Introduction
Despite multiple clinical trials aimed at improving
patient survival rates, lung cancer is the second leading cause of
cancer-related mortality worldwide following breast cancer. Among
all types of pulmonary cancer, ~85% of cases are diagnosed,
commonly at advanced stages, as non-small cell lung cancer (NSCLC)
(1). Lung adenocarcinoma is the
predominant NSCLC histological subtype, which accounts for 20–30%
of primary lung cancer cases among patients <45 years of age,
regardless of smoking history (2).
The majority of patients with NSCLC exhibit advanced disease, thus
are unsuitable for surgery; therefore, chemotherapy remains the
cornerstone of treatment.
Histone deacetylases (HDACs) are enzymes that remove
histone acetylation markers, resulting in compaction of the
chromatin structure and transcriptional repression (3). In addition to histones, HDACs have
various nonhistone protein substrates involved in the regulation of
gene expression, cell proliferation, cell migration, apoptosis and
angiogenesis (4). A number of
naturally occurring and synthetic HDAC inhibitors have been shown
in preclinical studies to exert potent anticancer activity
(5).
HDAC1 and 2 are similar enzymes that belong to the
class I HDAC family of molecules. In vivo, HDAC1 and 2
cooperate within a complex of proteins including Sin3, nucleosome
remodeling and deacetylating, and repressor element-1 silencing
transcription factor corepressor 1 (6). In addition to functions exerted
through these complexes, HDAC1 and 2 also bind directly to
DNA-binding proteins, including Yin and Yang 1, retinoblastoma
binding protein-1 and specificity protein 1 (6).
c-Myc, as a transcription factor, is responsible for
the regulation of multiple genes associated with cellular
activities including cellular proliferation, growth, apoptosis and
differentiation (7). Due to the
extensive functions of c-Myc, it is considered to be a potent
oncogene. In addition, downregulated c-Myc expression has been
observed in ~70% of all types of human tumor (8). The expression of c-Myc is controlled
at a number of different levels, including gene transcription, mRNA
stability and through the post-translational control of protein
stability (9–11). Post-translational regulation of
c-Myc is mediated by S-phase kinase-associated protein 2 (Skp2) and
F-box and WD repeat-containing protein 7 (Fbw7) (12), two recognition subunits of the
Skp1/Cullin/F-box protein complex-type E3 ligase that have the
ability to recognize the substrates responsible for proteasomal
degradation.
Skp2 overexpression is frequently observed in human
cancer specimens and has been suggested to be an oncogene. With the
exception of c-Myc, Skp2 is reported to recognize cyclin-dependent
kinase inhibitors and tumor suppressor proteins such as p27 Kip1,
p57 Kip2, p130 and Tob1 (13–16).
Fbw7 has frequently been found to be inactivated by mutation,
deletion or promoter hypermethylation in multiple types of
neoplasm, including breast cancer (17,18),
colon cancer (19,20) and leukemia (21).
The involvement of Fbw7 deficiency in human cancer
drug resistance has been recently detected (22,23).
In the regulation of c-Myc, c-Myc phosphorylation at Thr58 is
required for Fbw7-mediated proteasomal degradation. Glycogen
synthase kinase 3β (GSK3β) is the only known kinase that
phosphorylates c-Myc at Thr58 (24). In contrast to Fbw7, Skp2-mediated
ubiquitylation of c-Myc does not require its phosphorylation
(25). Fbw7 is expressed as three
isoforms in humans, designated α, β and γ, located in the nucleus,
cytoplasm and nucleolus, respectively (26,27).
Ampelopsin [AMP;
(2R,3R)-3,5,7-trihydroxy-2-(3,4,5-trihydroxyphenyl)-2,3-dihydrochromen-4-one;
Fig. 1A], is a naturally occurring
flavonoid isolated from the plant species Ampelopsis
grossedentata (Hand.-Mazz) W.T. Wang. The anticancer activity
of AMP has been reported in various human cell lines, including
bladder carcinoma (28), melanoma
(29), GLC-82 lung cancer
(30), hepatocellular carcinoma
(31), K562/ADR leukemia (32) and prostate cancer cells (33). AMP activity is mediated by the
induction of apoptosis and cell differentiation, which is regulated
by various genes and proteins (34). In the present study, the effect of
AMP upon cell proliferation and apoptosis in the A549 human lung
adenocarcinoma epithelial cell line was assessed. In addition, the
regulatory effects and underlying functions of the c-Myc/Skp2/Fbw7
and HDAC1/2 pathways involved in the apoptotic effect were
investigated.
Materials and methods
Reagents and antibodies
AMP (purity >98%) was supplied by the Institute
of Biology at Guizhou Academy of Sciences (Guiyang, China). The
following antibodies were used: Fbw7 (cell division control protein
4, H-300) and phosphorylated-(p−)c-Myc (Thr 58) were provided by
Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA); c-Myc and
GSK3β were bought from Proteintech Group, Inc. (Chicago, IL, USA);
HDAC1 and 2, Skp2, survivin, B-cell lymphoma 2 (Bcl-2) and β-actin
were obtained from Wuhan Boster Biological Technology, Ltd. (Wuhan,
China); myeloid cell leukemia 1 (Mcl-1) and X-linked inhibitor of
apoptosis protein (XIAP) were provided by Bioss (Beijing, China);
and poly ADP ribose polymerase (PARP) was purchased from Sino
Biological Inc. (Beijing, China).
Cell culture
A549 human pulmonary adenocarcinoma cells were
provided by the Cell Bank of the Animal Experiment Center, North
School Region, Sun Yat-Sen University (Guangzhou, China). The A549
cells were cultured in RPMI-1640 medium containing 10% fetal bovine
serum (Hangzhou Sijiqing Biological Engineering Materials Co.,
Ltd., Hangzhou, China) at 37°C and 5% CO2.
MTT assay
To measure cell viability, A549 cells harvested with
trypsin were seeded onto 96-well plates at a density of
1×104 cells per well. Following overnight incubation,
the culture medium was removed and the cells were incubated with
different concentrations of AMP in culture medium. After 48 h, MTT
was added to each well and incubated at 37°C for an additional 4 h
to allow mitochondrial dehydrogenase to convert the MTT into
insoluble formazan crystals. The medium was then discarded and 100
μl dimethylsulfoxide (Sigma, St. Louis, MO, USA) was added to each
well to dissolve the formazan crystals. The absorption of
solubilized formazan was measured at 490 nm using a EL340
microplate reader (Bio-Tek Instruments, Inc., Winooski, VT,
USA).
Nuclear staining
A549 cells were stained using a DAPI staining kit
(Nanjing KeyGen Biotech. Co. Ltd., Nanjing, China). Following
exposure to graded AMP concentrations for 48 h, the cells were
washed and incubated with 1–2 μg/ml DAPI working solution at 37°C
for 15 min. The cells were subsequently washed with methanol
solution. Buffer A containing 60% glycerol in 10 mM
phosphate-buffered saline (PBS; pH 7.6) was added to the
suspension. The A549 cells were viewed using an Eclipse Ti Nikon
microscope (Nikon Corporation, Tokyo, Japan).
Mitochondrial membrane potential
A JC-1 fluorescent, lipophilic, cationic probe
(Beyotime Co., Shanghai, China), was used to measure the
mitochondrial membrane potential (Δψm) of the A549 cells, according
to the manufacturer’s instructions. Briefly, the cells exposed to
AMP were incubated with 1× JC-1 staining solution for 20 min at
37°C. The cells were then washed twice with JC-1 staining buffer
and images were captured with the Eclipse Ti Nikon microscope.
Apoptosis assay
A549 cells were labeled with fluorescein
isothiocyanate (FITC)-labeled Annexin V and propidium iodide (PI)
using an Annexin V-FITC apoptosis detection kit (Nanjing KeyGen
Biotech. Co. Ltd.) according to the manufacturer’s instructions.
Briefly, after 48 h exposure to different AMP concentrations, the
cells were washed with cold PBS and then resuspended in 1× binding
buffer. Aliquots of 105 cells were mixed with 5 μl
Annexin V-FITC and 5 μl PI for 15 min at room temperature in the
dark. Fluorescence (530 nm) was detected within 1 h using flow
cytometry at a wavelength of 530 nm (FACS Aria; BD Biosciences,
Franklin Lakes, NJ, USA).
Reverse transcription quantitative
polymerase chain reaction (RT-qPCR) analysis
RT-qPCR with a SYBR® Green reporter was
conducted. The A549 cells exposed to AMP were washed with PBS.
Total RNA was purified using RNAiso Plus (Takara, Dalian, China).
The resultant RNA was first reverse transcribed to cDNA using a
PrimeScript® RT Master Mix kit (Takara). Gene-specific
primers were combined with SYBR® Premix Ex Taq™ (Takara)
and amplified using an ABI 7500 Real-Time PCR System (Applied
Biosystems, Foster City, CA, USA). All qPCR reactions were
conducted independently on five samples. The relative mRNA
expression levels were calculated using the 2−ΔΔCt
method. The primer sequences are described in Table I.
 | Table IPrimer sequences for quantitative
polymerase chain reaction. |
Table I
Primer sequences for quantitative
polymerase chain reaction.
| Gene | Orientation | Primer sequence
(5′-3′) |
|---|
| GAPDH | Forward |
GAAATCCCATCACCATCTTCCAGG |
| Reverse |
GAGCCCCAGCCTTCTCCATG |
| HDAC1 | Forward |
TAAATTCTTGCGCTCCATCC |
| Reverse |
AACAGGCCATCGAATACTGG |
| HDAC2 | Forward |
CGTGTAATGACGGTATCATTCC |
| Reverse |
ACCAGATAATGAGTCTGCACC |
| c-Myc | Forward |
AGCGACTCTGAGGAGGAACAAG |
| Reverse |
GTGGCACCTCTTGAGGACCA |
| Skp2 | Forward |
TGGGAATCTTTTCCTGTCTG |
| Reverse |
GAACACTGAGACAGTATGCC |
| Fbw7α | Forward |
AGTAGTATTGTGGACCTGCCCGTT |
| Reverse |
GACCTCAGAACCATGGTCCAACTT |
| GSK3β | Forward |
GGCAGCATGAAAGTTAGCAGA |
| Reverse |
GGCGACCAGTTCTCCTGAATC |
Western blot analysis
Following washes with PBS, the cells were lysed with
radioimmunoprecipitation assay buffer containing 50 mM Tris (pH
7.4), 150 mM NaCl, 1% nonyl phenoxypolyethoxylethanol-40, 0.5%
sodium deoxycholate, 0.1% sodium dodecyl sulfate (SDS), sodium
orthovanadate, EDTA, sodium fluoride and leupeptin (Beyotime Co.)
supplemented with phenylmethylsulfonyl fluoride protease inhibitor
(Beyotime Co.). Cytoplasmic proteins were separated using a nuclear
and cytoplasmic protein extraction kit (Nanjing KeyGen Biotech. Co.
Ltd.). The concentrations of soluble proteins were determined with
a bicinchoninic acid protein assay kit (Beyotime Co.). The cell
lysates were boiled in loading buffer for 5 min and then separated
on a 15% SDS-PAGE gel. The proteins were subsequently transferred
to a polyvinylidene difluoride membrane (Millipore, Billerica, MA,
USA). The membranes were blocked with Tris-buffered saline-Tween 20
(TBST) containing 5% non-fat milk at room temperature for 1 h, and
were incubated with the aforementioned monoclonal primary
antibodies (1:1,000) overnight at 4°C. Subsequent to washing three
times for 5 min each with 15 ml TBST, the membranes were incubated
with the corresponding horseradish peroxidase-conjugated secondary
antibodies (1:10,000; Beyotime Co.) for 1 h at room temperature and
visualized with enhanced chemiluminescence detection reagents
(Beyotime Co.).
Statistical analysis
Data analysis was performed using SPSS version 17.0
software (SPSS, Inc., Chicago, IL, USA). The results are expressed
as the mean ± standard error of the mean from a minimum of three
independent experiments. The statistical significance between
groups was determined by one-way analysis of variance. P<0.05
was considered to indicate a statistically significant
difference.
Results
AMP inhibits cell growth and induces cell
apoptosis
Cell viability was assessed using an MTT assay
together with DAPI staining and flow cytometric analysis, to
investigate apoptosis following exposure to different
concentrations of AMP. The MTT analysis indicated that AMP
inhibited cell growth in a dose-dependent manner (Fig. 1B) and this was confirmed by the
results of the flow cytometric analysis using Annexin V-PI
(Fig. 1C). As compared with the
control group, the early- and late-stage apoptotic rates increased
subsequent to exposure to all concentrations of AMP. In the 30 μM
AMP group, the total apoptosis rate exceeded 50%. As shown in
Fig. 1D, DAPI staining revealed
signs of condensed and cleaved nuclei in the cells administered 30
μM AMP, but only clear nuclei with pale blue staining were observed
in the control group.
The results also revealed an association between AMP
treatment and a dose-dependent decline in the mitochondrial
potential (Δψm), with reduced red fluorescence (JC-1 polymer) and
increased green fluorescence (JC-1 monomer), as presented in
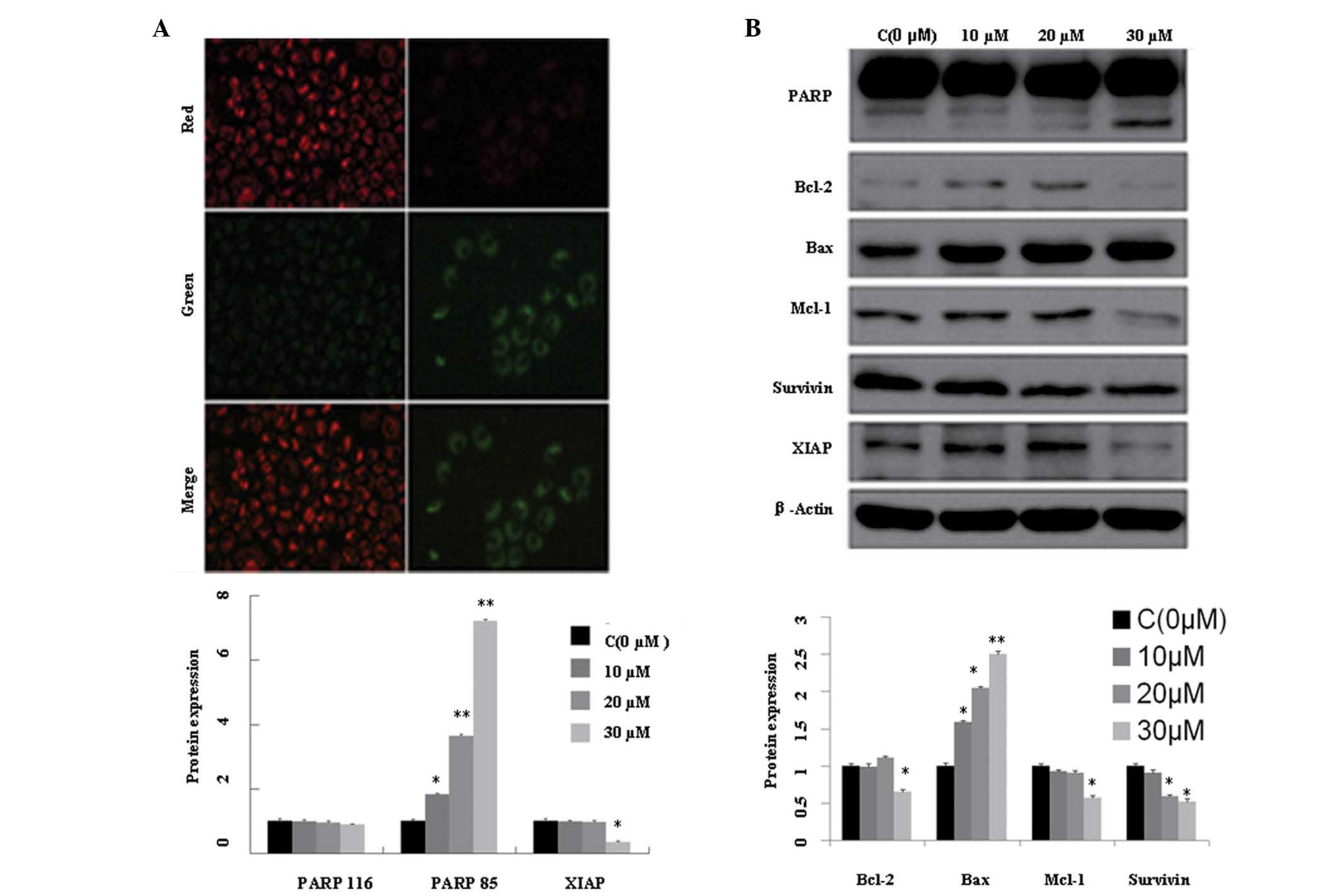
Fig. 2A. This result may have been
due to the downregulation of Mcl-1 and Bcl-2 (Fig. 2B). The upregulation of cleaved
fragments from PARP (PARP 85) and the downregulation of XIAP and
survivin suggest that the apoptotic process continued following
mitochondrial damage.
AMP downregulates HDAC2 at the mRNA and
protein levels
Significant downregulation of HDAC1 and 2 mRNA
following 30 μM AMP treatment, as compared with the control
treatment, was detected by qPCR (Fig.
3A). This result is consistent with the finding that HDAC2
protein expression was downregulated in a dose-dependent manner
following exposure to different concentrations of AMP for 48 h
(Fig. 3B). These changes may
result in a subsequent proportional increase in the levels of
histone acetylation, therefore enhancing the expression levels of
tumor suppressive proteins.
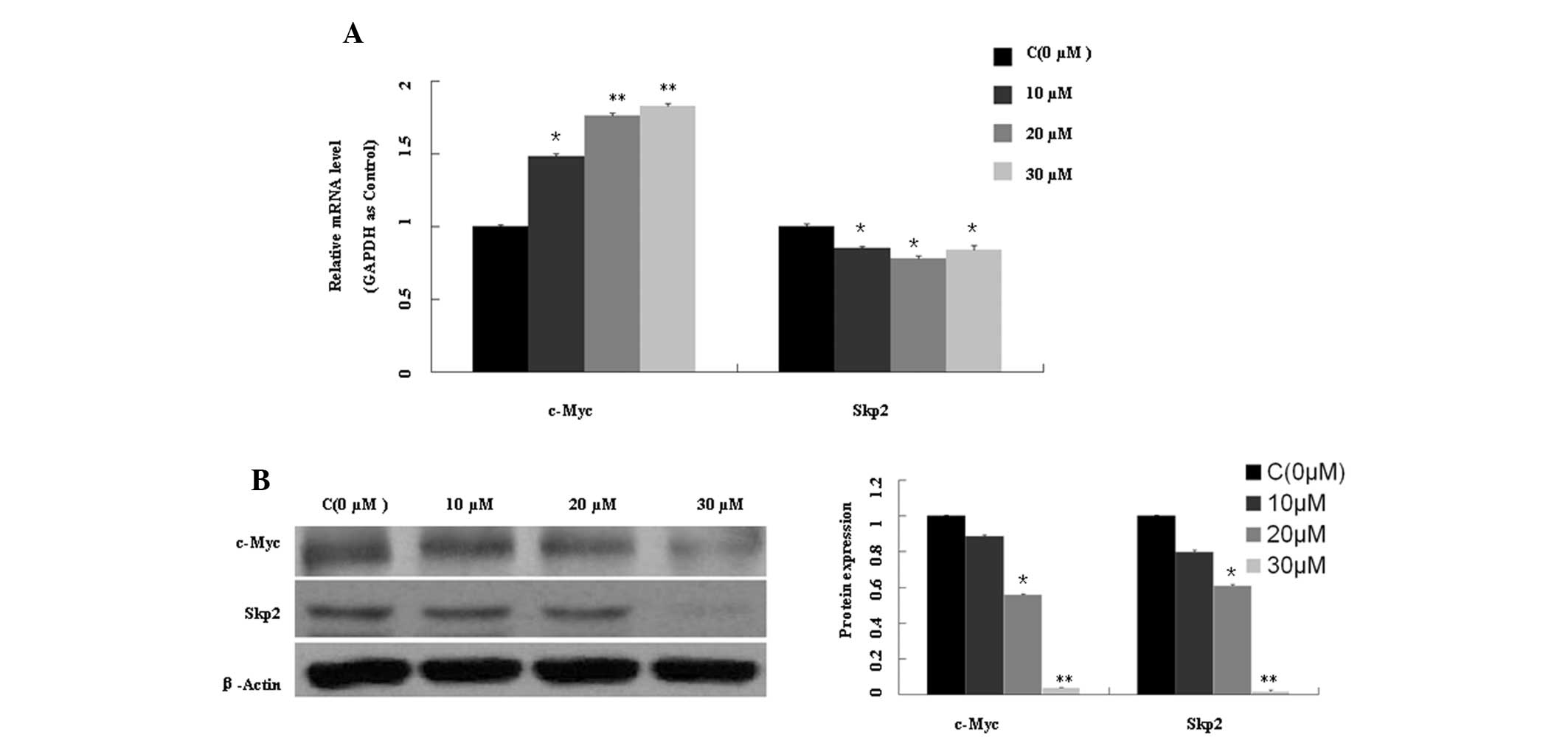
AMP increases c-Myc mRNA expression
levels, but downregulates c-Myc and Skp2 expression at the protein
level
As shown in Fig.
4A, Skp2 mRNA expression levels were significantly reduced
(P<0.05), whereas c-Myc mRNA expression levels were increased up
to ~1.6-fold following exposure to AMP (10, 20 and 30 μM) for 48 h,
as compared with the control treatment. c-Myc and Skp2 were also
downregulated at the protein level (Fig. 4B). These findings indicate that a
proteasomal degradation pathway may be involved in this process.
However, as AMP treatment resulted in reduced Skp2 mRNA expression
levels, a different proteasome recognition subunit, Fbw7 (including
the three isoforms in humans, Fbw7α, -β and -γ), which targets
c-Myc, was subsequently analyzed.
AMP reduces Fbw7α, Fbw7γ and GSK3β
expression, and increases Thr58-Myc expression at the protein
level
Following exposure to different concentrations of
AMP, the Fbw7α and Fbw7γ mRNA expression levels were increased
(Fig. 5A), but with a
corresponding reduction in Fbw7α and Fbw7γ protein expression
levels (Fig. 5B). Thr58
phosphorylation of c-Myc is a requirement for c-Myc degradation and
is mediated by GSK3β. GSK3β is the only kinase that has been
previously demonstrated to phosphorylate c-Myc at Thr58 (35,36).
However, in the present study, GSK3β expression following AMP
treatment was reduced at the mRNA (Fig. 5A) and protein levels (Fig. 5B), as compared with the control
treatment, with a certain degree of increase in Thr58
phosphorylation of c-Myc (Fig.
5B). These findings indicate that the phosphorylation of c-Myc
at Thr58 may occur independently of GSK3β. The process may
therefore involve reduced Fbw7α, Fbw7γ and Skp2 expression levels,
but the exact pathway for c-Myc degradation requires further
investigation.
Discussion
AMP is a naturally occurring flavonoid extracted
from Scutellaria baicalensis Radix, which has been reported
to exert antineoplastic activity in various types of cancer
(28–34). In the present study, the roles of
the c-Myc/Skp2/Fbw7 and HDAC1/2 pathways, which are associated with
tumor progression, on the anticancer effects of AMP in the A549
NSCLC line were investigated. The effects of AMP upon A549 cell
proliferation and apoptosis were evaluated using MTT assays and
Annexin V-PI double staining. The results indicated that AMP
inhibited cell growth at half maximal inhibitory concentration
<30 μM in a dose-dependent manner. In response to AMP treatment,
the early-stage apoptotic rate was increased, since Annexin
V-positive cells gradually became Annexin V-negative. Significant
apoptosis was observed following 30 μM AMP treatment, as compared
with the control treatment, a finding consistent with the
morphologic changes observed subsequent to cell nuclear DAPI
staining. As shown in Fig. 1D,
condensed and cleaved nuclei were detected only in the treatment
groups.
To identify the apoptotic effects of AMP treatment
at the protein level, the expression levels of apoptosis-related
proteins, including Mcl-1, PARP, survivin, Bcl-2 and XIAP were
assessed. The results indicated that AMP influenced mitochondrial
membrane stability and reduced the mitochondrial membrane potential
(Δψm; Fig. 2A). This process may
be associated with the reduced Bcl-2 and increased Bax expression
levels observed (Fig. 2B). Cleaved
PARP and reduced XIAP and survivin expression levels may also have
partially contributed to the progression of apoptosis.
HDAC1 and 2, Class I HDACs, not only deacetylate
histone/non-histone proteins, but also inhibit gene expression and
modify tumor suppressive proteins associated with tumor progression
(3,4). The results of the present study
revealed that the expression levels of HDAC1/2 were reduced at the
mRNA and protein levels in the presence of AMP (Fig. 3), indicating that acetylated
histone proteins may promote the expression of tumor suppressive
proteins and thus inhibit tumor progression.
A previous study demonstrated that c-Myc, as a
transcription factor, has the ability to regulate various genes
involved in cellular proliferation, differentiation, growth and
apoptosis (7). The
post-translational regulation of c-Myc is mediated by Skp2 and Fbw7
(12). The c-Myc and Skp2
oncoproteins are inter-related such that c-Myc promotes Skp2
expression and Skp2 targets c-Myc for ubiquitin-dependent
degradation. In the present study, AMP downregulated Skp2
expression at the mRNA level and protein levels. In addition, AMP
downregulated c-Myc at the protein level, but c-Myc mRNA expression
levels were increased (Fig. 4).
These findings suggest that the proteasomal degradation pathway may
be associated with the reversal of c-Myc. However, since Skp2
expression levels were reduced, subsequent experiments were focused
on Fbw7 (which has three isoforms in humans: Fbw7α, β and γ),
another proteasome recognition subunit that targets c-Myc.
The involvement of Fbw7 deficiency in drug
resistance in human cancer has been recently identified (22,23).
In the regulation of c-Myc, Fbw7 is different from Skp2. Thr58
phosphorylation of c-Myc is required for Fbw7α, β and γ, which have
been shown to mediate c-Myc ubiquitin-dependent degradation
(25), and GSK3β is the only known
kinase that phosphorylates c-Myc at Thr58 (24). In the present study, the expression
levels of Fbw7α, Fbw7β, Fbw7γ, p-c-Myc (Thr58) and GSK3β were
evaluated.
Fbw7α and Fbw7γ expression following AMP treatment
were found to be reduced at the protein level, but increased at the
mRNA level (Fig. 5). Thr58
phosphorylation of c-Myc was increased to a certain degree, and
GSK3β expression at the mRNA and protein levels was reduced. These
results highlight the lack of conformity between Fbw7α/γ mRNA and
protein levels, the reduced expression levels of GSK3β and the
increased phosphorylation of c-Myc at Thr58. These findings suggest
that AMP induces apoptosis independently of Fbwα/γ and that the
phosphorylation of c-Myc at Thr58 may occur independently of GSK3β,
a finding concurrent with those of previous studies that observed
phosphorylation of Thr58 in A549 cells occurring independently of
GSK3β (37,38).
In conclusion, the results of the present study
indicate that AMP influences multiple biochemical pathways, which
may explain AMP activity against various types of cancer. In
addition, the results may also partially explain why cells
deficient in certain genes, for instance Fbw7-deficient cells, are
resistant to AMP and suggest a possible use of AMP in
drug-resistant cancer associated with Fbw7 deficiency. However, the
exact underlying mechanism of AMP action, which pathways are
associated with c-Myc and Fbw7α/γ reversal and whether Thr58
phosphorylation of c-Myc is dependent on GSK3β requires further
investigation.
Acknowledgements
This study was supported by the National Science
Foundation of China (no. 81372466); the Ministry of Science and
Technology of Guizhou Province, China [nos. (2012) 7006 and NY
(2011)3072]; and the Foundation for Young Scientists of Guangzhou
Educational Committee (no. 2012C118).
References
|
1
|
Juergens R and Brahmer J: Targeting the
epidermal growth factor receptor in non-small-cell lung cancer:
who, which, when, and how? Curr Oncol Rep. 4:255–264. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Crocetti E and Paci E: Trends in lung
adenocarcinoma incidence and survival. Lung Cancer. 2:215–216.
2002. View Article : Google Scholar
|
|
3
|
Ruthenburg AJ, Li H, Patel DJ and Allis
CD: Multivalent engagement of chromatin modifications by linked
binding modules. Nat Rev Mol Cell Biol. 8:983–994. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Marks PA and Xu WS: Histone deacetylase
inhibitors: Potential in cancer therapy. J Cell Biochem.
107:600–608. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Slingerland M, Guchelaar HJ and Gelderblom
H: Histone deacetylase inhibitors: an overview of the clinical
studies in solid tumors. Anticancer Drugs. 2:140–149. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
de Ruijter AJ, van Gennip AH, Caron HN,
Kemp S and van Kuilenburg AB: Histone deacetylases (HDACs):
characterization of the classical HDAC family. Biochem J.
370:737–749. 2003.PubMed/NCBI
|
|
7
|
Dang CV, Resar LM, Emison E, Kim S, Li Q,
Prescott JE, Wonsey D and Zeller K: Function of the c-Myc oncogenic
transcription factor. Exp Cell Res. 253:63–77. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Nesbit CE, Tersak JM and Prochownik EV:
MYC oncogenes and human neoplastic disease. Oncogene. 18:3004–3016.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Flinn EM, Busch CM and Wright AP: Myc
boxes, which are conserved in Myc family proteins, are signals for
protein degradation via the proteasome. Mol Cell Biol.
18:5961–5969. 1998.PubMed/NCBI
|
|
10
|
Jones TR and Cole MD: Rapid cytoplasmic
turnover of c-Myc mRNA: requirement of the 3′ untranslated
sequences. Mol Cell Biol. 7:4513–4521. 1987.PubMed/NCBI
|
|
11
|
Kelly K, Cochran H, Stiles CD and Leder P:
Cell-specific regulation of the c-Myc gene by lymphocyte mitogens
and platelet-derived growth factor. Cell. 35:603–610. 1983.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Amati B: Myc degradation: Dancing with
ubiquitin ligases. Proc Natl Acad Sci USA. 101:8843–8844. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Carrano AC, Eytan E, Hershko A and Pagano
M: Skp2 is required for ubiquitin-mediated degradation of the CDK
inhibitor p27. Nat Cell Biol. 1:193–199. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kamura T, Hara T, Kotoshiba S, Yada M,
Ishida N, Imaki H, Hatakeyama S, Nakayama K and Nakayama KI:
Degradation of p57 Kip2 mediated by SCFSkp2-dependent
ubiquitylation. Proc Natl Acad Sci USA. 100:10231–10236. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Tedesco D, Lukas J and Reed SI: The
pRb-related protein p130 is regulated by phosphorylation-dependent
proteolysis via the protein-ubiquitin ligase SCF (Skp2). Genes Dev.
16:2946–2957. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hiramatsu Y, Kitagawa K, Suzuki T, Uchida
C, Hattori T, Kikuchi H, Oda T, Hatakeyama S, Nakayama KI, Yamamoto
T, et al: Degradation of Tob1 mediated by SCFSkp2-dependent
ubiquitination. Cancer Res. 66:8477–8483. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhao D, Zheng HQ, Zhou Z and Chen C: The
Fbw7 tumor suppressor targets KLF5 for ubiquitin-mediated
degradation and suppresses breast cell proliferation. Cancer Res.
70:4728–4738. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Akhoondi S, Lindström L, Widschwendter M,
Corcoran M, Bergh J, Spruck C, Grandér D and Sangfelt O:
Inactivation of FBXW7/hCDC4-beta expression by promoter
hypermethylation is associated with favorable prognosis in primary
breast cancer. Breast Cancer Res. 12:R1052010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Inuzuka H, Shaik S, Onoyama I, et al:
SCF(FBW7) regulates cellular apoptosis by targeting MCL1 for
ubiquitylation and destruction. Nature. 471:104–109. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sancho R, Jandke A, Davis H, Diefenbacher
ME, Tomlinson I and Behrens A: F-box and WD repeat
domain-containing 7 regulates intestinal cell lineage commitment
and is a haploinsufficient tumor suppressor. Gastroenterology.
139:929–941. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
O’Neil J and Look AT: Mechanisms of
transcription factor deregulation in lymphoid cell transformation.
Oncogene. 26:6838–6849. 2007.PubMed/NCBI
|
|
22
|
Wang Z, Fukushima H, Gao D, Inuzuka H, Wan
L, Lau AW, Liu P and Wei W: The two faces of FBW7 in cancer drug
resistance. Bioessays. 11:851–859. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wertz IE, Kusam S, Lam C, et al:
Sensitivity to antitubulin chemotherapeutics is regulated by MCL1
and FBW7. Nature. 471:110–114. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Pulverer BJ, Fisher C, Vousden K,
Littlewood T, Evan G and Woodgett JR: Site-specific modulation of
c-Myc cotransformation by residues phosphorylated in vivo.
Oncogene. 9:59–70. 1994.PubMed/NCBI
|
|
25
|
Kim SY, Herbst A, Tworkowski KA, Salghetti
SE and Tansey WP: Skp2 regulates Myc protein stability and
activity. Mol Cell. 11:1177–1188. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Spruck CH, Strohmaier H, Sangfelt O,
Müller HM, Hubalek M, Müller-Holzner E, Marth C, Widschwendter M
and Reed SI: hCDC4 gene mutations in endometrial cancer. Cancer
Res. 62:4535–4539. 2002.PubMed/NCBI
|
|
27
|
Welcker M, Orian A, Grim JE, Eisenman RN
and Clurman BE: A nucleolar isoform of the Fbw7 ubiquitin ligase
regulates c-Myc and cell size. Curr Biol. 14:1852–1857. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhang B, Dong S, Cen X, Wang X, Liu X,
Zhang H, Zhao X and Wu Y: Ampelopsin sodium exhibits antitumor
effects against bladder carcinoma in orthotopic xenograft models.
Anticancer Drugs. 6:590–596. 2012. View Article : Google Scholar
|
|
29
|
Zheng HQ and Liu DY: Anti-invasive and
anti-metastatic effect of ampelopsin on melanoma. Ai Zheng.
22:363–367. 2003.(In Chinese).
|
|
30
|
Zeng S, Liu D, Ye Y, Wang L and Wang W:
Anti-tumor effects of ampelopsin on human lung cancer GLC-82
implanted in nude mice. Zhong Yao Cai. 27:842–845. 2004.(In
Chinese).
|
|
31
|
Luo GQ, Zeng S and Liu DY: Inhibitory
effects of ampelopsin on angiogenesis. Zhong Yao Cai. 29:146–150.
2006.(In Chinese).
|
|
32
|
Ye J, Zheng Y and Liu D: Reversal effect
and its mechanism of ampelopsin on multidrug resistance in K562/ADR
cells. Zhongguo Zhong Yao Za Zhi. 34:761–765. 2009.(In
Chinese).
|
|
33
|
Ni F, Gong Y, Li L, Abdolmaleky HM and
Zhou JR: Flavonoid ampelopsin inhibits the growth and metastasis of
prostate cancer in vitro and in mice. PLoS One. 7:e388022012.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kou X, Shen K, An Y, Qi S, Dai WX and Yin
Z: Ampelopsin inhibits H2O2-induced apoptosis
by ERK and Akt signaling pathways and up-regulation of heme
oxygenase-1. Phytother Res. 7:988–994. 2012.
|
|
35
|
Gregory MA, Qi Y and Hann SR:
Phosphorylation by glycogen synthase kinase-3 controls c-myc
proteolysis and subnuclear localization. J Biol Chem.
51:51606–51612. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kamemura K, Hayes BK, Comer FI and Hart
GW: Dynamic interplay between O-glycosylation and O-phosphorylation
of nucleocytoplasmic proteins: alternative
glycosylation/phosphorylation of THR-58, a known mutational hot
spot of c-Myc in lymphomas, is regulated by mitogens. J Biol Chem.
21:19229–19235. 2002. View Article : Google Scholar
|
|
37
|
Chen XM, Bai Y, Zhong YJ, Xie XL, Long HW,
Yang YY, Wu SG, Jia Q and Wang XH: Wogonin has multiple anti-cancer
effects by regulating c-Myc/SKP2/Fbw7a and HDAC1/HDAC2 pathways and
inducing apoptosis in human lung adenocarcinoma cell line A549.
PLoS One. 8:e792012013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Li Q, Kluz T, Sun H and Costa M:
Mechanisms of c-Myc degradation by nickel compounds and hypoxia.
PLoS One. 4:e85312009. View Article : Google Scholar : PubMed/NCBI
|