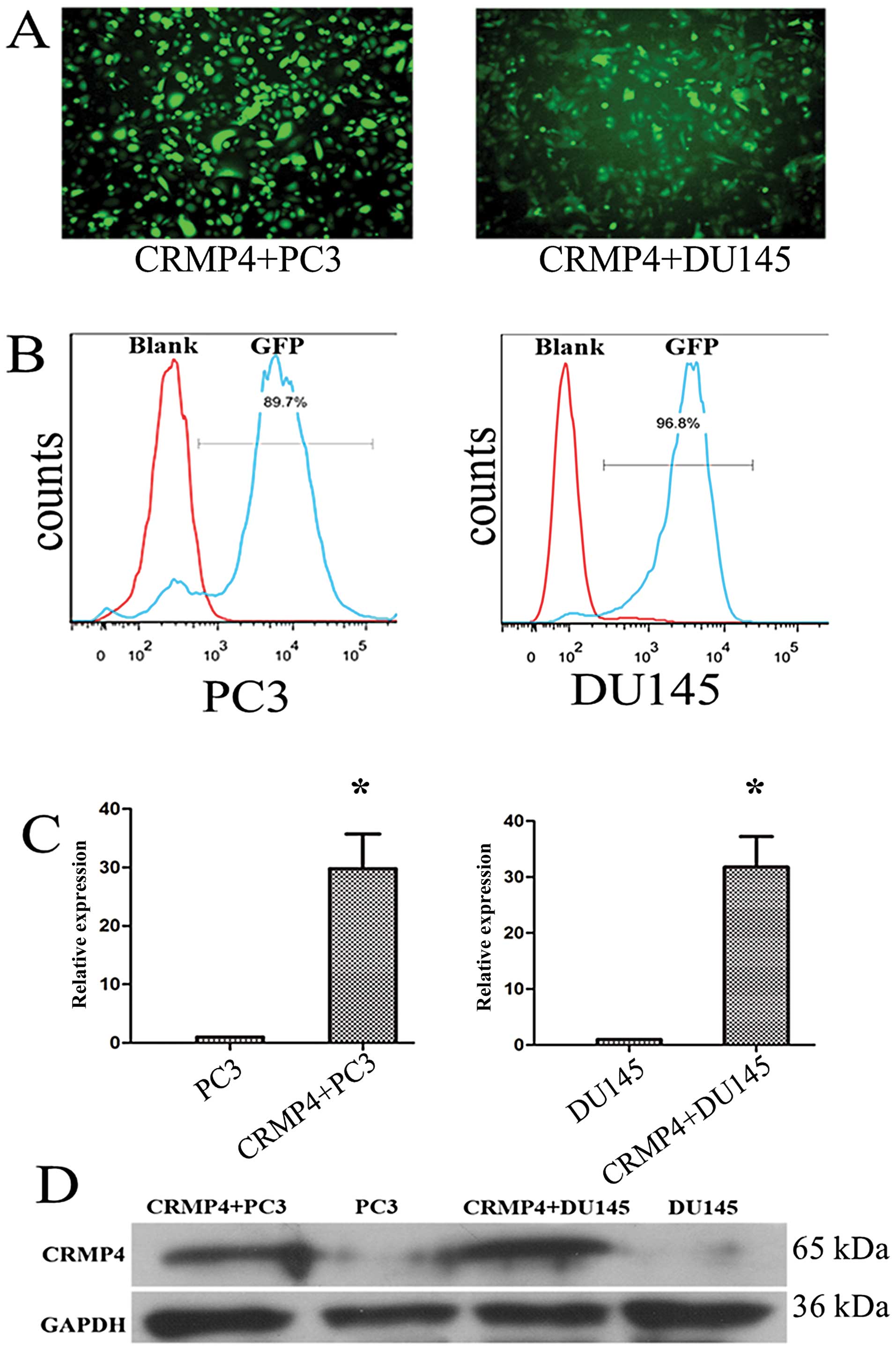
Construction of the stable CRMP4 gene
overexpressing PC3 and DU145 cell lines
After transduction of the CRMP4+
lentivirus of PC3 and DU145 cell lines, FACS was used for a stable
overexpressing CRMP4 PC3 and DU145 cell lines. We obtained 30-fold
overexpression of CRMP4 at mRNA level by qRT-PCR examination and
obvious differential expression at protein level by western blot
analysis compared to control PC3 and DU145 cells (Fig. 1).
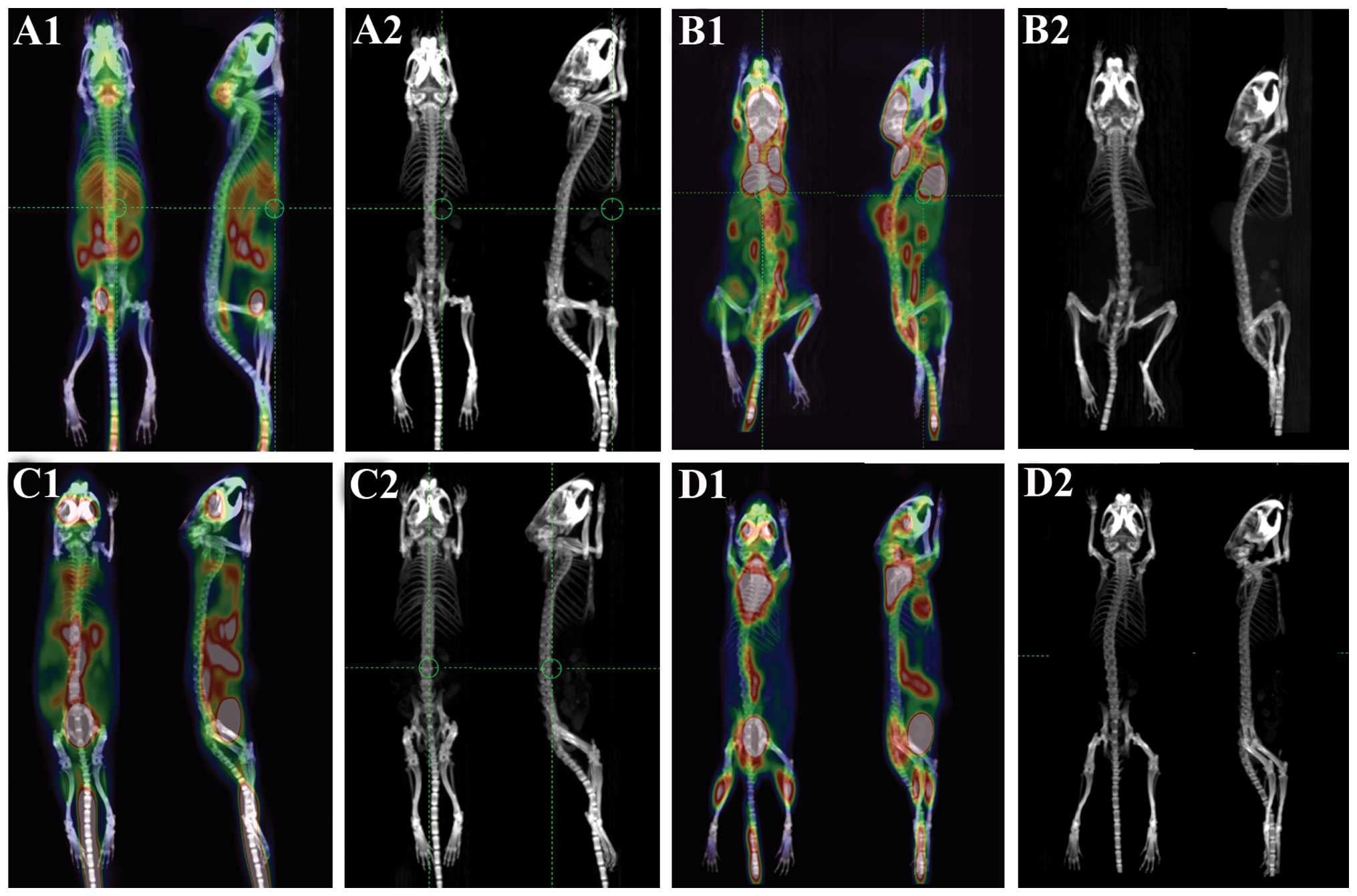
CRMP4 overexpression shows no difference
of bone metastasis capacity in orthotopic implantation and
intracardiac injection models
PET/CT scanning of two groups of models showed no
obvious bone destruction. In the orthotopic group, PET/CT
examination showed abdominal high signal, mandible and abdominal
wall high signal were occasionally observed (3/14), but both
CRMP4+ and control groups showed no bone high signal, CT
scanning showed skeleton system undamaged (Fig. 2A and C). In the intracardiac
injection groups, soft tissue metastasis (subcutaneous and muscle
tissue) is the most common metastasis which was 3/7 in the
CRMP4+ group and 7/7 in the control group. Lung
metastasis (1/7 CRMP4+ group and 5/7 control group) is
the second most common metastasis and the other metastasis included
liver metastasis (2/7 control group). Some of the animals showed
pelvis and spine abnormal signal but no bone destruction in CT
scanning and no abnormal bone tissue was identified in anatomy and
H&E staining (Fig. 2B and
D).
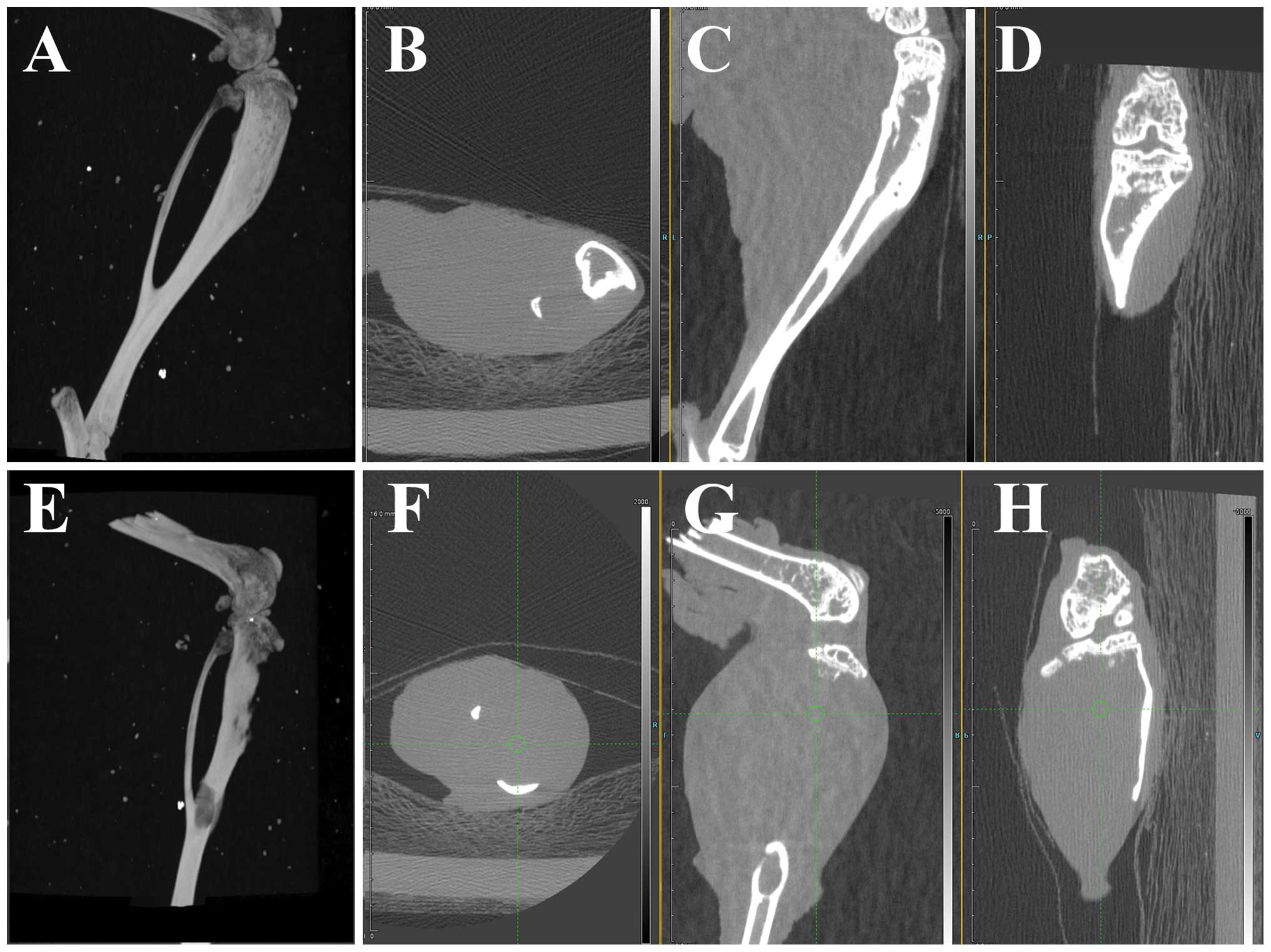
CRMP4 overexpression inhibits tumor
growth in intratibial injection models
High resolution CT scanning of left limb of
intratibial injection models showed that both groups developed
osteolytic bone destruction. In the CRMP4+ group, three
dimensional reconstructive CT imaging showed integrated tibia form
and sagittal and coronal view showed an osteoclastic reaction in
medullary cavity (Fig. 3A–D).
Internal wall of cortical bone decay was common (7/7) and
micro-cortical perforation was observed (3/7). Two mice developed
fracture but showed no severe bone destruction. In the control
group, reconstructive CT images showed apparent bone resorption,
cortex destruction and fracture was common (4/7) after 40 days of
injection (Fig. 3E–H). CT imaging
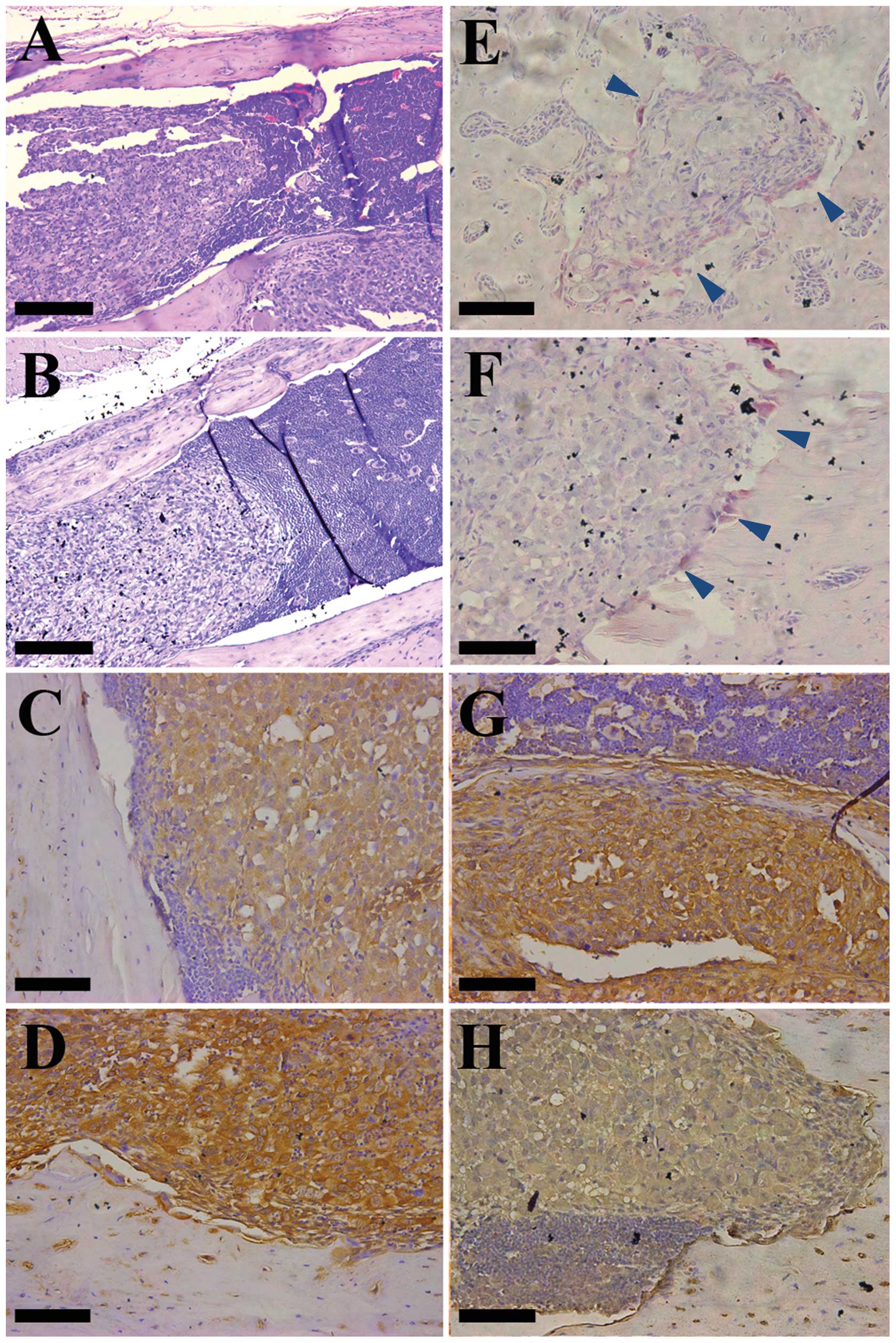
score showed statistical difference (P<0.05). H&E staining
showed large and irregular tumor cells with a light stain and a big
nucleus, compare to a small and round deep stain in normal bone
marrow cells. In the CRMP4+ group, tumor cell growth was
restricted to the marrow cavity. The boundary between cancer cells
and bone marrow cells is clear. In the control group, bone cortex
destruction is common and in some cases bone marrow cells
disappeared completely. Both groups showed cancer cells mixed with
smaller bone marrow cells and both group had osteolytic bone
destruction (Fig. 5A and B). TRAP
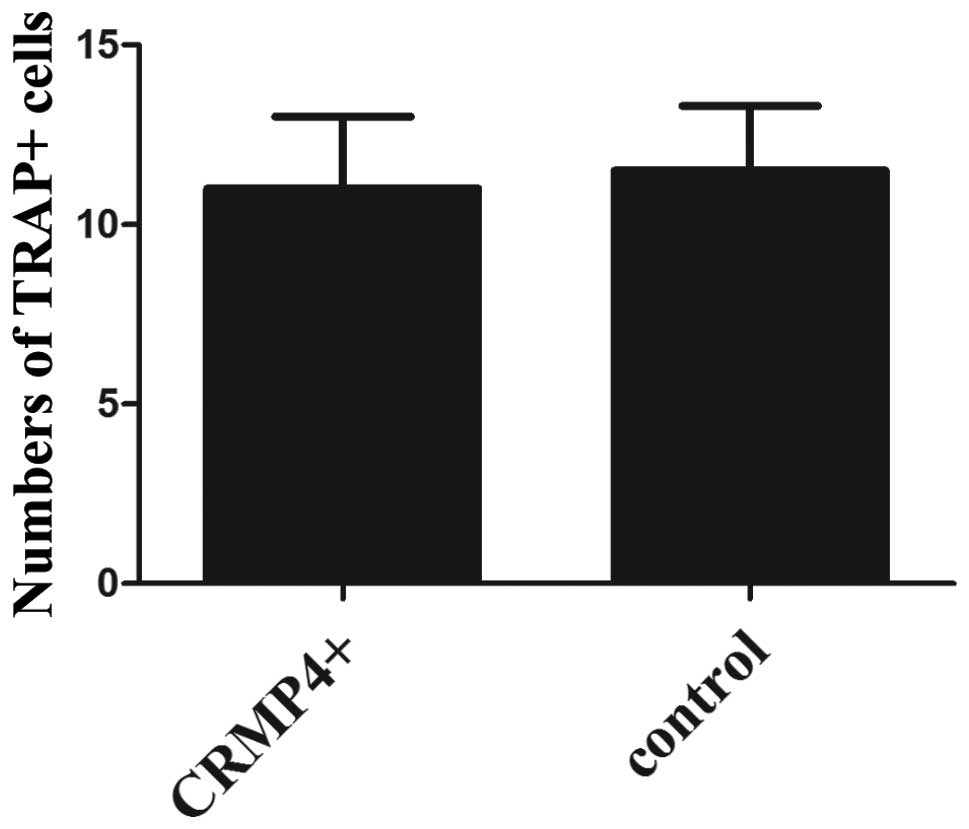
staining showed TRAP-positive cells (osteoclast) located at the
tumor-bone matrix interface, and no statistical difference of
positive cells between the two groups (Figs. 5E and F and 6). IHC staining showed CRMP4+
tumor cells overexpressed Noggin and low expression of NRP1
(P<0.05), conforming with the in vitro experiments
(Fig. 5C, D, G and H).
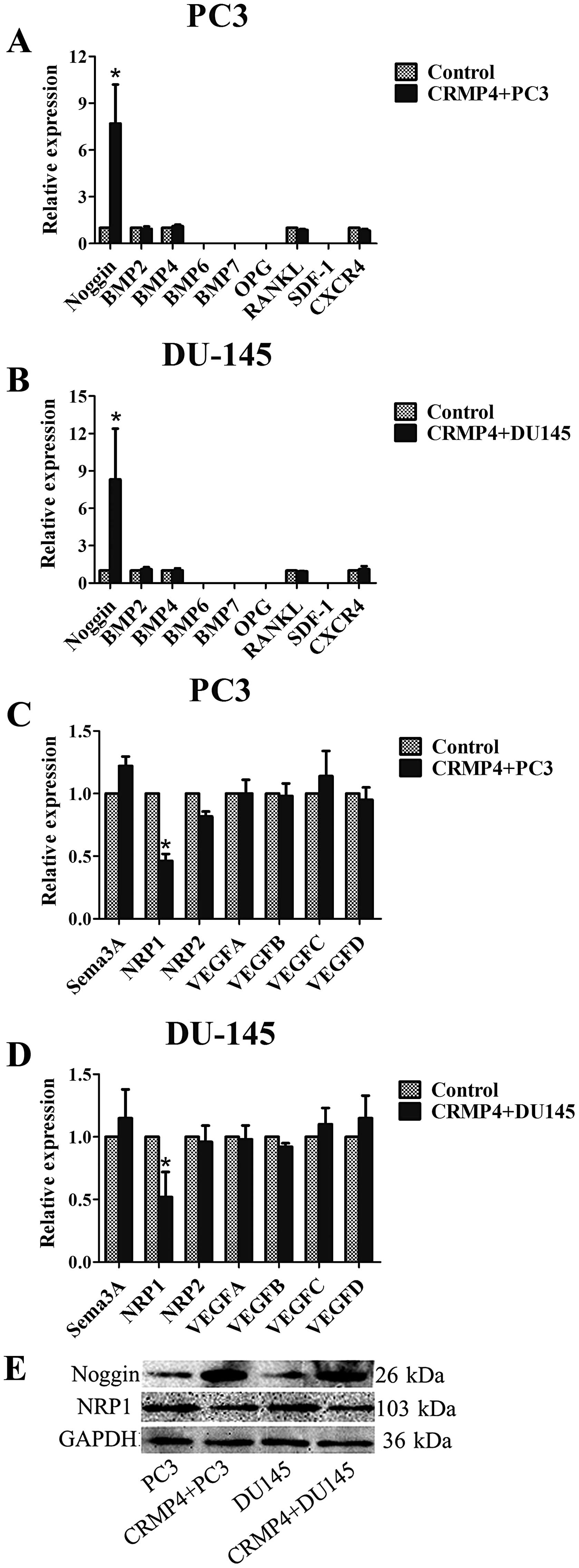
CRMP4 overexpression enhance Noggin
expression in vitro
Noggin/BMP signaling was reported involved in
prostate cancer bone metastasis (27), both Noggin/BMP and CRMP play an
important role in the developing nervous system (28). We hypothesized CRMP4 combined in
the Noggin/BMP signaling in prostate cancer in the signal pathway.
qRT-PCR examination showed both PC3 and DU145 cell line expressed
low level of Noggin, BMP2 and BMP4, the cell lines expressed no
BMP6 and BMP7 (no CT value detected). CRMP4+PC3 and
DU145 cell lines overexpress Noggin both in mRNA and protein
levels. No significant difference was recorded in the RNA
expression of BMP2 and BMP4 in the cell lines (Fig. 4A, B and E).
CRMP4 overexpression inhibits NRP1
expression
CRMP4 is a downstream protein in Sema3A-NRP1
signaling and NRP1 is an important negative regulation
transmembranous protein in this signaling (1), NRP1 is a coreceptor for VEGF and
promotes VEGFR activation (29,30).
We hypothesized that CRMP4 inhibits VEGF combined to its receptor
by inhibiting NRP1. qRT-PCR examination showed both PC3 and DU145
cell lines highly express all four VEGF members and NRP1, with low
expression of NRP2 and Sema3A. CRMP4 overexpression reduced NRP1
expression, but had no effect on sema3A, NRP2 and VEGF expression.
Western blot examination was confirmed with qRT-PCR examination,
NRP1 downregulation was observed in both CRMP4+ cell
lines (Fig. 4C–E).
CRMP4 and other cytokines involved in
prostate bone metastasis
SDF-1/CXCR4 pathway is an element in the processes
of hematopoietic cell homing to bone during embryonic development
(31) and related to cancer bone
metastasis (32,33). In our research, we find that both
PC3 and DU145 express no SDF-1 gene and low expression of CXCR4
mRNA, and CRMP4 over-expression did not affect CXCR4 mRNA
expression. OPG/ RANKL signaling is an important regulator of
osteolytic/ osteoblastic balance in bone (34) and it has been shown that OPG and
RANKL are key factors mediating osteolytic bone injury in prostate
cancer bone metastasis (35). Our
research showed both cell lines express OPG and RANKL at a low mRNA
lever but these factors are not affected by CRMP4 overexpression
(Fig. 4A and B).
Discussion
Cancer bone metastasis is a complex biological
process involved in numerous cell and protein interactions. CRMP is
a new cancer related protein, understanding its role in cancer
process and the relationship with other cancer related proteins is
likely to improve our understanding of cancer physiology and
pathology.
In our research, orthotopic implantation and
intracardiac injection in nude mouse models showed low rate of bone
metastasis. In agreement, previous research showed most immune
deficiency mouse implantation models revealed low bone metastatic
capability (36). Yang et
al (37) reported a prostate
orthotopic implantation model by a GFP-PC3 cell line and
micrometastasis were identified in the skeleton including the
skull, rib, pelvis, femur, and tibia. Corey et al (38) reported a prostate orthotopic
implantation model with PSA+ LuCaP 23.8 and LuCaP 35
cells and dectected PSA expression in bone marrow by reverse
transcription PCR (RT-PCR) examination, which indicates
mircrometastasis formed in the bone. Our experiment showed low
skeleton metastasis by 18F-FDG probe PET/CT examination
in prostate orthotopic implantation (0/14) and intracardiac
injection models (0/14). All the animals formed apparent
subcutaneous or abdominal tumors 40 days after implantation but
PET/CT showed no obvious bone abnormal signal and the CT scan
showed no bone destruction. This result may imply micrometastatic
cancer cell dormancy in a low metabolic rate can not be detected by
18F-FDG examination, this result imply tumor cell
dormancy in bone mircometastasis and the animal probably die of
other metastasis before dormant bone metastatic tumor recur and
form a macroscopic metastasis. Intratibial injection model produce
stable tumor growth in bone compared to intracardiac injection and
orthotopic implantation, this method to some degree is incorrectly
referred to as the metastasis model. It failed to reveal the early
stage of tumor metastasis, but this method offer a stable condition
of tumor cell growth in the bone environment. In the present study,
the tumor growth is suppressed by CRMP4 overexpression in bone
environment, but whether CRMP4 affects the early stage of prostate
cancer bone metastasis need to be further investigated.
Noggin is a bone morphogenetic protein (BMP)
antagonist and is an essential regulator of BMP activity (39). Both Noggin-BMP and Sema-CRMP play
important roles in the early stage of the nervous system
development. Noggin/BMPs and the relationship with CRMP were
reported in a developing Xenopus nervous system. Kamata
et al (40) reported CRMP-2
expression was activated by Noggin and dominant negative BMP
receptor in Xenopus embryos. The authors considered that the
transcriptional control of XCRMP-2 gene is one of the targets of
BMP4 signaling. In prostate cancer, Noggin was demonstrated as a
potent protector in bone metastasis (41,42)
and BMP family, which include more than a dozen members,
demonstrated to have different function in numerous studies
(27,42,43).
We believe Noggin overexpression may suppress the cell interaction
between tumor cells and BMPs in bone matrix, but the specific
mechanism is not clear. Based on the reported function of BMP and
CRMP in neural development, our results suggest a feedback
regulation may exist between the CRMP, Noggin and BMP in prostate
cancer.
NRP1 is an important transmembranous co-receptor for
VEGF and semaphorin family member. NRP1 acts as a co-receptor for
VEGF and promotes VEGFR activation (44), VEGF bind to VEGFR to activate
downstream signaling including PI3K, ERK and MAPK pathway,
consequently promoting tumor cells proliferation, survival,
differentiation, migration and angiogenesis (30). It was shown that Sema3 competes
with VEGF for NRP1 binding and prevents angiogenesis (45,46).
CRMP4 overexpression and NRP1 down-regulation may suppress VEGF in
autocrine and paracrine manner by both tumor cells and bone marrow
cells, binding to tumor cells VEGFR blocks the downstream pathway.
NRP1 downregulation may derive from the activation of Sema3A
signaling in tumor cells since most of Sema family proteins are
expressed in prostate cancer cells, and further research is needed
on this aspect.
OPG/RANKL signaling is a key regulator for
osteoclast/ osteoblast balance (34). RANKL binds to its receptor RANK to
control osteoclast differentiation, activation and survival.
Osteoprotegerin (OPG) blocks ligand binding to RANK, thereby
preventing osteoclast differentiation and activation. It has been
shown (47,48) that OPG plays protective role in
cancer bone metastasis and RANKL promotes osteolytic bone
metastasis. In our investigation, CRMP4 overexpression showed no
influence on RANKL and OPG expression and the in vivo study
showed both groups developed osteolytic bone destruction and no
osteoclast cell difference in TRAP staining, indicating CRMP4
overexpression may have no effect on osteoclast/osteoblast balance
in bone microenvironment. SDF1/CXCR4 which is reported to promote
prostate spread to the bone (32,33),
is not affected by CRMP4 overexpression, indicating CRMP4 may not
be involved in the early stage of bone metastasis.
In this study, we sought to determine the role of
CRMP4 in prostate cancer bone metastasis and the relationship
between CRMP4 and cytokines related with bone metastasis. The
results from this study demonstrated that CRMP4 inhibits nude mouse
tumor growth in a nude intratibial injection model and Noggin and
NRP1 may act as downstream cytokines of CRMP4 to mediate the tumor
cells in bone environment reaction. We believe other pathways may
also exist in CRMP4 regulating NRP1 and Noggin expression in
prostate cancer and we will further investigate the downstream
biological effects of CRMP4. In conclusion, our research suggested
CRMP4 as a potential target for prevention of prostate bone
metastasis.
Acknowledgements
This study was supported by the Natural Science
Foundation of China (31170947), the China Postdoctoral Science
Foundation (2014M552272), the Guangdong Natural Sciences Foundation
of China (S2012020011099). We sincerely thank Tie Niu, Nan Cai,
Changchang Jia (Biotherapy Center, The Third Affiliated Hospital of
Sun Yat-sen University) for their help in this study.
References
|
1
|
Schmidt EF and Strittmatter SM: The CRMP
family of proteins and their role in Sema3A signaling. Adv Exp Med
Biol. 600:1–11. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wang LH and Strittmatter SM: Brain CRMP
forms heterotetramers similar to liver dihydropyrimidinase. J
Neurochem. 69:2261–2269. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Deo RC, Schmidt EF, Elhabazi A, et al:
Structural bases for CRMP function in plexin-dependent semaphorin3A
signaling. EMBO J. 23:9–22. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Goshima Y, Nakamura F, Strittmatter P, et
al: Collapsin-induced growth cone collapse mediated by an
intracellular protein related to UNC-33. Nature. 376:509–514. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Fukata Y, Itoh TJ, Kimura T, et al: CRMP-2
binds to tubulin heterodimers to promote microtubule assembly. Nat
Cell Biol. 4:583–591. 2002.PubMed/NCBI
|
|
6
|
Gu Y and Ihara Y: Evidence that collapsin
response mediator protein-2 is involved in the dynamics of
microtubules. J Biol Chem. 275:17917–17920. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yuasa-Kawada J, Suzuki R, Kano F, et al:
Axonal morphogenesis controlled by antagonistic roles of two CRMP
subtypes in microtubule organization. Eur J Neurosci. 17:2329–2343.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Arimura N, Inagaki N, Chihara K, et al:
Phosphorylation of collapsin response mediator protein-2 by
Rho-kinase. Evidence for two separate signaling pathways for growth
cone collapse. J Biol Chem. 275:23973–23980. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kawano Y, Yoshimura T, Tsuboi D, et al:
CRMP-2 is involved in kinesin-1-dependent transport of the
Sra-1/WAVE1 complex and axon formation. Mol Cell Biol.
25:9920–9935. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Santolini E, Puri C, Salcini AE, et al:
Numb is an endocytic protein. J Cell Biol. 151:1345–1352. 2000.
View Article : Google Scholar
|
|
11
|
Nishimura T, Fukata Y, Kato K, et al:
CRMP-2 regulates polarized Numb-mediated endocytosis for axon
growth. Nat Cell Biol. 5:819–826. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Tan F, Thiele CJ and Li Z: Collapsin
response mediator proteins: potential diagnostic and prognostic
biomarkers in cancers. Oncol Lett. 7:1333–1340. 2014.PubMed/NCBI
|
|
13
|
Shih JY, Yang SC, Hong TM, et al:
Collapsin response mediator protein-1 and the invasion and
metastasis of cancer cells. J Natl Cancer Inst. 93:1392–1400. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Pan SH, Chao YC, Hung PF, et al: The
ability of LCRMP-1 to promote cancer invasion by enhancing
filopodia formation is antagonized by CRMP-1. J Clin Invest.
121:3189–3205. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang WL, Hong TM, Chang YL, et al:
Phosphorylation of LCRMP-1 by GSK3beta promotes filopodia
formation, migration and invasion abilities in lung cancer cells.
PLoS One. 7:e316892012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wu CC, Chen HC, Chen SJ, et al:
Identification of collapsing response mediator protein-2 as a
potential marker of colorectal carcinoma by comparative analysis of
cancer cell secretomes. Proteomics. 8:316–332. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Oliemuller E, Peláez R, Garasa S, et al:
Phosphorylated tubulin adaptor protein CRMP-2 as prognostic marker
and candidate therapeutic target for NSCLC. Int J Cancer.
132:1986–1995. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Shimada K, Ishikawa T, Nakamura F, et al:
Collapsin response mediator protein 2 is involved in regulating
breast cancer progression. Breast Cancer. Feb 5–2013.(Epub ahead of
print).
|
|
19
|
Meyronet D, Massoma P, Thivolet F, et al:
Extensive expression of collapsin response mediator protein 5
(CRMP5) is a specific marker of high-grade lung neuroendocrine
carcinoma. Am J Surg Pathol. 32:1699–1708. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gao X, Pang J, Li LY, et al: Expression
profiling identifies new function of collapsin response mediator
protein 4 as a metastasis-suppressor in prostate cancer. Oncogene.
29:4555–4566. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Valastyan S and Weinberg RA: Tumor
metastasis: molecular insights and evolving paradigms. Cell.
147:275–292. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chambers AF, Groom AC and MacDonald IC:
Dissemination and growth of cancer cells in metastatic sites. Nat
Rev Cancer. 2:563–572. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
23
|
Nagrath S, Sequist LV, Maheswaran S, et
al: Isolation of rare circulating tumour cells in cancer patients
by microchip technology. Nature. 450:1235–1239. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Choueiri MB, Tu SM, Yu-Lee LY, et al: The
central role of osteoblasts in the metastasis of prostate cancer.
Cancer Metastasis Rev. 25:601–609. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chung LW: Prostate carcinoma bone-stroma
interaction and its biologic and therapeutic implications. Cancer.
97:772–778. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wu TT, Sikes RA, Cui Q, et al:
Establishing human prostate cancer cell xenografts in bone:
induction of osteoblastic reaction by prostate-specific
antigen-producing tumors in athymic and SCID/bg mice using LNCaP
and lineage-derived metastatic sublines. Int J Cancer. 77:887–894.
1998. View Article : Google Scholar
|
|
27
|
Yang S, Zhong C, Frenkel B, et al: Diverse
biological effect and smad signaling of bone morphogenetic protein
7 in prostate tumor cells. Cancer Res. 65:5769–5777. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Pera EM, Acosta H, Gouignard N, et al:
Active signals, gradient formation and regional specificity in
neural induction. Exp Cell Res. 321:25–31. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ferrara N, Gerber HP and LeCouter J: The
biology of VEGF and its receptors. Nat Med. 9:669–676. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Takahashi H and Shibuya M: The vascular
endothelial growth factor (VEGF)/VEGF receptor system and its role
under physiological and pathological conditions. Clin Sci.
109:227–241. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hardy CL: The homing of hematopoietic stem
cells to the marrow. Am J Med Sci. 309:260–266. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Taichman RS, Cooper C, Keller ET, et al:
Use of the stromal cell-derived factor-1/cxcr4 pathway in prostate
cancer metastasis to bone. Cancer Res. 62:1832–1837.
2002.PubMed/NCBI
|
|
33
|
Zhang XH, Wang Q, Gerald W, et al: Latent
bone metastasis in breast cancer tied to Src-dependent survival
signals. Cancer Cell. 16:67–78. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Hsu H, Lacey DL, Dunstan CR, et al: Tumor
necrosis factor receptor family member RANK mediates osteoclast
differentiation and activation induced by osteoprotegerin ligand.
Proc Natl Acad Sci USA. 96:3540–3545. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Virk MS, Petrigliano FA and Liu NQ:
Influence of simultaneous targeting of the bone morphogenetic
protein pathway and RANK-RANKL axis in osteolytic prostate cancer
lesion in bone. Bone. 44:160–167. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Singh AS and Figg WD: In vivo models of
prostate cancer metastasis to bone. J Urol. 174:820–826. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Yang M, Jiang P and Sun FX: A fluorescent
orthotopic bone metastasis model of human prostate cancer. Cancer
Res. 59:781–786. 1999.PubMed/NCBI
|
|
38
|
Corey E, Quinn JE, Vessella RL, et al: A
novel method of generating prostate cancer metastases from
orthotopic implants. Prostate. 56:110–114. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Krause C, Guzman A and Knaus P: Noggin.
Int J Biochem Cell Biol. 43:478–481. 2011. View Article : Google Scholar
|
|
40
|
Kamata T, Daar IO, Subleski M, et al:
Xenopus CRMP-2 is an early response gene to neural induction. Brain
Res Mol Brain Res. 57:201–210. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Feeley BT, Krenek L, Liu N, et al:
Overexpression of noggin inhibits BMP-mediated growth of osteolytic
prostate cancer lesions. Bone. 38:154–166. 2006. View Article : Google Scholar
|
|
42
|
Morrissey C, Brown LG, Pitts TE, et al:
Bone morphogenetic protein 7 is expressed in prostate cancer
metastases and its effects on prostate tumor cells depend on cell
phenotype and the tumor microenvironment. Neoplasia. 12:192–205.
2010.
|
|
43
|
Dai J, Keller J, Zhang J, et al: Bone
morphogenetic protein-6 promotes osteoblastic prostate cancer bone
metastases through adual mechanism. Cancer Res. 65:8274–8285. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Jackson MW: A potential autocrine role for
vascular endothelial growth factor in prostate cancer. Cancer Res.
62:854–859. 2002.PubMed/NCBI
|
|
45
|
Staton CA: Class 3 semaphorins and their
receptors in physiological and pathological angiogenesis. Biochem
Soc Trans. 39:1565–1570. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Miao HQ, Soker S, Feiner L, et al:
Neuropilin-1 mediates collapsin-1/semaphorin III inhibition of
endothelial cell motility: functional competition of collapsin-1
and vascular endothelial growth factor-165. J Cell Biol.
146:233–242. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Jones DH, Nakashima T, Sanchez OH, et al:
Regulation of cancer cell migration and bone metastasis by RANKL.
Nature. 440:692–696. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Sottnik JL1 and Keller ET: Understanding
and targeting osteoclastic activity in prostate cancer bone
metastases. Curr Mol Med. 13:626–639. 2013. View Article : Google Scholar : PubMed/NCBI
|