Introduction
Cancer is predominantly caused by mutations,
germ-line or somatic, without any separation of cell type observed
for a long period. It is known that certain stem cells are
associated with the occurrence of cancer. These cells are called
cancer stem cells because of their tumor-initiating and drug
resistance properties (1).
Isolation of cancer stem cells from hepatocellular carcinoma (HCC)
was previously reported (2).
A number of methods have been used to study cancer
stem cells, including cell sorting using specific surface markers
(3), culture of sphere-forming
cells (4) and analyses of the side
population (SP) (5), defined as
cells capable of efficient Hoechst 33342 dye efflux (6). SP cells can be used as a model to
study stem cells because of their self-renewing, differentiating
(7) and tumor-initiating abilities
(8). SP cells identified in liver
cancer initiated tumors (9),
expressed high levels of ABCG2 transporters (10), and expressed a marker of
hepatocarcinoma stem cells, CD133. Because of these properties,
these SP cells were suggested to be liver cancer stem cells
(9), and to associate with the
high drug resistance of liver cancer. Treatment with anticancer
drugs such as doxorubicin, 5-fluorouracil (5-FU), and gemcitabine,
increases the SP cell number. In addition, SP cells are known to
relate to the drug resistance of liver cancer associated with
transporters (11).
c-Jun-N-terminal kinase (JNK) signaling in SP cells
was previously associated with drug resistance in liver cancer
(12). JNK signaling maintains
proliferation and migration in healthy stem cells (13), and is essential for liver
regeneration processes (14). In
contrast to healthy liver cells, JNK signaling inhibits survival
and growth in liver cancer cells upon activation of c-Jun (15). To date, the relationship between SP
cells, drug resistance, and JNK signaling in liver cancer is
unclear.
In this study, we used the hepatocarcinoma cell
lines Huh7 and HepG2 to study the relationship between increased
numbers of SP cells and the signaling molecules involved in this
interaction. We analyzed the fraction of SP cells during and after
treatment with anticancer drugs, and examined cell survival rates
and expression of relevant signaling molecules.
Material and methods
Cell cultures
Huh7 and HepG2 cells were obtained from the Korean
Cell Line Bank (Seoul, Korea). The cells were cultured in
Gibco® Dulbecco’s modified Eagle’s medium (DMEM)
containing 10% Gibco® fetal bovine serum (FBS), both
purchased from Thermo Fisher Scientific (Waltham, MA, USA). To
obtain cultures of adherent cells, 5×105 cells were
seeded on Falcon® tissue culture dishes (Corning,
Tewksbury, MA, USA). All cultures were maintained at 37°C in a
humidified 5% CO2 atmosphere.
Cell survival rates after anticancer drug
treatment
Huh7 cells were seeded (5×105 cells) in
DMEM containing 10% FBS. After 24 h, the cells were washed twice
with phosphate-buffered saline (PBS; Gibco-BRL, Carslbad, CA, USA)
and fresh medium was added. The cells were treated for 72 h with
distilled water (control), 1, 10 or 100 μM 5-FU, or 1, 10 or 100 nM
paclitaxel (both from Sigma-Aldrich, St. Louis, MO, USA). The
survival rate was estimated as the number of viable cells, counted
in a Neubauer chamber (Millipore, Temecula, CA, USA) at different
time-points, as the cells that were positively stained with the
Trypan blue dye (0.4%; Gibco-BRL). Selected cells treated with 10
μM 5-FU or 10 nM paclitaxel were used to observe SP cells and JNK
expression following drug treatment.
SP cell analyses
Huh7 cells were seeded as described above. The cells
were treated with distilled water (control), 10 μM 5-FU, or 10 nM
paclitaxel for 72 h. SP cell analyses were performed as previously
described (16). Briefly, cells
were detached and collected as cell pellets. To analyze the SP
fraction, 1×106 cells/ml were incubated with the Hoechst
33342 dye (5 μg/ml; Sigma-Aldrich, St. Louis, MO, USA) in DMEM
containing 10% FBS, with vortexing for 90 min at 37°C. Cells were
also incubated with the Hoechst dye and 50–100 μM verapamil
(Sigma-Alrdich), an efflux blocker, to confirm the SP cell
population. At the end of the incubation, cells were centrifuged at
320 × g at 4°C and collected for analyses of the SP fraction.
Propidium iodide (1 μg/ml; Sigma-Aldrich) was added prior to
fluorescence-activated cell sorting (FACS) analysis, in order to
allow identification and exclusion of dead cells. Samples were
analyzed using a FACS Aria instrument (BD Biosciences, Franklin
Lakes, NJ, USA).
Phospho-stress-activated protein kinase
(p-SAPK)/JNK immunoblotting
Huh7 cells were seeded as described above. The cells
were treated for 72 h with distilled water (control), 10 μM 5-FU,
or 10 nM paclitaxel. Total cell lysates were prepared in 100 μl
Cell Lysis buffer (Cell Signaling Technology, Beverly, MA, USA).
Protein concentrations were measured using a Protein Assay kit
(Bio-Rad Laboratories, Hercules, CA, USA). Equal amounts of cell
lysates were separated using 10% sodium dodecyl
sulfate-polyacrylamide gel electrophoresis (SDS-PAGE), and proteins
were electrotransferred onto Hybond-ECL nitrocellulose membranes
(Amersham Biosciences, Buckinghamshire, UK). Blots were blocked for
1 h with blocking buffer and incubated overnight at 4°C with mouse
monoclonal anti-p-SAPK/JNK (dilution, 1:1,000, Thr183/Tyr185; Cell
Signaling Technology) and anti-β-actin (dilution, 1:1,000; Santa
Cruz Biotechnology, Inc., Santa Cruz, CA, USA) immunoglobulin G
(IgG). Blots were washed with 0.02% Tris-buffered saline with
Tween20 (Sigma-Alrdich) and incubated for 1 h at room temperature
with goat anti-mouse IgG (H+L; 1:2,500; Jackson ImmunoResearch
Laboratories, West Grove, PA, USA). Labeled proteins were detected
using an enhanced chemiluminescence detection system (Amersham
Biosciences).
p-SAPK/JNK, c-Jun and phospho-c-Jun
(p-c-Jun) expression after treatment with anticancer drugs and
SP600125
Huh7 and HepG2 cells were seeded as described above.
Cells were treated for 72 h with distilled water and dimethyl
sulfoxide (DMSO; control; Sigma-Aldrich), 10 μM SP600125
(Sigma-Aldrich), 10 μM 5-FU, or 10 μM SP600125 and 10 μM 5-FU. The
SP fraction, cell survival, expression of p-SAPK/JNK, c-Jun, and
p-c-Jun were measured as described above. c-Jun and p-c-Jun
expression were analyzed using c-Jun and p-c-Jun mouse monoclonal
IgGs (both at 1:1,000; Cell Signaling Technology).
HCC tissue immunohistochemistry
The Institutional Review Board of the Seoul National
University Hospital approved the use of human tissues. The tissues
were collected from six liver cancer patients who had surgery at
the Seoul National University Hospital. Tissues were embedded with
paraffin, cut into 3-μm sections, and deparaffinized with xylene.
Antigen retrieval was performed by blocking with PBS containing 10%
donkey serum for 1 h. Tissues were incubated overnight at 4°C with
the p-SAPK/JNK and p-c-Jun mouse monoclonal IgGs. Sections were
incubated for 1 h at 37°C with the biotinylated anti-mouse
secondary antibody (Vector Laboratories, Burlingame, CA, USA). The
Vectastain Elite ABC reagent (Vector Laboratories) was added, and
the formed antibody/enzyme conjugate was visualized with addition
of 3,3′-diaminobenzidine (DAB; Vector Laboratories). All sections
were counterstained with hematoxylin and eosin (Sigma-Alrdich). For
these analyses, six different HCC samples were evaluated per
experimental condition.
Statistical analyses
A minimum of three replicates were performed for all
experiments. Data were expressed as the mean ± standard error.
Comparisons between treated and control cell data were performed
using Student’s t-tests. P-values <0.05 was considered to
indicate statistically significant differences.
Results
Cell survival, SP cell fraction, and
p-SAPK/JNK expression upon 5-FU and paclitaxel treatment
To observe the effect of anticancer drugs, 5-FU or
paclitaxel were administered to Huh7 cells for 72 h, and the cell
survival was assessed by cell counting after trypan blue staining.
Huh7 cells exhibited a dose-dependent inhibition in survival
following treatment with the anticancer drugs (Fig. 1, upper panels). The survival of
Huh7 cells relative to that of untreated cells was 63% following
treatment with 1 μM 5-FU 32% with 10 μM, and 22% with 100 μM
(Fig. 1A, upper panel). The
survival of Huh7 cells relative to that of untreated cells was 73%
after treatment with 1 nM paclitaxel, 57% with 10 nM, and 8% with
100 nM (Fig. 1B, upper panel).
Based on these results, we selected the doses of 10 μM 5-FU and 10
nM paclitaxel to observe SP cells and the expression of JNK upon
anticancer drug treatment.
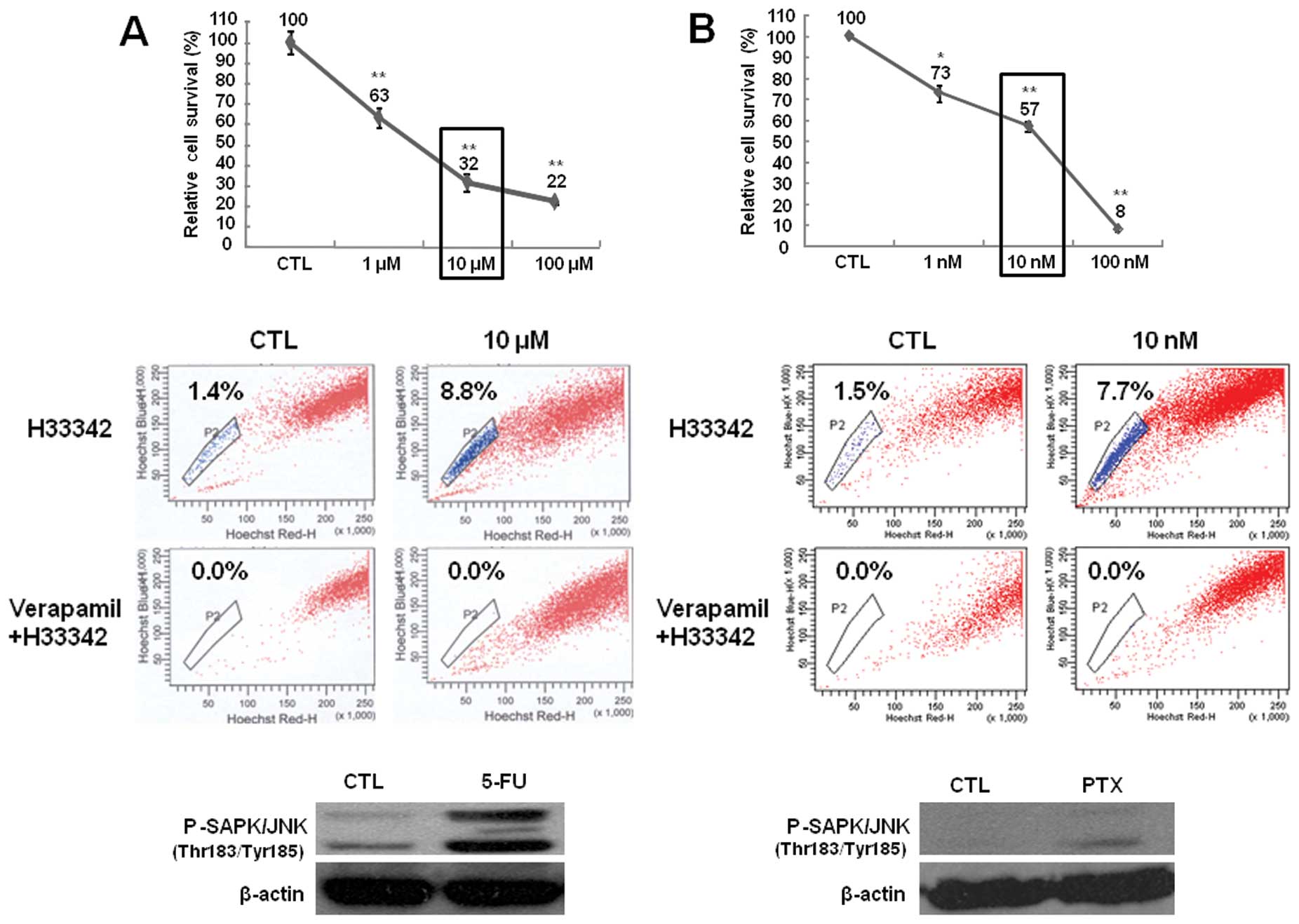 | Figure 1Cell survival rate, fraction of side
population (SP) cells, and phospho-stress-activated protein
kinase/c-Jun-N-terminal kinase (p-SAPK/JNK) expression in Huh7
cells after treatment with (A) 5-fluorouracil (5-FU) and (B)
paclitaxel. Upper panels: cell survival was assessed by counting
cells using trypan blue dye exclusion. Cells were treated with
distilled water (CTL), 1, 10 or 100 μM 5-FU, or 1, 10 or 100 nM
paclitaxel for 72 h. Survival rate is expressed as a percentage
relative to that of control cells. *P<0.05,
**P<0.01 compared with the control and treated cells,
respectively. Middle panels: for each group of cells, the
percentage of SP cells is shown in the upper quadrant. Cells were
treated with distilled water (CTL), 10 μM 5-FU, or 10 nM paclitaxel
for 72 h. Lower panels: p-SAPK/JNK expression was detected by
immunoblotting. Cells were treated with distilled water (CTL), 10
μM 5-FU, or 10 nM paclitaxel for 72 h. Representative blots (n=3)
are shown. β-actin was used as the loading control. |
Cells were treated with 10 μM 5-FU or 10 nM
paclitaxel for 72 h. Drug treatment increased the SP cell number
relative to untreated cells (Fig.
1, middle panels). The SP cell fraction was 1.4% in untreated
cells, 2.6% in 1 μM 5-FU-treated cells, 8.8% in 10 μM 5-FU-treated
cells, and 3.9% in 100 μM 5-FU-treated cells (Fig. 1A, middle panel). The SP cell
fraction was 1.5% in untreated cells, 2.4% in 1 nM
paclitaxel-treated cells, 7.7% in 10 nM paclitaxel-treated cells,
and 1.6% in 100 nM paclitaxel-treated cells (Fig. 1B, middle panel). In addition, we
observed JNK activation upon drug treatment. Both 5-FU and
paclitaxel increased the level of p-SAPK/JNK (Fig. 1, lower panels). These results
indicate that the increase in the SP cell fraction may be
associated with JNK signaling.
The SP cell fraction decreases upon
treatment with 5-FU and SP600125
To determine whether the increased number of SP
cells observed after treatment with anticancer drugs can be reduced
by inhibiting JNK, the SP fraction was evaluated in Huh7 and HepG2
cells treated for 72 h with 5-FU, SP600125 alone, or with both 5-FU
and SP600125. The SP fraction increased upon 5-FU treatment, and
this increase was blocked by SP600125 (Fig. 2). These results indicate that
inhibiting JNK signaling can block the increase in SP cells induced
by anticancer drugs in liver cancer.
The JNK inhibitor SP600125 inhibits
expression of p-SAPK/JNK upon 5-FU treatment
Treatment with anticancer drugs decreased the
survival rate and increased the level of p-JNK and the number of SP
cells. However, the increase in SP cells was blocked by the JNK
inhibitor. To examine whether the JNK inhibitor also enhances cell
survival or blocks upregulation of p-JNK, the relative survival
rate and the JNK expression level were evaluated in Huh7 and HepG2
cells treated for 72 h with 5-FU, SP600125, or both 5-FU and
SP600125. As shown in Fig. 3A,
cell survival was decreased in all treated groups (5-FU, SP600125,
or 5-FU + SP600125). SP600125 did not revert the inhibition on cell
survival induced by 5-FU (Fig.
3A). However, SP600125 attenuated the 5-FU-mediated
upregulation of p-SAPK/JNK, c-Jun, and p-c-Jun (Fig. 3B). These results indicate that the
JNK inhibitor blocks the activation of JNK signaling mediated by
5-FU.
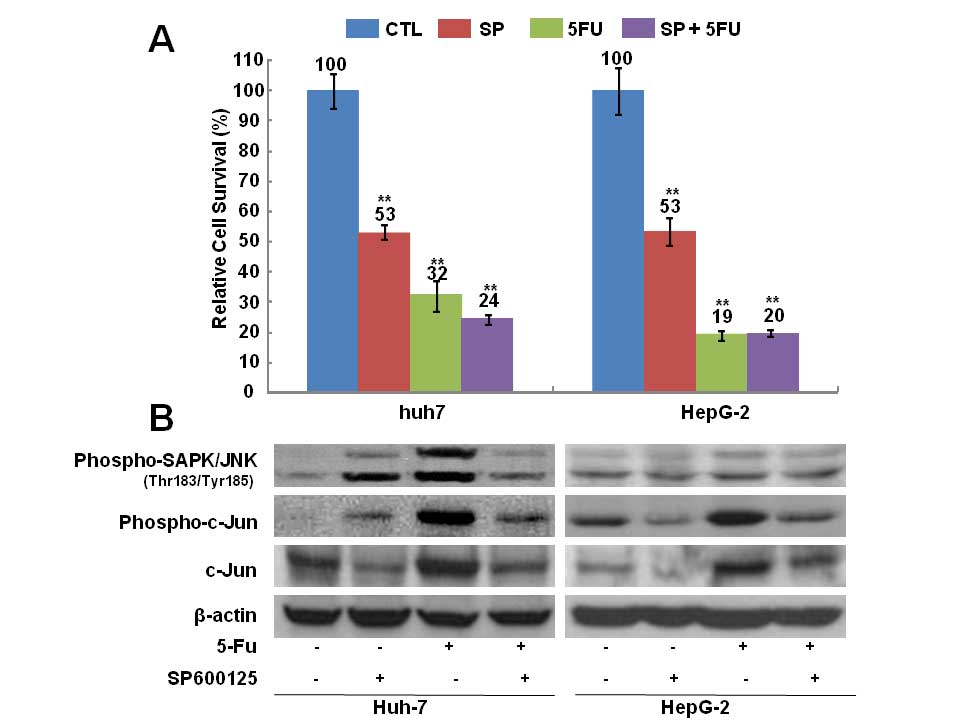 | Figure 3Cell survival rate and expression of
phospho-stress-activated protein kinase/c-Jun-N-terminal kinase
(p-SAPK/JNK), p-c-Jun, and c-Jun in Huh7 and HepG2 cells after
5-fluorouracil (5-FU) and SP600125 treatment. (A) Cell survival was
assessed by counting cells using trypan blue dye exclusion. Cells
were treated for 72 h with distilled water and dimethyl sulfoxide
(DMSO; CTL), 10 μM 5-FU, 10 μM SP600125 (SP), or 10 μM 5-FU + 10 μM
SP600125. Survival rate is expressed as a percentage relative to
that of control cells. Values represent the mean ± standard error
from at least three independent experiments. **P≤0.05.
(B) Expression of p-JNK, p-c-Jun, c-Jun was detected by
immunoblotting. Cells were treated with distilled water and DMSO
(lane 1 from left), 10 μM SP600125 (lane 2), 10 μM 5-FU (lane 3),
and 10 μM SP600125 + 10 μM 5-FU (lane 4). β-actin was used as the
loading control. |
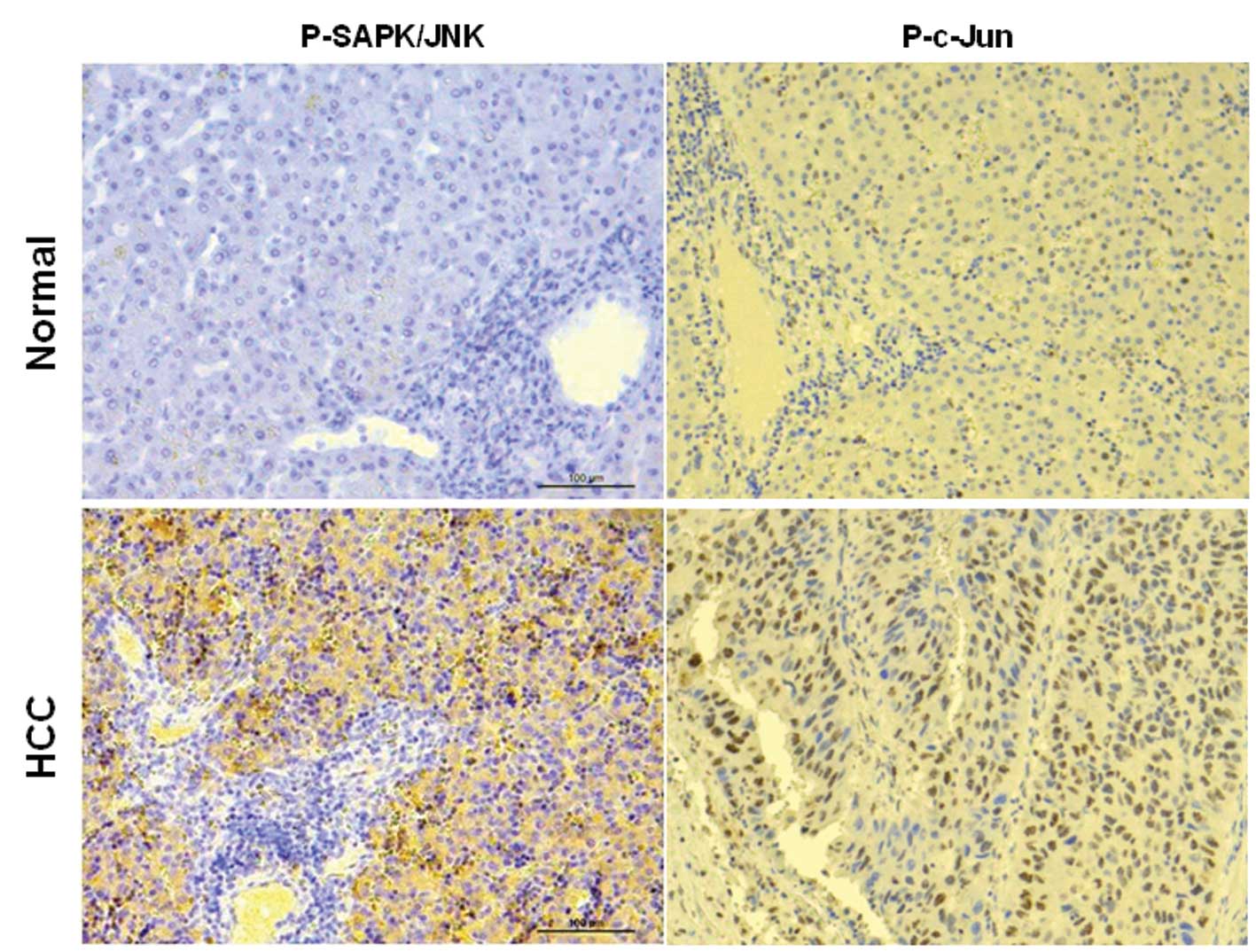
Expression of p-SAPK/JNK and p-c-Jun in
healthy and hepatocarcinoma tissues
p-SAPK/JNK and p-c-Jun expression was found to be
increased upon treatment with anticancer drugs and blocked by the
JNK inhibitor in the Huh-7 and HepG2 hepatocarcinoma cell lines. To
determine the relationship between p-SAPK/JNK and p-c-Jun
expression and the progression of hepatocarcinoma, we evaluated
p-SAPK/JNK and p-c-Jun expression in hepatocarcinoma tissues using
immunohistochemistry. Expression of p-SAPK/JNK and p-c-Jun appeared
high in hepatocarcinoma tissues (Fig.
4). These results indicate that JNK signaling may associate
with hepatocarcinoma progression.
Discussion
Numerous studies have evaluated the increase in
cancer stem cell number upon treatment with anticancer drugs,
focusing on the relationship between drug resistance and cancer
stem cells. In this study, we examined the SP cells, a model for
the study of cancer stem cells, and the involvement of JNK
signaling in liver cancer. To determine the relationship between
cancer stem cells and JNK signaling after treatment with anticancer
drugs, we examined the survival rate, SP cell ratio, and JNK
expression in hepatocarcinoma cell lines after anticancer drug
treatment. We observed that SP cells constitute a small
subpopulation involved in drug resistance in liver cancer, and that
the tested anticancer drugs decreased the survival rate and
increased the SP fraction in two hepatocarcinoma cell lines.
Similar to our results, a number of studies have
suggested that anticancer drugs do not target cancer stem cells,
but instead increase the cancer stem cell fraction in liver cancer.
Bonnet and Dick (17) reported
that cancer cells include a small number of cells within a
heterogeneous population, which have the ability to initiate tumor
formation. Regarding drug resistance induced by cancer cell
culturing, Sharma et al (18) reported that a small population of
cancer cells exhibiting drug resistance propagated and increased in
size after a long period of anticancer drug withdrawal. The authors
suggested that acquisition of drug resistance and the maintenance
of this drug-tolerant population involved activation of IGF-1R
signaling and a distinct chromatin state (18).
To investigate the signaling pathways involved in
the increased SP cell number, we evaluated the expression of JNK
signaling molecules in hepatocarcinoma cell lines following
treatment with anticancer drugs. JNK is also the main signaling
pathway involved in maintaining the stemness of glioma cells
(19). We found that the tested
anticancer drugs increase the expression level of p-JNK in
hepatocarcinoma cell lines, similar to a previous study (19). JNK expression was increased upon
anticancer drug treatment and in hepatocarcinoma tissues. Based on
these results, we simultaneously treated cells with a JNK inhibitor
and the anticancer drugs and observed a blocking effect on the
increased SP cell ratio and JNK expression. Similar to our results,
another study indicated that JNK signaling is associated with the
maintenance of cancer stem cells and drug resistance (13). With respect to the relationship
between JNK signaling and drug resistance in liver cancer, Yan
et al (12) suggested that
JNK1, JNK2, and JNK3 maintain drug resistance by upregulating the
P-glycoprotein. Nagata et al (20) also suggested that JNK signaling is
involved in inhibiting hepatocarcinoma progression based on
experiments using a JNK inhibitor (SP600125) and an animal model.
Similar to our results, Mucha et al (21) showed that the JNK inhibitor
(SP600125) inhibits cell growth and JNK activity.
JNK signaling is known to be involved in cancer
progression in liver cancer (22).
We found that JNK and c-Jun are highly expressed in hepatocarcinoma
compared to healthy tissues. There is a number of reports on the
expression of JNK in hepatocarcinoma. Wang et al (23) demonstrated that high p-p38 and low
p-JNK expression is associated with poor survival in
hepatocarcinoma tissues. Hagiwara et al (24) suggested that the high levels of JNK
and c-Jun may be predictive biomarkers for the high risk of
recurrence after surgery in non-cancerous liver tissue. These
results indicate that JNK activity is involved in liver cancer
progression.
In conclusion, we showed that JNK activity is
associated with an increase in the fraction of the side population
cells upon treatment with anticancer drugs. Side population cells
may be regulated by JNK signaling following anticancer drug
treatment. JNK activity is involved in liver cancer progression.
These results suggest that JNK signaling may be involved in the
maintenance of cancer stem cells and the progression of liver
cancer.
Acknowledgements
This study was supported by grants from the Hanmi
Pharmatheutical Company (no. 800-20130024) and the Yunhan
Corporation (no. 800-20130029).
References
|
1
|
Reya T, Morrison SJ, Clarke MF and
Weissman IL: Stem cells, cancer, and cancer stem cells. Nature.
414:105–111. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Suetsugu A, Nagaki M, Aoki H, Motohashi T,
Kunisada T and Moriwaki H: Characterization of CD133+
hepatocellular carcinoma cells as cancer stem/progenitor cells.
Biochem Biophys Res Commun. 351:820–824. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Al-Hajj M, Wicha MS, Benito-Hernandez A,
Morrison SJ and Clarke MF: Prospective identification of
tumorigenic breast cancer cells. Proc Natl Acad Sci USA.
100:3983–3988. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ponti D, Costa A, Zaffaroni N, Pratesi G,
Petrangolini G, Coradini D, Pilotti S, Pierotti MA and Daidone MG:
Isolation and in vitro propagation of tumorigenic breast cancer
cells with stem/progenitor cell properties. Cancer Res.
65:5506–5511. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Setoguchi T, Taga T and Kondo T: Cancer
stem cells persist in many cancer cell lines. Cell cycle.
3:414–415. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Feuring-Buske M and Hogge DE: Hoechst
33342 efflux identifies a subpopulation of cytogenetically normal
CD34(+)CD38(−) progenitor cells from patients with acute myeloid
leukemia. Blood. 97:3882–3889. 2001.PubMed/NCBI
|
|
7
|
Uchida N, Fujisaki T, Eaves AC and Eaves
CJ: Transplantable hematopoietic stem cells in human fetal liver
have a CD34(+) side population (SP)phenotype. J Clini Invest.
108:1071–1077. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ho MM, Ng AV, Lam S and Hung JY: Side
population in human lung cancer cell lines and tumors is enriched
with stem-like cancer cells. Cancer Res. 67:4827–4833. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chiba T, Kita K, Zheng YW, Yokosuka O,
Saisho H, Iwama A, Nakauchi H and Taniguchi H: Side population
purified from hepatocellular carcinoma cells harbors cancer stem
cell-like properties. Hepatology. 44:240–251. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zen Y, Fujii T, Yoshikawa S, Takamura H,
Tani T, Ohta T and Nakanuma Y: Histological and culture studies
with respect to ABCG2 expression support the existence of a cancer
cell hierarchy in human hepatocellular carcinoma. Am J Pathol.
170:1750–1762. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hu C, Li H, Li J, Zhu Z, Yin S, Hao X, Yao
M, Zheng S and Gu J: Analysis of ABCG2 expression and side
population identifies intrinsic drug efflux in the HCC cell line
MHCC-97L and its modulation by Akt signaling. Carcinogenesis.
29:2289–2297. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yan F, Wang XM, Liu ZC, Pan C, Yuan SB and
Ma QM: JNK1, JNK2, and JNK3 are involved in P-glycoprotein-mediated
multidrug resistance of hepatocellular carcinoma cells.
Hepatobiliary Pancreat Dis Int. 9:287–295. 2010.PubMed/NCBI
|
|
13
|
Sancho R, Nateri AS, de Vinuesa AG,
Aguilera C, Nye E, Spencer-Dene B and Behrens A: JNK signalling
modulates intestinal homeostasis and tumourigenesis in mice. EMBO
J. 28:1843–1854. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lu S, Shen KC, Wang Y, Brooks SC and Wang
YA: Impaired hepatocyte survival and liver regeneration in
Atm-deficient mice. Hum Mol Genet. 14:3019–3025. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Komoda F, Shino Y, Hirano T, Okutomi Y,
Okamoto H, Hayashi Y, Suyama T, Ebara M, Saisho H and Shirasawa H:
MEKK1 induces c-Jun complexes that act as negative regulators for
cell survival and proliferation of HCC cells. Int J Oncol.
21:553–559. 2002.PubMed/NCBI
|
|
16
|
Goodell MA: Multipotential stem cells and
‘side population’ cells. Cytotherapy. 4:507–508. 2002.
|
|
17
|
Bonnet D and Dick JE: Human acute myeloid
leukemia is organized as a hierarchy that originates from a
primitive hematopoietic cell. Nat Med. 3:730–737. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Sharma SV, Lee DY, Li B, Quinlan MP,
Takahashi F, Maheswaran S, McDermott U, Azizian N, Zou L, Fischbach
MA, Wong KK, Brandstetter K, Wittner B, Ramaswamy S, Classon M and
Settleman J: A chromatin-mediated reversible drug-tolerant state in
cancer cell subpopulations. Cell. 141:69–80. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yoon CH, Kim MJ, Kim RK, Lim EJ, Choi KS,
An S, Hwang SG, Kang SG, Suh Y, Park MJ and Lee SJ: c-Jun
N-terminal kinase has a pivotal role in the maintenance of
self-renewal and tumorigenicity in glioma stem-like cells.
Oncogene. 31:4655–4666. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Nagata H, Hatano E, Tada M, Murata M,
Kitamura K, Asechi H, Narita M, Yanagida A, Tamaki N, Yagi S, Ikai
I, Matsuzaki K and Uemoto S: Inhibition of c-Jun NH2-terminal
kinase switches Smad3 signaling from oncogenesis to tumor-
suppression in rat hepatocellular carcinoma. Hepatology.
49:1944–1953. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Mucha SR, Rizzani A, Gerbes AL, Camaj P,
Thasler WE, Bruns CJ, Eichhorst ST, Gallmeier E, Kolligs FT, Goke B
and De Toni EN: JNK inhibition sensitises hepatocellular carcinoma
cells but not normal hepatocytes to the TNF-related
apoptosis-inducing ligand. Gut. 58:688–698. 2009. View Article : Google Scholar
|
|
22
|
Seki E, Brenner DA and Karin M: A liver
full of JNK: signaling in regulation of cell function and disease
pathogenesis, and clinical approaches. Gastroenterology.
143:307–320. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhang DM, Shu C, Chen JJ, Sodani K, Wang
J, Bhatnagar J, Lan P, Ruan ZX, Xiao ZJ, Ambudkar SV, Chen WM, Chen
ZS and Ye WC: BBA, a derivative of 23-hydroxybetulinic acid,
potently reverses ABCB1-mediated drug resistance in vitro and in
vivo. Mol Pharm. 9:3147–3159. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hagiwara S, Kudo M, Chung H, Ueshima K,
Inoue T, Haji S, Watanabe T, Park AM, Munakata H and Sakurai T:
2012. Activation of c-Jun N-terminal kinase in non-cancerous liver
tissue predicts a high risk of recurrence after hepatic resection
for hepatocellular carcinoma. Hepatol Res. 42:394–400. 2012.
View Article : Google Scholar
|