Introduction
Ischemic stroke is one of the leading causes of
mortality (1) and long-term
disability in adults worldwide (2). Three months following a stroke,
~15–30% of stroke survivors are permanently disabled and 20%
require costly long-term care (3).
Deficits include partial paralysis, and difficulties with memory,
thinking, language and movement. According to the current data,
~80% of strokes are ischemic (4).
Ischemic strokes result from a transient or permanent reduction in
cerebral blood flow that is restricted to the territory of a major
brain artery (5). The reduction in
flow is, in the majority of cases, caused by middle cerebral artery
occlusion (MCAO) either by an embolus or local thrombosis. In the
center of the ischemic territory, oxygen and glucose deprivation,
neuronal depolarization and Ca2+-mediated excitotoxicity
induces necrotic and apoptotic cell death (6). The amount of excitotoxicity and
oxidative damage in cerebral tissue depends on several factors,
including the degree and the duration of ischemia, and the
capability of the brain to recover and repair itself (3).
Rigorous laboratory investigations of cerebral
ischemia conducted over the past two decades have identified
various factors that are involved in the pathogenesis of ischemic
stroke, including inflammation, excitotoxicity and ionic imbalance,
oxidative and nitrosative stress, as well as apoptotic-like cell
death (7). In particular,
increasing evidence demonstrates that serological markers of
inflammation, including C-reactive protein and soluble
intercellular adhesion molecule account for the pathogenic
progression of ischemic stroke (8). Despite advances in the understanding
of the pathophysiology of ischemic stroke, the precise molecular
mechanisms involved in ischemic stroke induced by MCAO remain
poorly understood.
Therefore, in the present study, microarrays were
utilized to identify the differentially expressed genes (DEGs)
between sham samples and MCAO-induced focal ischemic samples at
various time-points (1, 3 and 7 days). Gene Ontology (GO)enrichment
analysis was performed and a protein-protein interaction (PPI)
network was constructed by mapping the DEGs to the PPI data. This
information may facilitate the understanding of the molecular
mechanisms underlying ischemic stroke and thus aid in selecting an
appropriate and effective treatment strategy for patients.
Materials and methods
Affymetrix microarray data
The transcriptional profile of GSE35338 (9) was obtained from National Center of
Biotechnology Information Gene Expression Omnibus (GEO) database
(http://www.ncbi.nlm.nih.gov/geo/), which
is based on the Affymetrix Mouse Genome 430 2.0 Array (Affymetrix,
Inc., Santa Clara, CA, USA). In total, 21 specimens, obtained one
day (n=5), three days (n=3) and seven days (n=3) following
MCAO-induced ischemic stroke, and one day (n=4), three days (n=3)
and seven days (n=3) following control sham surgery, were available
based on the GPL1261 Platform.
Data preprocessing
The probe-level data in CEL files (Affymetrix Inc.)
were converted into expression measures and background correction
was performed by the robust multiarray average algorithm (10) with defaulted parameters in the R
affy package (11,12). If there were multiple probe sets
that corresponded to the same gene, the expression values of those
probe sets were averaged.
DEG analysis
For the GSE 35338 dataset, LIMMA package (13) in R language (Affymetrix Inc.) was
used to identify DEGs between the MCAO and sham control samples.
Only the DEGs with a fold change value >1.5 and a P-value
<0.05 were selected.
GO and Kyoto Encyclopedia of Genes and
Genomes (KEGG) pathway analysis
GO analysis has become a commonly utilized approach
for functional annotation of large-scale genomic data (14).
The KEGG pathway database (15) (http://www.genome.jp/kegg/pathway.html) contains
information of the manner in which molecules or genes are
networked. It is complementary to the majority of the existing
molecular biology databases that contain information of individual
molecules or individual genes.
The database for annotation, visualization and
integrated discovery (DAVID Bioinformatics Resources 6.7;
http://david.abcc.ncifcrf.gov/home.jsp), a
high-throughput and integrated data-mining environment, analyzes
gene lists derived from high-throughput genomic experiments
(16). In the present study, DAVID
was used to identify over-represented GO categories in biological
processes and significant pathways with a value of P<0.05.
PPI network construction
To demonstrate the potential PPI correlation, the
DEGs were mapped to the PPI data that were collected from the
Search Tool for the Retrieval of Interacting Genes (STRING)
(17) database. STRING is a large
dataset containing functional links between proteins on the basis
of experimental evidence for PPIs as well as interactions predicted
by comparative genomics and text mining. It uses a scoring system
that is intended to reflect the evidence of predicted interactions.
In the present study, interactions with a score ≥0.7 were included.
Next, a PPI network was constructed by Cytoscape (18) based on the PPI correlations.
Molecular Complex Detection (MCODE)
analysis
MCODE (ftp://ftp.mshri.on.ca/pub/BIND/Tools/MCODE) detects
densely connected regions in large PPI networks that may represent
molecular complexes (19). In the
present study, clusters of highly intra-connected nodes (n>10)
in the network were searched using an MCODE plug-in in the
Cytoscape network. Next, the identified clusters were used for
functional enrichment analysis.
Results
DEG selection
In order to obtain the DEGs between MCAO reactive
astrocytes and sham controls at various time points, publically
available microarray datasets were obtained from the GEO. A total
of 294 genes were selected as DEGs between samples obtained from
one day following MCAO and sham specimens; 87 DEGs between samples
from three days following MCAO and sham samples; and 57 DEGs
between samples from seven days following MCAO and sham controls
with a fold-change >1.5 and P<0.05. The samples obtained from
one, three and seven days following MCAO had overlapping but
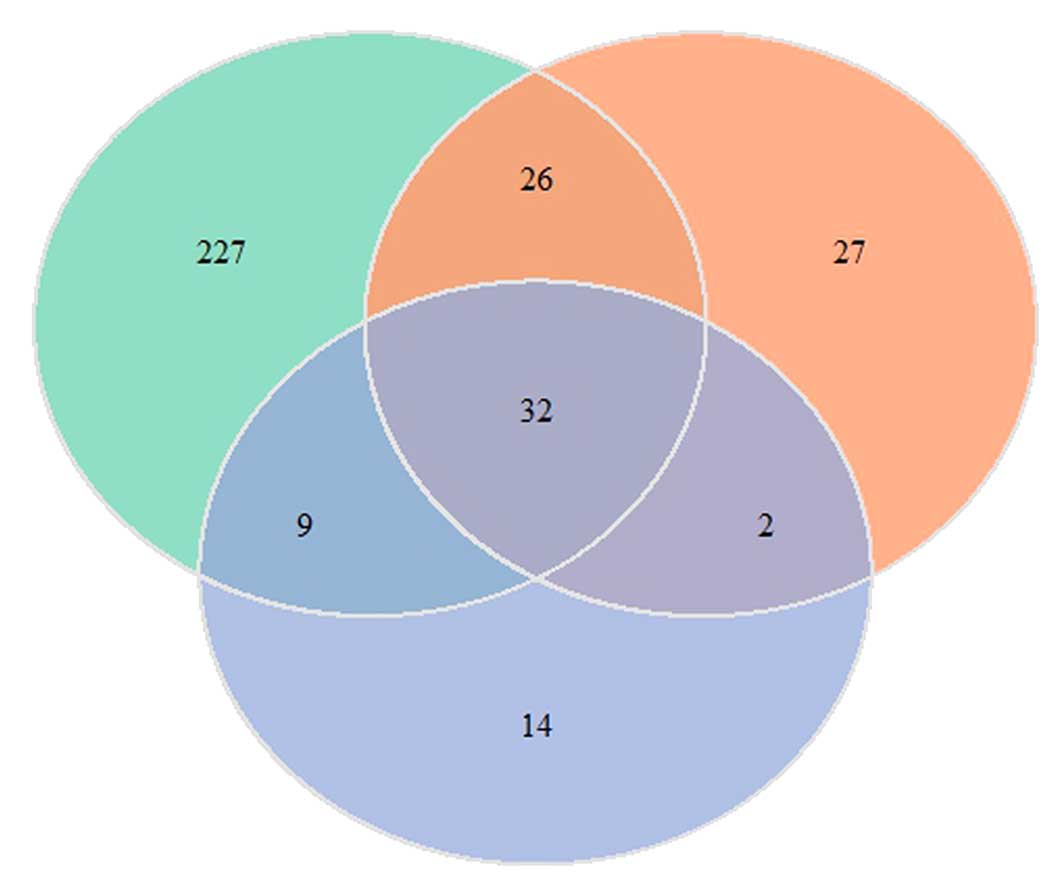
distinct sets of DEGs. The Venn diagram (Fig. 1) demonstrates that 32 genes are
common to the three MCAO samples and all of these genes were
upregulated in the MCAO-reactive astrocytes. There were 227, 27 and
14 distinct DEGs in the samples taken from one, three and seven
days following MCAO, respectively.
GO enrichment analysis of DEGs
To investigate the functional changes in the
pathological course of MCAO, the DEGs were mapped to the GO
database. This project provided three structured networks of
defined terms to describe the gene product attributes: Biological
process (BP), molecular function (MF) and cellular compartment
(CC). In the present study, the majority of the enriched genes were
upregulated in the MCAO samples, particularly in the samples from
seven days following MCAO. The DEGs for the samples taken from one
day following MCAO were most commonly associated with BP and CC,
including the extracellular region, response to wounding and immune
response (Table I). Similarly, the
DEGs in samples taken from three days following MCAO were also
mainly associated with BP and CC, for instance, extracellular
region, cell cycle, response to wounding and defense response
(Table II). Table III demonstrates that the enriched
GO terms of DEGs in the samples taken from seven days following
MCAO were correlated with all of the three defined terms. The
enriched BP GO terms included immune response, response to wounding
and inflammatory response. The enriched CC GO terms included
extracellular region and extracellular space. The enriched MF GO
terms included Ca2+ ion binding and enzyme inhibitor
activity. In addition, significantly enriched GO terms with high
counts of distinct DEGs in each MCAO sample were identified. The
enriched terms of the distinct DEGs in samples obtained from one
day following MCAO included cell death, oxidation reduction and
response to wounding (Table V).
The enriched terms of the distinct DEGs in samples obtained from
three days following MCAO included cell cycle, cell division and
nuclear division. The enriched term of the specific DEGs in samples
obtained from seven days following MCAO was cell-cell
signaling.
 | Table ITop ten significantly enriched GO
terms with a high count of DEGs in the samples one day following
MCAO. |
Table I
Top ten significantly enriched GO
terms with a high count of DEGs in the samples one day following
MCAO.
| Term | Category | Description | Count | P-value |
|---|
| GO: 0005886 | CC | Plasma
membrane | 50 | 2.34E-02 |
| GO: 0005576 | CC | Extracellular
region | 49 | 4.91E-08 |
| GO: 0044421 | CC | Extracellular
region part | 35 | 2.22E-10 |
| GO: 0009611 | BP | Response to
wounding | 27 | 6.07E-13 |
| GO: 0006955 | BP | Immune
response | 23 | 2.53E-07 |
| GO: 0005615 | CC | Extracellular
space | 23 | 7.05E-07 |
| GO: 0042127 | BP | Regulation of cell
proliferation | 22 | 8.30E-06 |
| GO: 0006952 | BP | Defense
response | 21 | 1.86E-06 |
| GO: 0008219 | BP | Cell death | 20 | 4.01E-05 |
| GO: 0016265 | BP | Death | 20 | 5.50E-05 |
| Table IITop ten significantly enriched GO
terms with a high count of DEGs in the samples three days following
MCAO. |
Table II
Top ten significantly enriched GO
terms with a high count of DEGs in the samples three days following
MCAO.
| Term | Category | Description | Count | P-value |
|---|
| GO: 0005576 | CC | Extracellular
region | 18 | 2.24E-04 |
| GO: 0044421 | CC | Extracellular
region part | 13 | 4.90E-05 |
| GO: 0007049 | BP | Cell cycle | 12 | 1.36E-04 |
| GO: 0042127 | BP | Regulation of cell
proliferation | 11 | 2.19E-04 |
| GO: 0009611 | BP | Response to
wounding | 10 | 3.77E-05 |
| GO: 0051301 | BP | Cell division | 9 | 5.50E-05 |
| GO: 0006955 | BP | Immune
response | 9 | 1.74E-03 |
| GO: 0005615 | CC | Extracellular
space | 9 | 9.95E-04 |
| GO: 0000278 | BP | Mitotic cell
cycle | 8 | 1.58E-04 |
| GO: 0022403 | BP | Cell cycle
phase | 8 | 9.39E-04 |
| Table IIITop ten significantly enriched GO
terms with a high count of DEGs in the samples seven days following
MCAO. |
Table III
Top ten significantly enriched GO
terms with a high count of DEGs in the samples seven days following
MCAO.
| Term | Category | Description | Count | P-value |
|---|
| GO: 0005576 | CC | Extracellular
region | 21 | 3.08E-08 |
| GO: 0044421 | CC | Extracellular
region part | 16 | 6.15E-09 |
| GO: 0005615 | CC | Extracellular
space | 13 | 3.75E-08 |
| GO: 0006955 | BP | Immune
response | 11 | 1.56E-06 |
| GO: 0009611 | BP | Response to
wounding | 9 | 1.11E-05 |
| GO: 0006952 | BP | Defense
response | 9 | 6.89E-05 |
| GO: 0006954 | BP | Inflammatory
response | 8 | 6.03E-06 |
| GO: 0005509 | MF | Calcium ion
binding | 8 | 1.34E-02 |
| GO: 0042127 | BP | Regulation of cell
proliferation | 7 | 6.57E-03 |
| GO: 0004857 | MF | Enzyme inhibitor
activity | 6 | 8.59E-04 |
| Table VSignificantly enriched GO terms with
a high count of distinct DEGs in MCAO samples. |
Table V
Significantly enriched GO terms with
a high count of distinct DEGs in MCAO samples.
| Term | Description | Count | P-value |
|---|
| One day after
MCAO |
| GO: 0008219 | Cell death | 16 | 1.32E-04 |
| GO: 0016265 | Death | 16 | 5.22E-04 |
| GO: 0055114 | Oxidation
reduction | 16 | 6.65E-04 |
| GO: 0012501 | Programmed cell
death | 15 | 8.01E-04 |
| GO: 0009611 | Response to
wounding | 14 | 1.55E-03 |
| GO: 0006915 | Apoptosis | 14 | 2.05E-03 |
| GO: 0042127 | Regulation of cell
proliferation | 13 | 2.82E-03 |
| GO: 0042981 | Regulation of
apoptosis | 12 | 3.28E-03 |
| GO: 0043067 | Regulation of
programmed cell death | 12 | 3.67E-03 |
| GO: 0010941 | Regulation of cell
death | 12 | 7.83E-03 |
| Three days after
MCAO |
| GO: 0007049 | Cell cycle | 10 | 7.63E-08 |
| GO: 0051301 | Cell division | 8 | 9.24E-08 |
| GO: 0000280 | Nuclear
division | 7 | 2.27E-07 |
| GO: 0007067 | Mitosis | 7 | 2.27E-07 |
| GO: 0000087 | M phase of mitotic
cell cycle | 7 | 2.57E-07 |
| GO: :0048285 | Organelle
fission | 7 | 2.82E-07 |
| GO: 0000278 | Mitotic cell
cycle | 7 | 9.90E-07 |
| GO: 0000279 | M phase | 7 | 2.35E-06 |
| GO: 0022403 | Cell cycle
phase | 7 | 5.50E-06 |
| GO: 0022402 | Cell cycle
process | 7 | 1.55E-05 |
| Seven days after
MCAO |
| GO: 0007267 | Cell-cell
signaling | 3 | 0.046035188 |
Pathway enrichment analysis
To gain further insights into the changes in the
biological pathways in the cells in the MCAO samples, the online
biological classification tool DAVID was used and significant
enrichment of these DEGs in multiple KEGG terms was observed
(Table IV). The most
significantly enriched pathway that the DEGs in samples from one
day following MCAO were involved in was cytokine-cytokine receptor
interaction. The most significantly enriched pathway that the DEGs
in samples from three days following MCAO were involved in was the
p53 signaling pathway. The DEGs in this group were also shown to be
involved in the cell cycle, cytokine-cytokine receptor interaction
and cytosolic DNA-sensing pathway. In the samples from seven days
following MCAO, the pathways correlated with the DEGs were
cytokine-cytokine receptor interaction, the nucleotide-binding
oligomerization domain (NOD)-like receptor signaling pathway and
the chemokine signaling pathway.
| Table IVEnriched KEGG pathway of DEGs in MCAO
samples one day following MCAO. |
Table IV
Enriched KEGG pathway of DEGs in MCAO
samples one day following MCAO.
| Pathway | Genes | P-value |
|---|
| One day following
MCAO |
| Cytokine-cytokine
receptor interaction | CXCL1, IL-6, CCL2,
CCL7, OSMR, MET, CXCL2, EDA2R, CXCL10, IL11, CLCF1, FAS, IL13RA1,
TNFRSF12A | 5.87E-05 |
| p53 signaling
pathway | CDKN1A, CCND1,
SERPINE1, CASP8, RPRM, CDK6, FAS, GADD45B, IGFBP3, GADD45A | 6.80E-07 |
| MAPK signaling
pathway | MAP3K6, PLA2G4A,
BDNF, HSPB1, FAS, GADD45B, FLNC, GADD45A, CD14, FLNA | 1.706E-02 |
| Focal
adhesion | CCND1, CAV1, MET,
COL6A2, COL6A1, FLNC, FLNA, SPP1 | 2.83E-02 |
| Complement and
coagulation cascades | A2M, THBD,
SERPINE1, SERPING1, C1S, BDKRB2, PROS1 | 8.37E-04 |
| Jak-STAT signaling
pathway | CCND1, IL-6, CLCF1,
OSMR, SOCS3, IL13RA1, IL11 | 2.59E-02 |
| NOD-like receptor
signaling pathway | CXCL1, IL-6, CCL2,
CXCL2, CASP8, CCL7 | 2.23E-03 |
| Toll-like receptor
signaling pathway | IL-6, MYD88, CASP8,
CD14, CXCL10, SPP1 | 1.60E-02 |
| Hematopoietic cell
lineage | IL-6, CD44, CD24A,
CD14, IL-11 | 3.67E-02 |
| Melanoma | CDKN1A, CCND1, MET,
CDK6 | 8.99E 02 |
| Three days
following MCAO |
| p53 signaling
pathway | CDK1, CCND1, CCNB2,
RRM2, SERPINE1, IGFBP3 | 2.45E-05 |
| Cell cycle | CDK1, CCND1, CCNB2,
CDKN2B, CDC20 | 4.13E-03 |
| Cytokine-cytokine
receptor interaction | IL-6, CCL2, CLCF1,
TNFRSF12A, CXCL10 | 3.67E-02 |
| Cytosolic
DNA-sensing pathway | IFI202B, IL6,
CXCL10 | 3.31E-02 |
| Seven days
following MCAO |
| Cytokine-cytokine
receptor interaction | CXCL1, CCL2, CXCL2,
TNFSF9, CXCL10 | 7.46E-03 |
| NOD-like receptor
signaling pathway | CXCL1, CCL2, CXCL2,
TNFAIP3 | 1.03E-03 |
| Chemokine
signaling pathway | CXCL1, CCL2, CXCL2,
CXCL10 | 2.09E-02 |
PPI network construction
To construct the PPI network, PPI data was obtained
from the STRING database. In the network, each edge is examined by
a score as the edge weight to quantify the interaction confidence.
To obtain the correlations, the PPIs with a score of ≥0.7 were
selected (Fig. 2). Next, the
degree of each node in the networks was calculated by iGrph, a
publicly available R package for analyzing graphs. The degree is
the number of edges connecting all of the nodes in the network. A
higher value for the degree indicates a highly connected network
and is likely to be more robust. A total of 22 nodes were screened
with degrees >10. Notably, the degrees of CXC motif chemokine 10
(CXCL10) and interleukin-6 (IL-6) were >20, suggesting they may
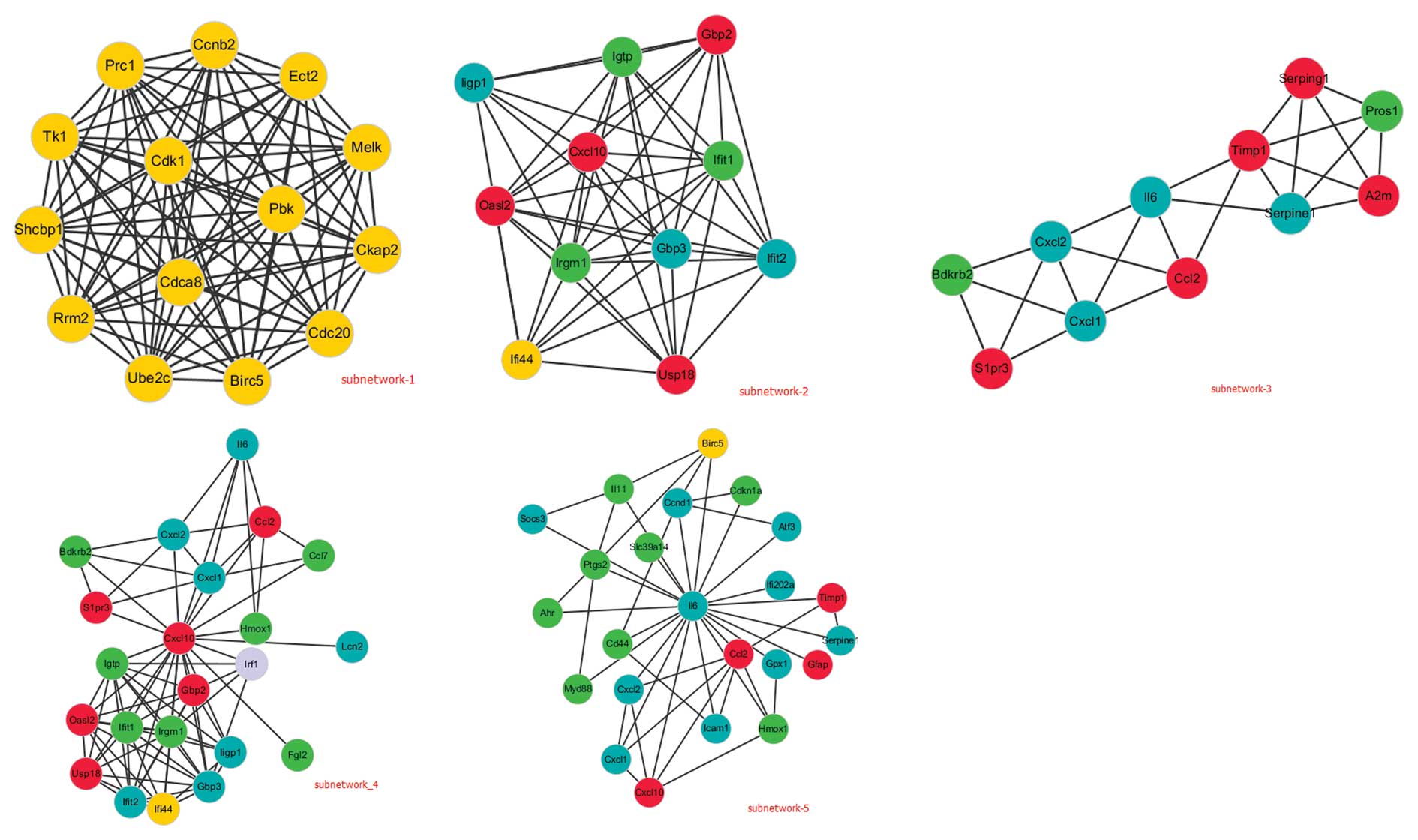
have an important role in MCAO-induced ischemia. In Fig. 3, these two DEGs as well as their
first nodes formed local networks (sub-network 4 and sub-network
5). In addition, the network was further analyzed by MCODE and
three sub-networks (sub-network 1–3) were searched with the
intra-connected nodes >10. The functions of these sub-networks
were mainly correlated with the cell cycle, immune response,
response to wounding and regulation of cell proliferation.
Discussion
Stroke is one of the most common causes of mortality
and disability, with marked financial repercussions on health
systems worldwide (20). Altered
gene expression is an important feature of ischemic cerebral injury
and affects proteins in numerous functional classes (21). Therefore, an understanding of the
molecular mechanisms underlying this disease is critically
important for developing effective management strategies. In the
present study, a bioinformatics method was utilized to examine the
molecular mechanism of MCAO-induced ischemic stroke development at
various time points. A total of 337 DEGs were identified between
the MCAO and sham control samples. These genes included 227
distinct DEGs in the samples obtained from one day following MCAO,
27 distinct DEGs in the samples from three days following MCAO and
14 distinct DEGs in the samples from seven days following MCAO. The
cytokine-cytokine receptor interaction pathway, p53 signaling
pathway and mitogen-activated protein kinase (MAPK) signaling
pathway were dysregulated in the MCAO samples. By mapping DEGs to a
PPT database, a PPT network was constructed, which revealed the
interaction of DEGs. Through this network, it was identified that
the node magnitude of CXCL10 and IL-6 were larger with degrees of
>20.
CXCL10, a chemokine that targets activated T cells
and natural killer cells expressing CXCR3, has been implicated in
inflammatory disease and is most commonly associated with T cell
responses (22–25). CXCL10 is expressed by neurons in
response to brain injury and leads to the recruitment of microglia
for the purpose of dendritic reorganization (26). Exogenous application of CXCL10 has
been demonstrated to induce neuronal apoptosis and to inhibit
herpes simplex virus replication in neurons in vitro
(27). The CXCL10 chemokines
appear to be essential for immune cell activation and trafficking
of peripheral immune cells across the blood-brain barrier (28,29).
Previously, CXCL10 has also been reported to have an important role
in ischemia/reperfusion-induced liver inflammation and
hepatocellular injury (25). In
the present study, CXCL10 acted as a hub node in the network
suggesting this gene has an important role in ischemic stroke
development and may be used as a specific therapeutic molecular
target in the treatment of ischemic stroke.
IL-6 is an acute phase reactant cytokine with pro-
and anti-inflammatory properties (30). IL-6 is produced by several cell
types, including fibroblasts, monocytes, adipocytes and endothelial
cells (31). IL-6 has been
demonstrated to be able to modulate cardiovascular function and
exert a negative inotropic effect via nitric oxide-dependent
pathways (32,33). An increasing number of experimental
observations suggest that IL-6 has a central role in the
pathogenesis of several ischemic cardiovascular disorders,
including unstable angina (34)
and acute coronary syndromes (35). Furthermore, IL-6 is also considered
to be associated with the initiation of liver regeneration in mice
(30). In humans, IL-6 is involved
in the acute phase response that follows cerebral ischemia, and
there is a correlation between high plasma levels of IL-6 and
occurrence of early neurological deterioration following stroke
(36) and progression of lacunar
infarction (37). In accordance
with the present findings, Flex et al (38) also suggested that IL-6 is
significantly and independently associated with a history of
ischemic stroke.
From the results of GO enrichment analysis, it was
identified that the majority of enriched GO terms of DEGs in the
samples obtained from one day following MCAO were correlated with
cell death and oxidant reduction. This suggested that cell death
and the lack of oxygen may have an important role in the onset of
MCAO-induced ischemic stroke. This finding is consistent with that
of a study by Mergenthaler et al (39), which suggested that programmed cell
death was initiated hours following ischemia onset and lasted over
a number of days. Oxidative stress contributes to the pathogenesis
of a number of neurological conditions, including stroke. Its
involvement in ischemic cell death results from the formation of
ROS/reactive nitrogen species through multiple injury mechanisms
(3). By three and seven days
following MCAO, the majority of the DEGs enriched in GO terms were
associated with the cell cycle and cell-cell signaling,
respectively. This indicated that cell proliferation and cell-cell
signaling may be essential in the pathogenesis of ischemic stroke
development. These results are consistent with a previous study by
Zamanian et al (9) who
reported that the expression of numerous genes associated with the
cell-cycle, including late-phase cyclin B and cyclin-dependent
kinase Cdk1, were not induced one day following MCAO but were
elevated 3-fold to 4-fold in MCAO reactive astrocytes three days
later. The results of GO enrichment analysis also indicated that
ischemic brain injury results from a complex sequence of
pathophysiological events that evolve over time.
The resulting PPI network is unweighted, since each
PPI occurred only once. As it is too large to yield more specific
information, it is necessary to divide the network into
sub-networks, which may represent functional modules or protein
sub-complexes. In the present study, clustering using MCODE and
first hub nodes identified five sub-networks. The main functions of
subnetwork-2 and subnetwork-4 were correlated with the immune
response. Lakhan et al (3)
reported that severe brain ischemia perturbed innate and adaptive
immune cells, resulting in systemic immunodepression that
predisposes stroke patients to life-threatening infections.
Manipulation of the immune system through mucosal tolerance may
provide a novel tool for stroke prophylaxis in humans (7). Notably, all of the DEGs enriched in
subnetwork-1 were only observed in the samples obtained from three
days following MCAO, whose GO terms were cell cycle and cell
division, suggesting that they may be involved in the processes of
the cell cycle.
In conclusion, the present study analyzed the gene
expression profiles and pathways that may be involved in the
progression of MCAO-induced ischemic stroke by using comprehensive
bioinformatics analysis. It was identified that CXCL10 and IL-6 may
have important roles in the progression of ischemic stroke and thus
may be used as specific therapeutic molecular targets. Furthermore,
ischemic brain injury resulted from a complex sequence of
pathophysiological events that evolved over time. Notably,
cell-cycle genes were only induced three days following MCAO.
However, further studies are required to confirm these observations
and determine their clinical utility in the therapeutic management
of ischemic stroke.
References
|
1
|
Yilmaz G and Granger DN: Cell adhesion
molecules and ischemic stroke. Neurol Res. 30:783–793. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Belayev L, Liu Y, et al: Human albumin
therapy of acute ischemic stroke: marked neuroprotective efficacy
at moderate doses and with a broad therapeutic window. Stroke.
32:553–560. 2001. View Article : Google Scholar
|
|
3
|
Lakhan SE, Kirchgessner A and Hofer M:
Inflammatory mechanisms in ischemic stroke: therapeutic approaches.
J Transl Med. 7:972009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Dichgans M: Genetics of ischaemic stroke.
Lancet Neurol. 6:149–161. 2007. View Article : Google Scholar
|
|
5
|
Dirnagl U, Iadecola C and Moskowitz MA:
Pathobiology of ischaemic stroke: an integrated view. Trends
Neurosci. 22:391–397. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Stoll G, Kleinschnitz C and Nieswandt B:
Molecular mechanisms of thrombus formation in ischemic stroke:
novel insights and targets for treatment. Blood. 112:3555–3562.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lo EH, Dalkara T and Moskowitz MA:
Mechanisms, challenges and opportunities in stroke. Nat Rev
Neurosci. 4:399–415. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Gorelick PB: Stroke prevention therapy
beyond antithrombotics: unifying mechanisms in ischemic stroke
pathogenesis and implications for therapy: an invited review.
Stroke. 33:862–875. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zamanian JL, Xu L, Foo LC, et al: Genomic
analysis of reactive astrogliosis. J Neurosci. 32:6391–6410. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Irizarry RA, Hobbs B, Collin F, et al:
Exploration, normalization, and summaries of high density
oligonucleotide array probe level data. Biostatistics. 4:249–264.
2003. View Article : Google Scholar
|
|
11
|
Gautier L, Cope L, Bolstad BM and Irizarry
RA: affy - analysis of Affymetrix GeneChip data at the probe level.
Bioinformatics. 20:307–315. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ihaka R and Gentleman R: R: A language for
data analysis and graphics. J Comput Graph Stat. 5:299–314.
1996.
|
|
13
|
Smyth GK: Linear models and empirical
bayes methods for assessing differential expression in microarray
experiments. Stat Appl Genet Mol Biol. 3:32004.PubMed/NCBI
|
|
14
|
Hulsegge I, Kommadath A and Smits MA:
Globaltest and GOEAST: two different approaches for Gene Ontology
analysis. BMC Proc. 3(Suppl 4): S102009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ogata H, Goto S, Sato K, et al: KEGG:
Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res.
27:29–34. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Huang da W, Sherman BT and Lempicki RA:
Systematic and integrative analysis of large gene lists using DAVID
bioinformatics resources. Nat Protoc. 4:44–57. 2009.PubMed/NCBI
|
|
17
|
Szklarczyk D, Franceschini A, Kuhn M, et
al: The STRING database in 2011: functional interaction networks of
proteins, globally integrated and scored. Nucleic Acids Res.
39:D561–D568. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Shannon P, Markiel A, Ozier O, et al:
Cytoscape: a software environment for integrated models of
biomolecular interaction networks. Genome Res. 13:2498–2504. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Bader GD and Hogue CW: An automated method
for finding molecular complexes in large protein interaction
networks. BMC Bioinformatics. 4:22003. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Allen C and Bayraktutan U: Oxidative
stress and its role in the pathogenesis of ischaemic stroke. Int J
Stroke. 4:461–470. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Mitsios N, Saka M, Krupinski J, et al: A
microarray study of gene and protein regulation in human and rat
brain following middle cerebral artery occlusion. BMC Neurosci.
8:932007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Belperio JA, Keane MP, Burdick MD, et al:
Critical role for CXCR3 chemokine biology in the pathogenesis of
bronchiolitis obliterans syndrome. J Immunol. 169:1037–1049. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hancock WW, Gao W, Csizmadia V, et al:
Donor-derived IP-10 initiates development of acute allograft
rejection. J Exp Med. 193:975–980. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Xie JH, Nomura N, Lu M, et al:
Antibody-mediated blockade of the CXCR3 chemokine receptor results
in diminished recruitment of T helper 1 cells into sites of
inflammation. J Leukocyte Biol. 73:771–780. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhai Y, Shen XD, Gao F, et al: CXCL10
regulates liver innate immune response against ischemia and
reperfusion injury. Hepatology. 47:207–214. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Rappert A, Bechmann I, Pivneva T, et al:
CXCR3-dependent microglial recruitment is essential for dendrite
loss after brain lesion. J Neurosci. 24:8500–8509. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Sui Y, Potula R, Dhillon N, et al:
Neuronal apoptosis is mediated by CXCL10 overexpression in simian
human immunodeficiency virus encephalitis. Am J Pathol.
164:1557–1566. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Dvoriantchikova G, Barakat D, Brambilla R,
et al: Inactivation of astroglial NF-kappa B promotes survival of
retinal neurons following ischemic injury. Eur J Neurosci.
30:175–185. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ubogu EE, Cossoy MB and Ransohoff RM: The
expression and function of chemokines involved in CNS inflammation.
Trends PharMCAOl Sci. 27:48–55. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Camargo CA Jr, Madden JF, Gao W, Selvan RS
and Clavien P: Interleukin-6 protects liver against warm
ischemia/reperfusion injury and promotes hepatocyte proliferation
in the rodent. Hepatology. 26:1513–1520. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yun S, Wu CY, Deng HM, et al: Synergistic
effect of -174 G/C polymorphism of the interleukin-6 gene promoter
and 469 E/K polymorphism of the intercellular adhesion molecule-1
gene in the populations of the Han nationality in Shenzhen with
cerebral infarction. Proceeding of Clinical Medicine. 24:584–587.
2009.
|
|
32
|
Finkel MS, Oddis CV, Jacob TD, et al:
Negative inotropic effects of cytokines on the heart mediated by
nitric oxide. Science. 257:387–389. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ono K, Matsumori A, Shioi T, Furukawa Y
and Sasayama S: Cytokine gene expression after myocardial
infarction in rat hearts possible implication in left ventricular
remodeling. Circulation. 98:149–156. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Hojo Y, Ikeda U, Takahashi M and Shimada
K: Increased levels of monocyte-related cytokines in patients with
unstable angina. Atherosclerosis. 161:403–408. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Plutzky J: Inflammatory pathways in
atherosclerosis and acute coronary syndromes. Am J Cardiol.
88:10K–15K. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Vila N, Castillo J, Dávalos A and Chamorro
A: Proinflammatory cytokines and early neurological worsening in
ischemic stroke. Stroke. 31:2325–2329. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Castellanos M, Castillo J, García MM, et
al: Inflammation-mediated damage in progressing lacunar infarctions
a potential therapeutic target. Stroke. 33:982–987. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Flex A, Gaetani E, Papaleo P, et al:
Proinflammatory genetic profiles in subjects with history of
ischemic stroke. Stroke. 35:2270–2275. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Mergenthaler P, Dirnagl U and Meisel A:
Pathophysiology of stroke: lessons from animal models. Metab Brain
Dis. 19:151–167. 2004. View Article : Google Scholar : PubMed/NCBI
|