Introduction
Hepatocellular carcinoma (HCC) has been identified
as a highly vascularized tumor (1,2).
Therefore, an effective option for the treatment of HCC is to
suppress angiogenesis, as has been widely recognized and utilized
by oncologists worldwide (3,4).
There have been numerous studies on tumor angiogenesis in previous
years. For example, studies have revealed that the major cytokines
responsible for tumor angiogenesis are secreted via the intricate
interaction between tumor cells and stromal cells (5,6).
Among these factors, vascular endothelial growth factor (VEGF) is
pivotal as the primary molecular driver in vasculogenesis (7). Inflammatory and tumor cells secrete
VEGF, which binds and activates the VEGF receptor (VRGFR) on the
surface of endothelial cells (ECs), stimulating various signaling
pathways to accelerate angiogenesis (8–10).
Therefore, the inhibition of tumor angiogenesis via the suppression
of VEGF is favorably recommended by clinicians globally.
Sorafenib, a tyrosine kinase inhibitor, is
recommended and widely used for patients with advanced HCC
(11–13). However, its therapeutic effect in
HCC is limited. Although it has been demonstrated to increase
overall survival rates in patients with advanced stage HCC, the
response rate is low, as resistance to and intolerance of sorafenib
are common. It has been reported that pregnane X and proliferation
and apoptosis adaptor protein 15 mediate sorafenib resistance
(14,15). Therefore, a combination of
sorafenib with other treatment modalities may be advantageous.
Bufalin, the major bioactive component isolated from
toad venom, has been confirmed as a potent antitumor drug through
its effect on tumor cell proliferation, apoptosis and migration
(16–18). In our previous study, it was
demonstrated that the antiangiogenic effect of sorafenib was
markedly improved by combination with bufalin via targeting
AKT/VEGF in human umbilical vein endothelial cells (HUVECs) and HCC
cells (19). Bufalin was also
demonstrated to suppress HCC metastasis via hypoxia inducible
factor/VEGF signaling (20).
Others reported the inhibitory effect of bufalin on HCC cell
proliferation (21,22). However, whether the synergistic
effect of bufalin combined with sorafenib is achieved in HCC cells
by targeting tumor cells and surrounding ECs remains to be
elucidated, and the underlying mechanisms have not been identified.
These issues were considered in the present study.
Materials and methods
Antibodies and reagents
The primary antibodies used in the present study
included: VEGF (Bioworld Technology, Inc., St. Louis Park, MN,
USA), phosphorylated AKT (p-AKT), p-mechanistic target of rapamycin
(p-mTOR), p-phosphatase and tensin homolog (p-PTEN),
p-extracellular signal-regulated kinase (p-ERK1/2), and AKT, mTOR,
PTEN, ERK and GAPDH (Cell Signaling Technology, Inc., Danvers, MA,
USA). The reagents used in the present study included: PI103
(Selleck Chemicals, Houston, TX, USA), the Human Angiogenesis Array
Q1 (RayBiotech, Norcross, GA, USA), sorafenib (Selleck Chemicals)
and bufalin (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany)
Cell lines
The in vitro experiments were performed with
HUVECs from the American Type Culture Collection (Manassas, VA,
USA), and SMMC-7721 and PLC/PRF/5 HCC cells, which were purchased
from the Cell Bank of Type Culture Collection of Chinese Academy of
Sciences (Shanghai, China). The cells were propagated with DMEM
containing 10% fetal bovine serum (FBS; HyClone, GE Healthcare Life
Sciences, Logan, UT, USA) and 1% penicillin/streptomycin at 37°C in
an atmosphere containing 5% CO2.
Animals
Six-week old male Balb/c nude mice were used in the
present study. The study was approved by Fudan University Shanghai
Cancer Center (Shanghai, China). The mice were purchased from Vital
River Laboratory Animal Technology Co., Ltd. (Beijing, China), and
were raised under the following pathogen-free conditions: Room
temperature, 20°C; relative humidity, ~50%. The mice were given
ad libitum access to food and water, and received humane
care according to the principles of animal care of Fudan University
(Shanghai, China). The mice were maintained under a 12-h light/dark
cycle. The principles were as described in the Guide for the Care
and Use of Laboratory Animals issued by the National Institutes of
Health (Bethesda, MD, USA). The mice were randomly divided into
four groups (n=6 mice/group). The mice in the experimental group
were intraperitoneally injected with 1 mg/kg bufalin (5 days/week),
oral uptake of 30 mg/kg/day sorafenib (5 days/week), or combination
treatment with the two drugs. The control mice were treated with
the vehicle only. The treatment lasted for 16 days, following which
the mice were sacrificed, and the tumors and blood were obtained.
All experiments conformed to the ethical principles of animal
experimentation stipulated by Fudan University.
In vivo tumorigenicity assay
A total of 5×106 SMMC-7721 cells in 0.2
ml phosphate-buffered saline (PBS) were injected into the right
flank of each mouse to form subcutaneous tumors. When the volume of
these subcutaneous tumors reached a size of 100–300 mm3,
the mice were treated with intraperitoneal injections of mg/kg
bufalin (5 days/week), 30 mg/kg oral sorafenib (5 days/week), or a
combination of the two drugs. The control mice were treated with
saline. Tumor size was measured every 4 days. The tumor-bearing
mice were sacrificed following 16 days of treatments, and tumors
and blood were extracted. The tumors were weighed and subjected to
immunohistochemistry. All procedures conformed to the ethical
principles of animal experimentation as stipulated by Fudan
University.
Cell viability assay
To evaluate the effects of different treatments on
cell viability, the HCC cells and HUVECs were plated in 96-well
plates at a density of 5,000 cells per well. A total of 20 nM
bufalin, 10 μM sorafenib, or a combination of the two were
added to the wells and incubated at 37°C for 24, 48 or 72 h. The
cells were subjected to a Cell Counting Kit-8 assay (Dojindo
Molecular Technologies, Inc., Kumamoto, Japan), used according to
the manufacturer’s protocol, and the absorbance was measured at a
wavelength of 450 nm with a microplate reader to determine the cell
viability rate.
Cell migration assay
To determine the effects of sorafenib, bufalin, or a
combination of the two on cell migration, an assay was performed
using Transwell inserts (8-μm pore; Corning Incorporated,
Corning, NY, USA). The lower side of the chamber was filled with
condition medium (CM) from the HCC cell lines (SMMC-7721 and
PLC/PRF/5) following different treatments (control, bufalin,
sorafenib, and combination). The HUVEC cells were plated on the
upper side of the chamber. Following a 48-h incubation period, the
migrated cells were fixed with methanol and stained with 0.1%
crystal violet. The numbers of migrated cells were calculated in
three different fields of view, at ×20 magnification using an
Olympus light microscope (Olympus Corporation, Tokyo, Japan).
Collection of CM
The SMMC7721 and PLC/PRF/5 cells were treated with
bufalin, sorafenib, or the two in combination. At 48 h
post-treatment, the cell supernatants were collected as CM. The
collected CM from each group was stored at −80°C if not used
immediately and was thawed prior to use.
Tube formation assay
A 96-well plate was coated with Matrigel and HUVEC
cells were seeded at density of 4×104 cells per well in
100 μl CM, with or without 1 or 6 ng/ml VEGF. The HUVECs
were then incubated at 37°C for 12, 24, 36 or 48 h. The central
region of each well and the tube networks were analyzed using an
Olympus light microscope (Olympus Corporation).
Detection of cytokines associated with
angiogenesis using the human angiogenesis array kit
To determine the cytokines in SMMC-7721 cells
treated with bufalin, sorafenib, or the two in combination, the
Human Angiogenesis Array Q1 kit was used to detect
angiogenesis-related cytokines in the CM extracted from HCC cells
in the different treatment groups, according to the kit protocol.
Following this, the VEGF concentration was determined using an
enzyme-linked immunosorbent assay (ELISA) kit, according to the
manufacturer’s protocol and analyzed using a Labsystems Multiscan
reader (Thermo Fisher Scientific, Inc., Waltham, MA, USA).
Western blot analysis
Western blot analysis was performed as previously
described (23). Briefly, proteins
were extracted from cells following the indicated treatments using
radioimmunoprecipitation assay buffer (R0020; Beijing Solarbio,
Science & Technology Co., Ltd., Beijing, China). Protein
concentrations were assessed using a bicinchoninic acid kit.
Subsequently, protein samples (12 μg) were loaded and
separated by 10% SDS-PAGE for the detection of various proteins.
Proteins were then transferred to polyvinylidene fluoride
membranes, which were blocked in 5% skim milk for 1 h at 37°C. The
expression levels of VEGF, AKT, p-AKT, mTOR, pmTOR, PTEN and p-PTEN
were detected by incubation with their respective primary
antibodies overnight at 4°C, after which the blots were washed
three times with PBS. The primary antibodies used in the present
study included: VEGF (20301555-1, 1:1,000; Bioworld Technology,
Inc.), phosphorylated AKT (p-AKT; 4060, 1:2,000), p-mechanistic
target of rapamycin (p-TOR; 5536, 1:1,000), p-phosphatase and
tensin homolog (p-PTEN; 9551, 1:1,000), p-extracellular
signal-regulated kinase (p-ERK; 4370, 1:2,000), and AKT (4691,
1:1,000), m-TOR (2983, 1:1,000), PTEN (9188, 1:1,000), ERK (9102,
1:1,000) and GAPDH (5174, 1:1,000) (all from Cell Signaling
Technology, Inc.). Finally, the blots were incubated with a
horseradish peroxidase-conjugated immunoglobulin G secondary
antibody (HAF008, 1:5,000; Novus Biologicals, LLC, Littleton, CO,
USA) for 1 h at 37°C, after which proteins were visualized by
enhanced chemiluminescence (WBKLS0500; Merck KGaA).
Semi-quantification of the blots was conducted using ImageJ
software (version no. k 1.45; National Institutes of Health).
ELISA
The mice were treated with the different treatments,
as described above. Blood samples were collected from the eyeballs
of the mice. The expression of VEGF in the blood was measured using
an ELISA kit, according to the manufacturer’s protocol.
Immunohistochemistry
Immunohistochemistry protocols were performed as
described previously (23).
Samples were dehydrated in 70% ethanol, embedded in paraffin and
sectioned (4 μm). Briefly, the tumor sections were stained
with rabbit anti-VEGF (1:1,000; cat. no. AP0742; Bioworld
Technology, Inc.) at 4°C overnight, followed by a goat-anti-rabbit
secondary antibody (1:1,000; cat. no. 7054; Cell Signaling
Technology, Inc.) for 1 h at 37°C. The expression of VEGF was
calculated by multiplying the intensity score by the percentage
score using an Olympus light microscope (Olympus Corporation). The
staining intensity (0, negative; 1, weak; 2, intense) and the ratio
of stained cells (0, ≤10%; 1, 11–25%; 2, 26–50%; 3, 51–75%; 4,
>75%) were evaluated by investigators blinded to the treatment
status.
Statistical analysis
The results were analyzed using SPSS 22.0 software
for Windows (IBM SPSS, Armonk, NY, USA). The comparisons between
two groups were made using Student’s t-test. Multi-group
comparisons of the means were made using one-way analysis of
variance with post hoc contrasts using the Student-Newman-Keuls
test. All experiments were repeated three times. P<0.05 was
considered to indicate a statistically significant difference.
Results
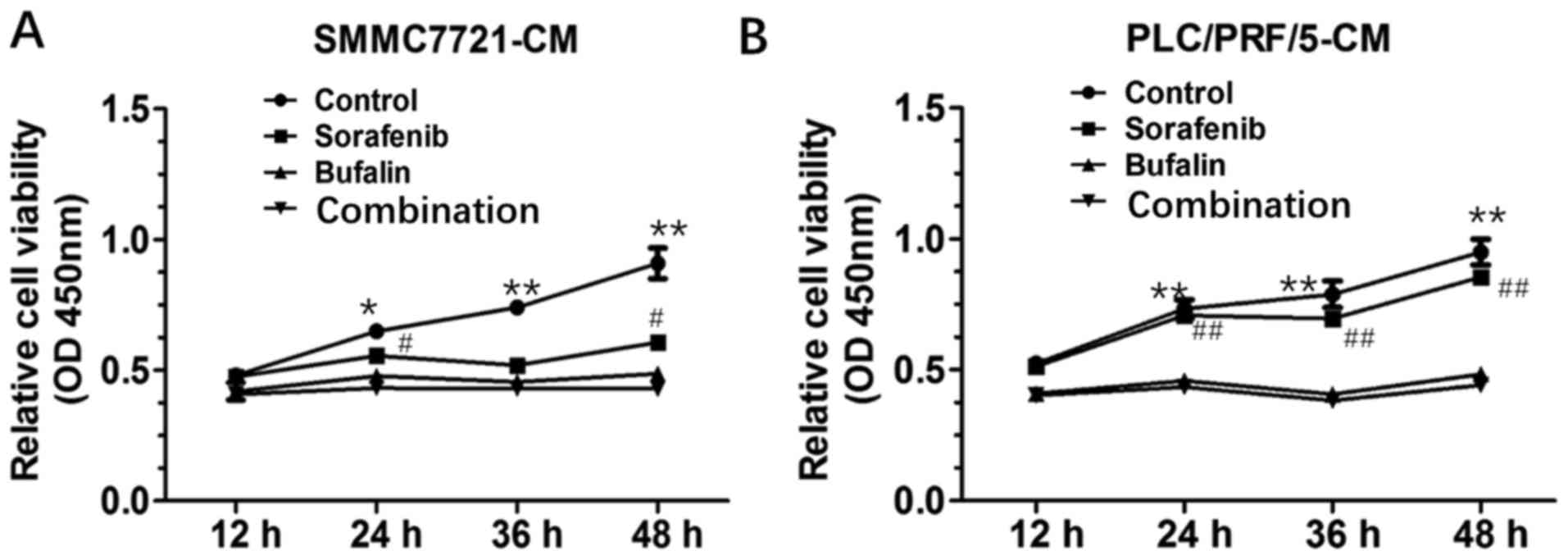
Combination-CM induced the most marked inhibition of
HUVEC proliferation. The SMMC-7721 and PLC/PRF/5 HCC cells were
treated with 10 μM sorafenib, 20 nM bufalin, or a
combination of the two for 12, 24, 36 or 48 h. The CM was collected
48 h following treatment. The HUVECs were subjected to CM from the
HCC cells in the different treatment groups, and the proliferation
of the HUVECs was evaluated. A CCK8 assay was used to determine
whether a synergistic effect was present in the combination-treated
HUVEC. The combination-CM led to a decrease in HUVEC proliferation
from 24 h, compared with that in the control-CM, bufalin-treated CM
and sorafenib-treated CM. The effect of the combination-CM on
HUVECs was most marked at 48 h post-incubation, compared with the
control-CM and sorafenib-treated CM (Fig. 1).
Combination-treated CM inhibits HUVEC
tube formation and migration
The SMMC-7721 and PLC/PRF/5 cells were incubated
with 10 μM sorafenib, 20 nM bufalin, or a combination of the
two. At 12, 24, 36 and 48 h post-treatment, the CM was collected
and incubated with HUVECs. A tube formation assay was performed to
observe the effect of the treatments. It was observed that the
combination-CM inhibited the blood vessel formation of the HUVECs
most markedly, compared with the other groups (Fig. 2A and B). The migration rates of the
HUVECs in the different treatments groups were also detected. The
migration rate of the HUVECs treated with the combination-CM was
reduced compared with that of the HUVECs in the other treatment
groups for the SMMC-7721 and PLC/PRF/5 cells (Fig. 2C and D).
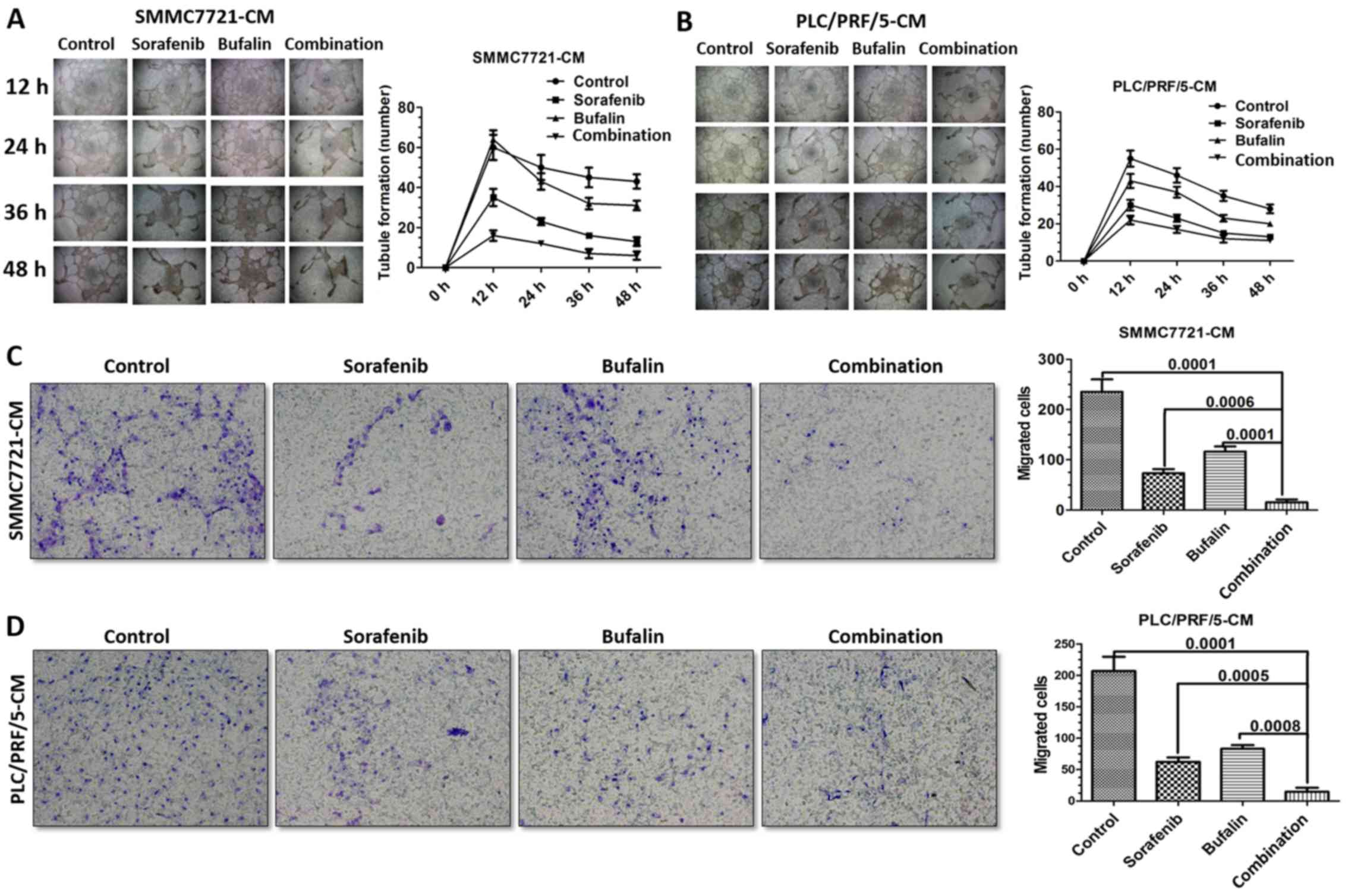 | Figure 2CM from cells receiving combined
treatment results in an enhanced inhibitory effect on HUVEC tube
formation in vitro. (A) Effects of SMMC-7721-CM from
different treatment groups on HUVEC tube formation at 12, 24, 36
and 48 h. (B) Effects of PLC/PRF/5-CM from different treatment
groups on HUVEC tube formation at 12, 24, 36 and 48 h. (C)
Representative images of migrated HUVECs treated with SMMC-7721-CM
from a Transwell migration assay, and statistical analysis. (D)
Representative images of migrated HUVECs treated with PLC/PRF/5-CM
from a Transwell migration assay, and statistical analysis. CM,
conditioned medium; HUVECs, human umbilical vein endothelial cells.
(A and B) Magnification, ×4; (C and D) magnification, ×20. |
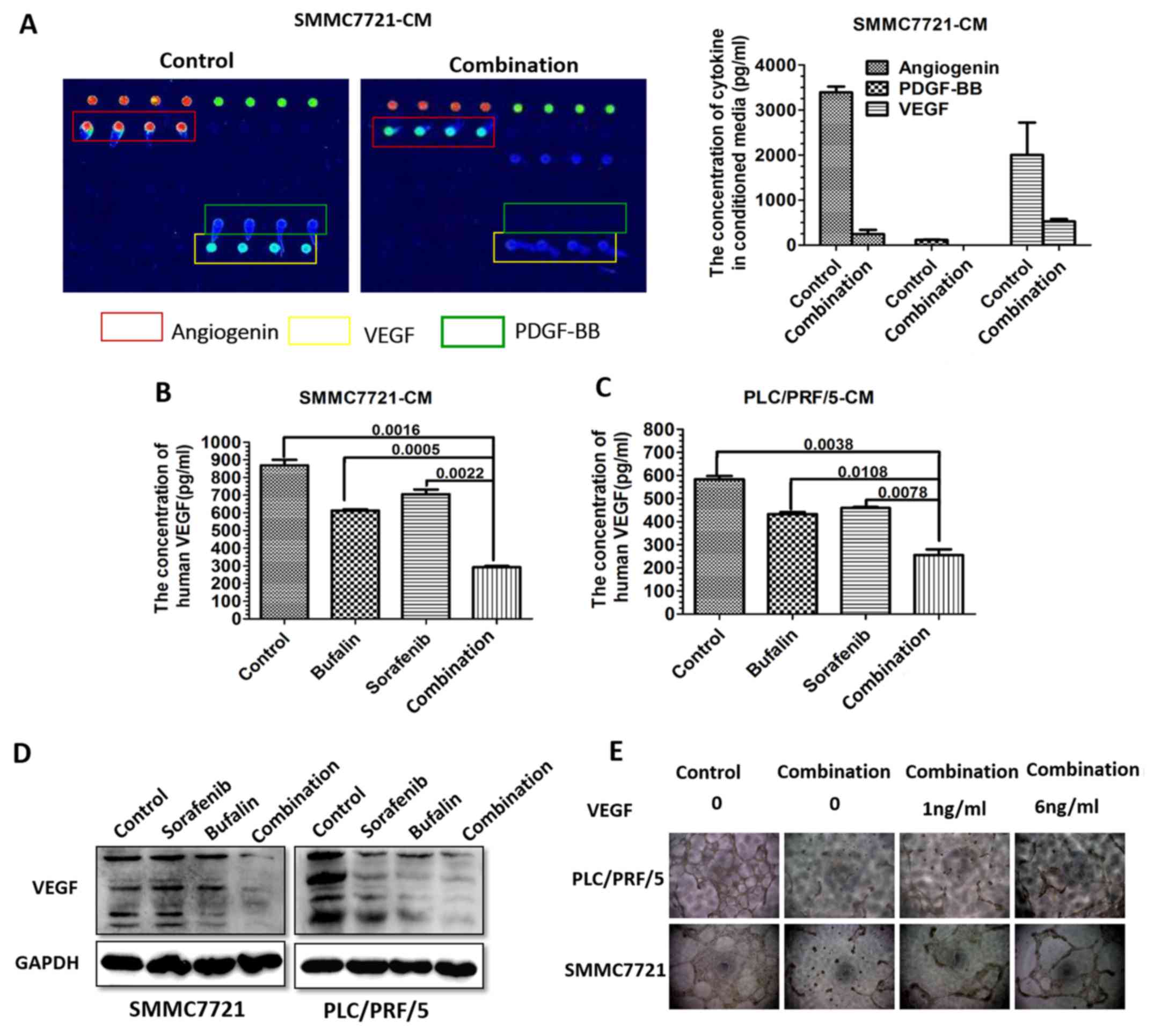
Expression of VEGF is significantly
reduced in combination-CM-treated HCC cells in vitro
As it was previously identified that
combination-CM-treated HUVECs exhibited reduced blood vessel
formation and inhibit migration, it was hypothesized that the
expression of cytokines associated with angiogenesis may be altered
in the CM from the combination-treated SMMC-7721 cells. Therefore,
the CM from the untreated and combination-CM-treated SMMC-7721
cells was analyzed using an angiogenesis-specific array for
cytokine detection. Angiogenin, platelet-derived growth factor
(PDGF-BB) and VEGF were altered most of the cytokines assessed,
indicative of a potential role in facilitating angiogenesis
(Fig. 3A). To further validate the
alterations in cytokine expression detected by the human
angiogenesis array, the concentration of VEGF was measured in the
CM from SMMC-7721 and PLC/PRF/5 cells in the different treatment
groups using ELISA (Fig. 3B and
C). The results of the western blot analysis confirmed the
ELISA results for the SMMC-7721 and PLC/PRF/5 cells (Fig. 3D).
CM from the SMMC7721 and PLC/PRF/5 cells collected
at 48 h was added to HUVECs with or without the addition of 1 or 6
ng/ml VEGF, and tube formation was observed. The combination-CM
suppressed HUVEC vessel formation. The incubation of HUVECs with
VEGF disrupted the inhibitory effect on tubule formation induced by
the addition of combination-CM. The higher concentration of VEGF
resulted in enhanced vessel formation (Fig. 3E).
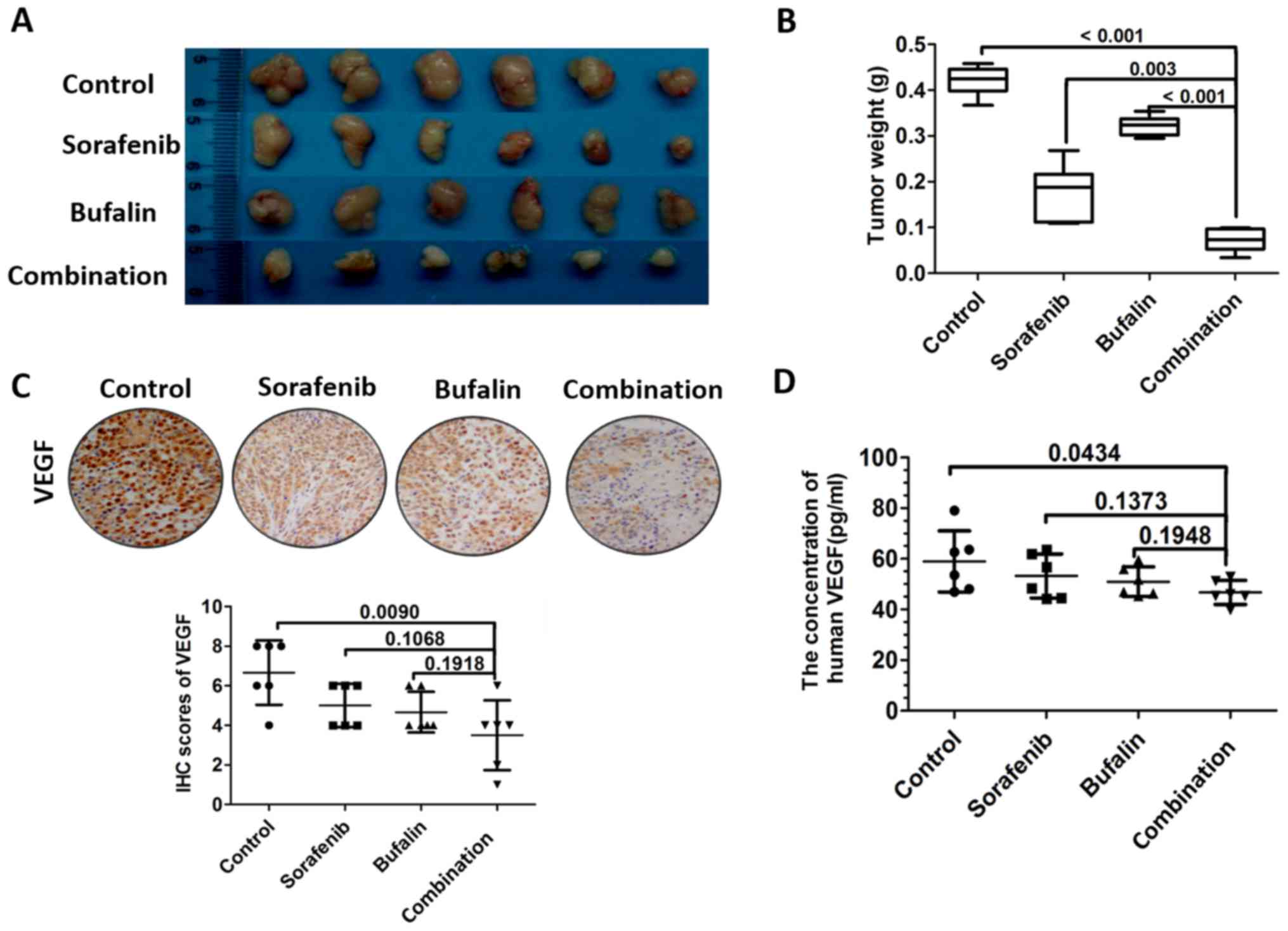
Combination treatment inhibits the
secretion of VEGF in vivo
To measure the effect of the combination treatment
on tumor growth in vivo, a subcutaneous tumor model was
established. Mice received 1 mg/kg/day bufalin, 30 mg/kg/day
sorafenib, or a combination of the two for 16 days. The tumors were
then excised and weighed. The results revealed that the
subcutaneous tumors from mice that received the combination
treatment were smaller, compared with those in the other groups
(Fig. 4A). The tumor weights were
significantly attenuated in mice that received the combination
treatment, compared with those treated with bufalin or sorafenib
alone (Fig. 4B). Further analyses
of the expression of VEGF in tumors revealed that the expression of
VEGF was lowest in the tumors and blood from mice that received the
combination treatment, as determined with immunohistochemistry and
ELISA (Fig. 4C and D).
Combination treatment affects HCC cells
via the mTOR/VEGF pathway
As it was identified that a decrease in the
expression of VEGF occurred due to the combined treatment and that
this may reduce the extent of angiogenesis in HCC, the pathways
associated with VEGF were examined. The proteins upstream of VEGF
were measured in HCC cells in the different treatments groups,
including the phosphorylated and total levels of AKT, mTOR, PTEN
and ERK, with western blot analysis. The HCC cells incubated with
either bufalin or the combination treatment demonstrated a marginal
decrease in the expression of p-AKT. The expression of p-mTOR was
also reduced in the HCC cells incubated with bufalin or the
combination treatment. However, no significant alterations were
identified in the phosphorylation of PTEN or ERK (Fig. 5A).
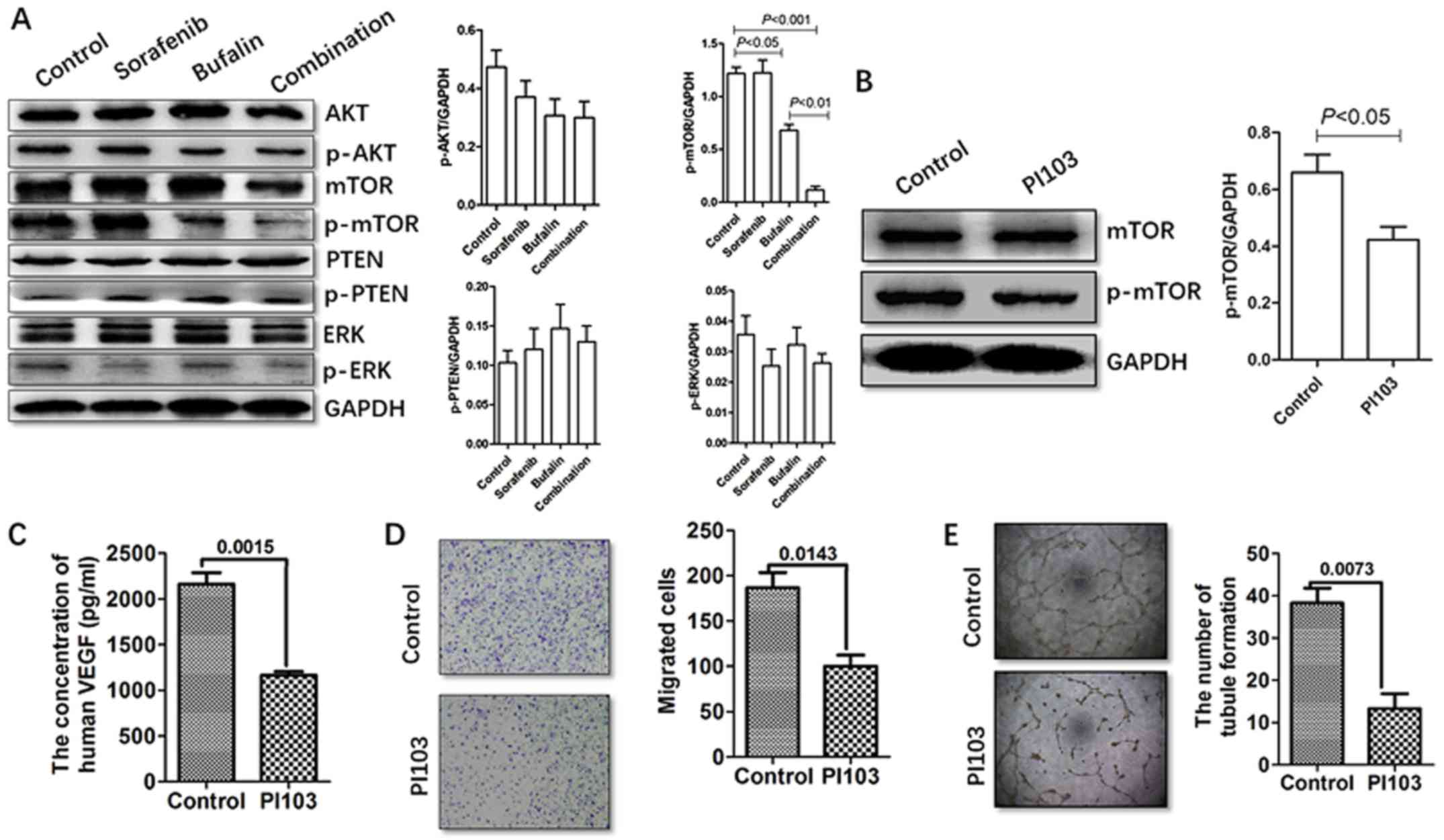 | Figure 5Combination CM regulates the
secretion of VEGF via the mTOR pathway. (A) Expression levels of
p-AKT, p-mTOR, p-PTEN and p-ERK were detected in HCC cells in
different treatment groups by western blot analysis. (B) HCC cells
were treated with the PI3K/AKT inhibitor PI103, followed by bufalin
for 48 h. The HCC cells were harvested, and p-mTOR and mTOR were
detected by western blot analysis. (C) HCC cells were treated with
the PI3K/AKT inhibitor PI103 (2 μM). The secretion of VEGF
was determined using an enzyme-linked immunosorbent assay. (D) HCC
cells were treated with the PI3K/AKT inhibitor PI103.
Representative images of the migrated HCC cells following
incubation with CM derived from cells in the different treatment
groups in the Transwell migration assay are presented and numbers
shown. Magnification, ×20. (E) Human umbilical vein endothelial
cells were treated with a PI3K/AKT inhibitor PI103. Magnification,
×4. Representative images of tubule formation following incubation
in CM derived from different treatment groups, and analysis of the
number of tubes formed is presented. CM, conditioned medium; VEGF,
vascular endothelial growth factor; mTOR, mechanistic target of
rapamycin; PTEN, phosphatase and tensin homolog; ERK, extracellular
signal-regulated kinase; HCC, hepatocellular carcinoma; PI3K,
phosphoinositide 3-kinase; p-, phosphorylated. |
As a decrease was observed in the expression of
p-AKT and p-mTOR in the combination-treated SMMC-7721 cells, their
involvement in the combination-induced reduction of VEGF was
examined. PI103 (2 μM) was selected as a specific
pharmacological inhibitor of the phosphoinositide 3-kinase
(PI3K)/AKT/mTOR pathway. The results demonstrated that pretreatment
with PI103 partially reversed the phosphorylation of mTOR (Fig. 5B). It was also observed that the
expression of VEGF was significantly downregulated in the HCC cells
pretreated with PI103, indicating the potential role of the mTOR
pathway in modulating VEGF (Fig.
5C). The present study also evaluated the effect of the CM from
cells pretreated with PI103 on HUVEC migration and tube formation.
Compared with the CM from the non-pretreated cells, HUVEC migration
and tube formation were suppressed by pretreatment with PI103
(Fig. 5D and E). Collectively,
these results demonstrated that the mTOR pathway was involved in
the reduced expression of VEGF induced by combination
treatment.
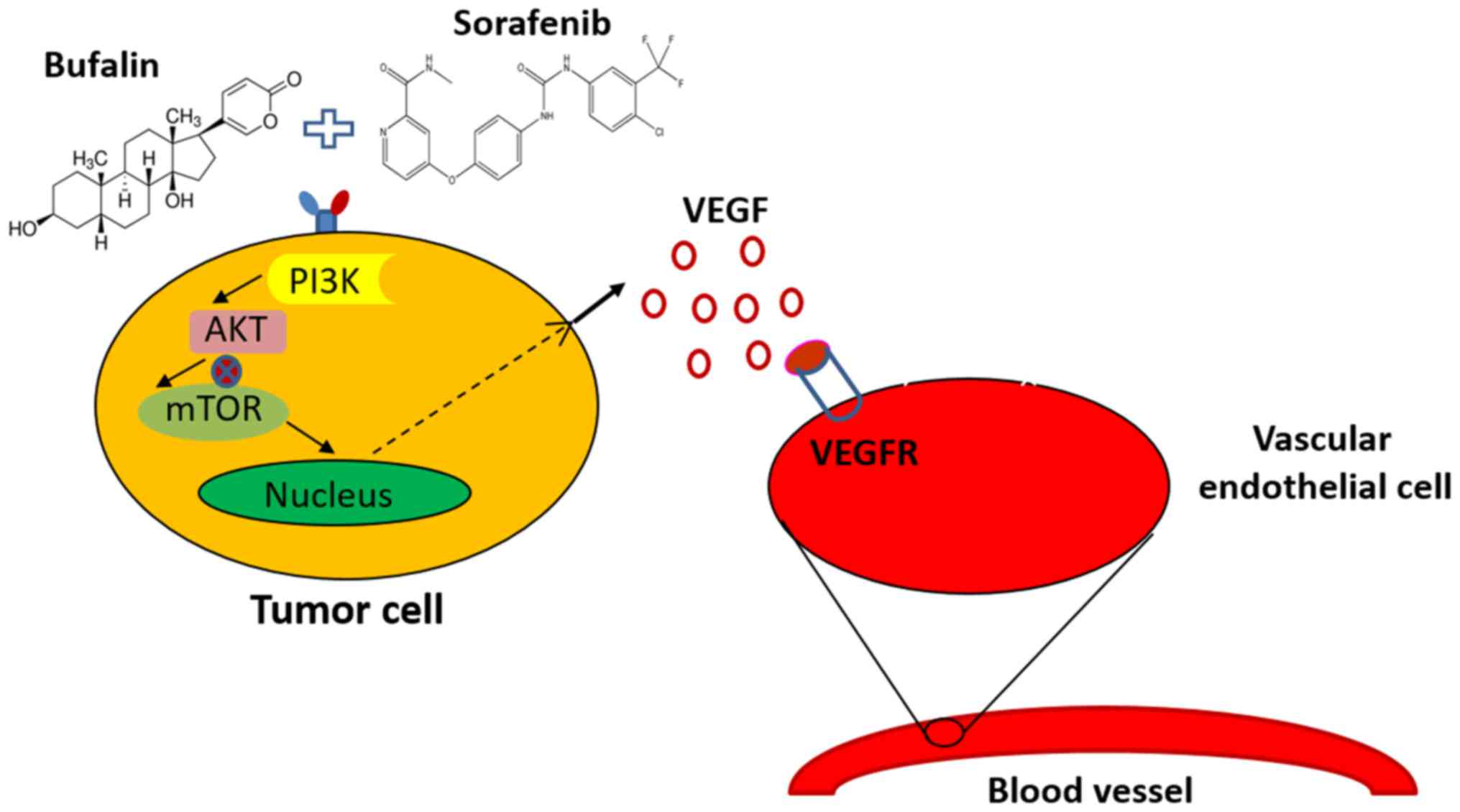
Proposed mechanism by which bufalin
augments the sorafenib-induced inhibition of angiogenesis through
the mTOR/VEGF signaling pathway in HCC
The present study identified bufalin as asynergistic
agent for HCC cells treated with sorafenib by regulating the
mTOR/VEGF pathway, leading to the reduced secretion of VEGF. The
reduced binding of VEGF to VEGFR on the HUVECs led to reduced tumor
angiogenesis. To the best of our knowledge, this is the first
evidence of the synergistic anti-hepatoma effect of bufalin
combined with sorafenib via the tumor vascular microenvironment by
targeting mTOR/VEGF signaling (Fig.
6).
Discussion
Angiogenesis has been reported to be intrinsically
linked with tumor growth, metastasis and development. Therefore,
anti-angiogenic cancer therapies have received much attention and
have progressed rapidly (2,24–27).
As HCC is a highly vascularized tumor, inhibition of its
angiogenesis by multiple drugs is considered a promising approach
with therapeutic value.
Increasing evidence is available regarding the
antitumor effect of bufalin on tumor cells. It has been reported to
suppress proliferation and migration, and induce apoptosis in tumor
cells (19–21). However, there are relatively few
reports regarding the effect of bufalin on ECs. In our previous
study, it was demonstrated that the synergistic effect of bufalin
with sorafenib may be due to inhibition of the AKT/VEGF signaling
pathway in HUVECs (22). Given the
inhibitory effect of bufalin treatment on tumor cells and the
established interactions between tumor cells and ECs, it was
hypothesized that the synergistic role of bufalin with sorafenib
may be partly explained by alterations to the cytokines in the
tumor microenvironment, which affect the angiogenesis and migration
of ECs.
To determine the effect of the combined treatment on
HCC proliferation, two HCC cell lines (SMMC-7721 and PLC/PRF/5)
were used. The proliferation assay demonstrated that HCC cell
viability was significantly reduced in HCC cells exposed to the
combined treatment, compared with those treated with sorafenib
alone. To test the hypothesis described above, CM from HCC cells in
each of the different treatment groups was collected at 24 h
post-incubation, and cultured with HUVECs for 12, 24, 36 or 48 h.
As described in the results, the HUVECs incubated with the
combination-CM underwent the least tube formation, suggesting there
may be either less pro-angiogenic or more antiangiogenic cytokines
produced by the HCC cells exposed to combination treatment.
Furthermore, the number of migrated HUVECs was lowest following
treatment with the combination-CM, as determined by a Transwell
migration assay, which may be reflective of a
metastasis-suppressive effect of the combined treatment. These
results were in accord, highlighting the dual properties of
combined treatment in the inhibition of endothelial angiogenesis
and metastasis.
Subsequently, the present study aimed to ascertain
the mechanisms that led to the attenuation of angiogenesis and
metastasis in the HUVECs. Therefore, a human angiogenesis array was
adopted. The results demonstrated that VEGF, PDGF-BB and angiogenin
were the cytokines with the most marked significant alteration
between the control cells and the cells exposed to the combined
treatment. Of the three most altered cytokines, VEGF has been
investigated most for its potent pro-angiogenic properties. VEGF
initiates signaling pathways in ECs to promote proliferation,
migration, survival and vascular permeability. VEGF inhibitors have
been developed, as the overexpression of VEGF is frequently
associated with a poorer prognosis in various types of cancer
(28,29). Subsequent ELISA and western blot
analyses also confirmed the cytokine changes reported in the human
angiogenesis array. To further corroborate that the inhibitory
effect of combined treatment on the secretion of VEGF mediated the
effect on angiogenesis, the HUVECs in the different treatment
groups were treated with VEGF. The extent of vessel sprouting in
the combination treatment HUVEC group was significantly reduced,
which was reversed by incubation with VEGF. Therefore, the
hypothesis was confirmed and validated.
The present study is notable as it focused not only
on tumor cells, but also on ECs. The tumor microenvironment is a
popular focus of investigations; the crosstalk between ECs and HCC
cells can affect angiogenesis and metastasis via the regulation of
inflammatory cytokines (30–32).
The present study observed that the combination-CM derived from HCC
cells attenuated angiogenesis and metastasis, indicating the
potential of the drug combination in reducing the secretion of
angiogenic cytokines.
In addition, in vivo experiments were
performed to analyze the combined effect of bufalin and sorafenib.
The expression of VEGF was reduced in the subcutaneous tumors and
serum of mice receiving the combined treatment, compared with that
in the mice receiving the drugs individually, as demonstrated by
immunohistochemistry and ELISA. These results further demonstrated
the synergistic role of bufalin and sorafenib in tumor inhibition
via the suppression of VEGF.
There are numerous pathways involved in
angiogenesis. For example, PI3K/AKT, PTEN/AKT, AKT/mTOR and
mitogen-activated protein kinase kinase/ERK have all been
associated with tumor angiogenesis via the upregulation of VEGF
(33–36). Previous studies have demonstrated
that the VEGF/PI3K/AKT pathway is associated with Nogo-B-induced
angiogenesis in primary human retinal ECs (37). VEGF activates PI3K, which triggers
the phosphorylation of AKT, leading to increased endothelial nitric
oxide synthase activity. PI3K/AKT/mTOR is a pivotal signaling
pathway responsible for cell proliferation, metabolism and motility
(38,39). Previous studies have also confirmed
its involvement in angiogenesis (40,41).
The present study determined the relative expression
of the main proteins involved in the PI3K/AKT, PTEN/AKT, AKT/mTOR
and MEK/ERK pathways. The results demonstrated that the level of
p-AKT was significantly altered in the HCC cells that received the
combined treatment, compared with that in the HCC cells treated
with sorafenib alone. The level of p-m-TOR was decreased in the
combination treatment group, compared with that in the SMMC-7721
cells treated with sorafenib. To further validate the role of the
PI3K/AKT pathway in the altered expression of mTOR, the PI3K
inhibitor PI103 was used. Incubation of the HCC cells with P1103
led to a marginal reduction of p-mTOR, whereas the application of
PI103 led to a significant reduction in the expression of VEGF,
reflecting the role of PI3K/AKT in the secretion of VEGF. This was
also demonstrated in the migration and tube formation assays.
However, there were a number of limitations in the
present study. First, the antiangiogenic effect of the combined
drug was a comprehensive result of the balance between the
production of fewer proangiogenic factors or increased secretion of
antiangiogenic factors. There may be several other cytokine changes
in the combination-treated CM, compared with the mono-drug-treated
CM in addition to VEGF, however, the present study focused on VEGF
only. Investigations are required to ascertain other angiogenic
factor changes induced by the combination treatment in the future.
Second, additional in vivo experiments are required to
confirm the synergistic anti-hepatoma effect of the combined
treatment of sorafenib and bufalin via mediating the tumor vascular
microenvironment by targeting mTOR/VEGF signaling.
In conclusion, the present study identified bufalin
as a synergistic agent for HCC treated with sorafenib by regulating
tumor cells and ECs. The combined treatment affected HCC cells via
the mTOR/VEGF pathway, leading to the reduced secretion of VEGF.
The decreased secretion of VEGF produced by SMMC-7721 cells
resulted in reduced migration and tube formation in HUVECs.
Therefore, the mTOR/VEGF pathway may be one of the targets
accounting for the enhanced effect of sorafenib treatment with
bufalin.
Acknowledgments
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant nos. 81573753 and
81603348), the China Postdoctoral Fund (grant no. 21300075311104)
and the Shandong Postdoctoral Innovation Special Fund (grant no.
201602012).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors’ contributions
HW performed the in vivo studies and designed
the study. CZ performed the in vitro studies and wrote the
original manuscript. HC conducted the western blot experiments and
analysed the data. ZM designed the study and reviewed the
article.
Ethics approval and consent to
participate
The study was approved by Fudan University Shanghai
Cancer Center and all experiments conformed to the ethical
principles of animal experimentation stipulated by Fudan
University. The principles were as described in the Guide for the
Care and Use of Laboratory Animals issued by the National
Institutes of Health.
Consent for publication
Not applicable.
Competing interests
The authors declare that there are no competing
interests.
References
|
1
|
Borzio M, Dionigi E, Parisi G, Raguzzi I
and Sacco R: Management of hepatocellular carcinoma in the elderly.
World J Hepatol. 7:1521–1529. 2015. View Article : Google Scholar
|
|
2
|
Folkman J: Tumor angiogenesis: Therapeutic
implications. N Engl J Med. 285:1182–1186. 1971. View Article : Google Scholar
|
|
3
|
Folkman J: Anti-angiogenesis: New concept
for therapy of solid tumors. Ann Surg. 175:409–416. 1972.
View Article : Google Scholar
|
|
4
|
Folkman J: Role of angiogenesis in tumor
growth and metastasis. Semin Oncol. 29(Suppl 16): 15–18. 2002.
View Article : Google Scholar
|
|
5
|
Portillo-Lara R and Annabi N:
Microengineered cancer-on-a-chip platforms to study the metastatic
microenvironment. Lab Chip. 16:4063–4081. 2016. View Article : Google Scholar
|
|
6
|
Maj E, Papiernik D and Wietrzyk J:
Antiangiogenic cancer treatment: The great discovery and greater
complexity (Review). Int J Oncol. 49:1773–1784. 2016. View Article : Google Scholar
|
|
7
|
Yang Y, Zhang Y, Iwamoto H, Hosaka K, Seki
T, Andersson P, Lim S, Fischer C, Nakamura M, Abe M, et al:
Discontinuation of anti-VEGF cancer therapy promotes metastasis
through a liver revascularization mechanism. Nat Commun.
7:126802016. View Article : Google Scholar
|
|
8
|
Falchook GS, Moulder S, Naing A, Wheler
JJ, Hong DS, Piha-Paul SA, Tsimberidou AM, Fu S, Zinner R, Janku F,
et al: A phase I trial of combination trastuzumab, lapatinib, and
bevacizumab in patients with advanced cancer. Invest New Drugs.
33:177–186. 2015. View Article : Google Scholar
|
|
9
|
Fan X, Krieg S, Kuo CJ, Wiegand SJ,
Rabinovitch M, Druzin ML, Brenner RM, Giudice LC and Nayak NR: VEGF
blockade inhibits angiogenesis and reepithelialization of
endometrium. FASEB J. 22:3571–3580. 2008. View Article : Google Scholar
|
|
10
|
Hong DS, Garrido-Laguna I, Ekmekcioglu S,
Falchook GS, Naing A, Wheler JJ, Fu S, Moulder SL, Piha-Paul S,
Tsimberidou AM, et al: Dual inhibition of the vascular endothelial
growth factor pathway: A phase 1 trial evaluating bevacizumab and
AZD2171 (cediranib) in patients with advanced solid tumors. Cancer.
120:2164–2173. 2014. View Article : Google Scholar
|
|
11
|
Liu L, Cao Y, Chen C, Zhang X, McNabola A,
Wilkie D, Wilhelm S, Lynch M and Carter C: Sorafenib blocks the
RAF/MEK/ERK pathway, inhibits tumor angiogenesis, and induces tumor
cell apoptosis in hepatocellular carcinoma model PLC/PRF/5. Cancer
Res. 66:11851–11858. 2006. View Article : Google Scholar
|
|
12
|
Pressiani T, Boni C, Rimassa L, Labianca
R, Fagiuoli S, Salvagni S, Ferrari D, Cortesi E, Porta C,
Mucciarini C, et al: Sorafenib in patients with Child-Pugh class A
and B advanced hepatocellular carcinoma: A prospective feasibility
analysis. Ann Oncol. 24:406–411. 2013. View Article : Google Scholar
|
|
13
|
Sposito C, Mariani L, Germini A, Flores
Reyes M, Bongini M, Grossi G, Bhoori S and Mazzaferro V:
Comparative efficacy of sorafenib versus best supportive care in
recurrent hepatocellular carcinoma after liver transplantation: A
case-control study. J Hepatol. 59:59–66. 2013. View Article : Google Scholar
|
|
14
|
Feng F, Jiang Q, Cao S, Cao Y, Li R, Shen
L, Zhu H, Wang T, Sun L, Liang E, et al: Pregnane X receptor
mediates sorafenib resistance in advanced hepatocellular carcinoma.
Biochim Biophys Acta. 1862:1017–1030. 2018. View Article : Google Scholar
|
|
15
|
Quintavalle C, Hindupur SK, Quagliata L,
Pallante P, Nigro C, Condorelli G, Andersen JB, Tagscherer KE, Roth
W, Beguinot F, et al: Phosphoprotein enriched in diabetes
(PED/PEA15) promotes migration in hepatocellular carcinoma and
confers resistance to sorafenib. Cell Death Dis. 8:e31382017.
View Article : Google Scholar
|
|
16
|
Takai N, Ueda T, Nishida M, Nasu K and
Narahara H: Bufalin induces growth inhibition, cell cycle arrest
and apoptosis in human endometrial and ovarian cancer cells. Int J
Mol Med. 21:637–643. 2008.
|
|
17
|
Huang WW, Yang JS, Pai SJ, Wu PP, Chang
SJ, Chueh FS, Fan MJ, Chiou SM, Kuo HM, Yeh CC, et al: Bufalin
induces G0/G1 phase arrest through inhibiting the levels of cyclin
D, cyclin E, CDK2 and CDK4, and triggers apoptosis via
mitochondrial signaling pathway in T24 human bladder cancer cells.
Mutat Res. 732:26–33. 2012. View Article : Google Scholar
|
|
18
|
Yin JQ, Shen JN, Su WW, Wang J, Huang G,
Jin S, Guo QC, Zou CY, Li HM and Li FB: Bufalin induces apoptosis
in human osteosarcoma U-2OS and U-2OS methotrexate300-resistant
cell lines. Acta Pharmacol Sin. 28:712–720. 2007. View Article : Google Scholar
|
|
19
|
Wang H, Zhang C, Ning Z, Xu L, Zhu X and
Meng Z: Bufalin enhances anti-angiogenic effect of sorafenib via
AKT/VEGF signaling. Int J Oncol. 48:1229–1241. 2016. View Article : Google Scholar
|
|
20
|
Wang H, Zhang C, Xu L, Zang K, Ning Z,
Jiang F, Chi H, Zhu X and Meng Z: Bufalin suppresses hepatocellular
carcinoma invasion and metastasis by targeting HIF-1α via the
PI3K/AKT/mTOR pathway. Oncotarget. 7:20193–20208. 2016.
|
|
21
|
Qiu DZ, Zhang ZJ, Wu WZ and Yang YK:
Bufalin, a component in Chansu, inhibits proliferation and invasion
of hepatocellular carcinoma cells. BMC Complement Altern Med.
13:1852013. View Article : Google Scholar
|
|
22
|
Gao Y, Li HX, Xu LT, Wang P, Xu LY, Cohen
L, Yang PY, Gu K and Meng ZQ: Bufalin enhances the
anti-proliferative effect of sorafenib on human hepatocellular
carcinoma cells through downregulation of ERK. Mol Biol Rep.
39:1683–1689. 2012. View Article : Google Scholar
|
|
23
|
Zhuang L, Xu L, Wang P, Jiang Y, Yong P,
Zhang C, Zhang H, Meng Z and Yang P:
Na+/K+-ATPase α1 subunit, a novel therapeutic
target for hepatocellular carcinoma. Oncotarget. 6:28183–28193.
2015.
|
|
24
|
Butler JM, Kobayashi H and Rafii S:
Instructive role of the vascular niche in promoting tumour growth
and tissue repair by angiocrine factors. Nat Rev Cancer.
10:138–146. 2010. View
Article : Google Scholar
|
|
25
|
Kerbel RS: Tumor angiogenesis. N Engl J
Med. 358:2039–2049. 2008. View Article : Google Scholar
|
|
26
|
Ferrara N and Kerbel RS: Angiogenesis as a
therapeutic target. Nature. 438:967–974. 2005. View Article : Google Scholar
|
|
27
|
Bridges EM and Harris AL: The angiogenic
process as a therapeutic target in cancer. Biochem Pharmacol.
81:1183–1191. 2011. View Article : Google Scholar
|
|
28
|
Fu L, Shi K, Wang J, Chen W, Shi D, Tian
Y, Guo W, Yu W, Xiao X, Kang T, et al: TFAP2B overexpression
contributes to tumor growth and a poor prognosis of human lung
adenocarcinoma through modulation of ERK and VEGF/PEDF signaling.
Mol Cancer. 13:892014. View Article : Google Scholar
|
|
29
|
Chen C, Chi H, Min L and Junhua Z:
Downregulation of guanine nucleotide-binding protein beta 1 (GNB1)
is associated with worsened prognosis of clearcell renal cell
carcinoma and is related to VEGF signaling pathway. J BUON.
22:1441–1446. 2017.
|
|
30
|
Neiva KG, Zhang Z, Miyazawa M, Warner KA,
Karl E and Nör JE: Cross talk initiated by endothelial cells
enhances migration and inhibits anoikis of squamous cell carcinoma
cells through STAT3/Akt/ERK signaling. Neoplasia. 11:583–593. 2009.
View Article : Google Scholar
|
|
31
|
Warner KA, Miyazawa M, Cordeiro MM, Love
WJ, Pinsky MS, Neiva KG, Spalding AC and Nör JE: Endothelial cells
enhance tumor cell invasion through a crosstalk mediated by CXC
chemokine signaling. Neoplasia. 10:131–139. 2008. View Article : Google Scholar
|
|
32
|
Manzi M, Bacigalupo ML, Carabias P, Elola
MT, Wolfenstein-Todel C, Rabinovich GA, Espelt MV and Troncoso MF:
Galectin-1 controls the proliferation and migration of liver
sinusoidal endothelial cells and their interaction with
hepatocarcinoma cells. J Cell Physiol. 231:1522–1533. 2016.
View Article : Google Scholar
|
|
33
|
Liu WL, Gao M, Tzen KY, Tsai CL, Hsu FM,
Cheng AL and Cheng JC: Targeting phosphatidylinositide 3-kinase/Akt
pathway by BKM120 for radiosensitization in hepatocellular
carcinoma. Oncotarget. 5:3662–3672. 2014. View Article : Google Scholar
|
|
34
|
Paplomata E and O’Regan R: The
PI3K/AKT/mTOR pathway in breast cancer: Targets, trials and
biomarkers. Ther Adv Med Oncol. 6:154–166. 2014. View Article : Google Scholar
|
|
35
|
Lee MS, Jeong MH, Lee HW, Han HJ, Ko A,
Hewitt SM, Kim JH, Chun KH, Chung JY, Lee C, et al: PI3K/AKT
activation induces PTEN ubiquitination and destabilization
accelerating tumourigenesis. Nat Commun. 6:77692015. View Article : Google Scholar
|
|
36
|
Zhang D, Li X, Yao Z, Wei C, Ning N and Li
J: GABAergic signaling facilitates breast cancer metastasis by
promoting ERK1/2-dependent phosphorylation. Cancer Lett.
348:100–108. 2014. View Article : Google Scholar
|
|
37
|
Zhang Y, Wang L, Zhang Y, Wang M, Sun Q,
Xia F, Wang R and Liu L: Nogo-B promotes angiogenesis in
proliferative diabetic retinopathy via VEGF/PI3K/Akt pathway in an
autocrine manner. Cell Physiol Biochem. 43:1742–1754. 2017.
View Article : Google Scholar
|
|
38
|
Philippova M, Joshi MB, Pfaff D,
Kyriakakis E, Maslova K, Erne P and Resink TJ: T-cadherin
attenuates insulin-dependent signalling, eNOS activation, and
angiogenesis in vascular endothelial cells. Cardiovasc Res.
93:498–507. 2012. View Article : Google Scholar
|
|
39
|
Sharma S, Guru SK, Manda S, Kumar A,
Mintoo MJ, Prasad VD, Sharma PR, Mondhe DM, Bharate SB and Bhushan
S: A marine sponge alkaloid derivative 4-chloro fascaplysin
inhibits tumor growth and VEGF mediated angiogenesis by disrupting
PI3K/Akt/mTOR signaling cascade. Chem Biol Interact. 275:47–60.
2017. View Article : Google Scholar
|
|
40
|
Zhang W, Xiong Z, Wei T, Li Q, Tan Y, Ling
L and Feng X: Nuclear factor 90 promotes angiogenesis by regulating
HIF-1α/VEGF-A expression through the PI3K/Akt signaling pathway in
human cervical cancer. Cell Death Dis. 9:2762018. View Article : Google Scholar
|
|
41
|
Song ZY, Wang F, Cui SX and Qu XJ:
Knockdown of CXCR4 inhibits CXCL12-induced angiogenesis in HUVECs
through downregulation of the MAPK/ERK and PI3K/AKT and the
Wnt/β-catenin pathways. Cancer Invest. 36:10–18. 2018. View Article : Google Scholar
|