Introduction
Lung cancer is the leading cause of mortality
worldwide, being the cause of >1,000,000 mortalities yearly
(1). In 2013, an estimated 82
males and 38 females per 100,000 will die from lung cancer
(2). Currently, surgery,
radiotherapy and chemotherapy serve as the most common treatments
in cancer therapy. In a large number of cases, patients get
diagnosed with cancer past the time-point of surgical treatment
being the most promising treatment method. Moreover, conventional
therapies, including radiotherapy and chemotherapy, frequently have
severe side effects as they do not selectively kill cancer cells
but also normal cells (3).
Therefore, it is important to discover novel treatments for lung
cancer.
Oncolytic therapy is a novel biological treatment
integrating gene and immune therapy, which is widely studied all
over the world. Oncolytic virotherapy is a promising form of gene
therapy for cancer, employing nature’s own agents to identify and
destroy malignant cells (4).
Newcastle disease virus (NDV) is one of these oncolytic viruses.
The first study on NDV being applied to treat uterine carcinoma
dates back to the early 1950s (5).
Since then NDV has been studied as an ideal candidate for cancer
therapy. NDV has numerous advantages making it an attractive viral
vector to cure cancer. For example, NDV has the ability to
selectively kill malignant cells, while not targeting normal cells.
It also has the ability to replicate in cancer cells, with a 10,000
fold higher efficiency opposed to that in normal cells (5).
NDV is a non-segmented negative-strand RNA virus,
which is part of the genus Avulavirus from the
Paramyxoviridae family. According to their pathogenic properties,
NDVs can be divided into three major pathotypes: Lentogenic
strains, mesogenic strains and velogenic strains (6,7),
while the LaSota NDV strain (or wild-type NDV strain) is a
lentogenic strain, which causes clinically insignificant or
inapparent respiratory disease. The RNA of the NDV genome encodes
six major structural proteins: Nucleoprotein (NP), phosphoprotein
(P), matrix protein (M), fusion protein (F),
haemagglutinin-neuraminidase (HN) and large (L) RNA-dependent RNA
polymerase, with the six genes being in the order
3′-NP-P-M-F-HN-L-5′ (8). The
infectious ability of NDV depends on hemagglutinin-neuraminidase
(HN) and fusion (F) glycoproteins. The two glycoproteins are
embedded in the viral lipid membrane. The F glycoprotein precursor
(F0) is proteolytically cleaved to F1 and F2 (4). Finally, the penetration of NDV to
target cells is completed by the endocytosis process (9).
The rabies virus (RV) is an enveloped, bullet-shaped
virus, which belongs to the genus Lyssavirus of the family
Rhabdoviridae. The RV genome is composed of 3′-N-P-M-G-L-5′,
encoding nucleoprotein (N), phosphoprotein (P), matrix protein (M),
glycoprotein (G) and RNA polymerase (L) (10). The rabies virus glycoprotein (RVG)
has been associated with apoptosis of host cells. Préhaud et
al (11) showed that the
Evelyn Rokitnicki Abelseth (ERA) strain of RV triggers the
caspase-dependent apoptosis of human cells. It is the induction of
G protein expression instead of N protein expression that resulted
in apoptosis. RV was used as the first virus to be assessed for the
treatment of 30 patients with melanomatosis, eight of whom showed
tumor regression in experimental settings, and in further clinical
trials during the 1950s and 1960s (4).
The rL-RVG used in the present study was provided by
the Harbin Veterinary Research Institute (13). The RVG was inserted at the position
between the P and M genes. rL-RVG retained its titers of up to
109.8 50% egg infective doses (EID50)/ml of
allantoic fluid. rL-RVG has the ability to spread from cell to cell
and to alter its self-replication in cell culture (12). Therefore, it was hypothesized that
the rL-RVG had the ability to self-enhance its effect on cancer
cells. Since the mechanisms of rL-RVG to kill cancer cells are not
well understood, the objective of the present study was to explore
the oncolytic effect of rL-RVG on A549 cells in vitro. The
findings indicated that rL-RVG may be used as a novel
biotherapeutic drug for lung cancer treatment.
Materials and methods
The recombinant NDV strain, the wild-type NDV LaSota
strain and the anti-NDV antibody were kindly provided by the Harbin
Veterinary Research Institute (Harbin, China), which were stored at
−70°C. All viruses were grown and titrated in A549 cells in 96-well
plates. Infection of viruses in A549 cells was confirmed by using
an indirect immunofluorescence assay: NDV infection of A549 cells
were grown in 96-well plates, cells were fixed in ice-cold 4%
paraformaldehyde in phosphatebuffered saline (PBS) for 15 min at
room temperature and subsequently washed with PBS, three times.
Cells were blocked in PBS containing 1% (wt/vol) bovine
serumalbumin (BSA) at 4°C for 1 h. Cells were then incubated with
chicken antibody against NDV for 30 min at room temperature, washed
three times with PBS containing 0.05% Tween 20 and stained with a
Cy3-conjugated goat anti-chicken antibody for 30 min. Following
this, the cells were washed three times with PBS, and their nuclei
stained with Hoechst 33342. Cells were subsequently analyzed under
an immunofluorescence microscope (Axio Observer A1; Carl Zeiss,
Oberkochen, Germany). Viral titration results were calculated by
using the method of Reed and Muench (12). The lung adenocarcinoma cell line
A549 was purchased from the Cell Culture Center of the Basic
Institute of Medical Sciences, Peking Union Medical College
(Peking, China). A549 cells were cultured in 24 well-plates with
Dulbecco’s modified Eagle’ medium (DMEM; Gibco Life Technologies.
Carlsbad, CA, USA)) containing 10% fetal bovine serum culture
medium, at 37°C and 5% CO2 incubator for cultivation.
The cell culture reagents were obtained from Gibco-BRL (Invitrogen
Life Technologies, Carlsbad, CA, USA). The 96-well plates, 24-well
plates and six-well plates were purchased from Costar Corning
(Corning, NY, USA). All polymerase chain reaction (PCR) primers
were synthesized by the Shanghai Sangon Biological Engineering
Technology & Services Co. Ltd (Shanghai, China). The TRIzol
reagent was purchased from Invitrogen Life Technologies. The PCR
master mix was obtained from Toyobo (Osaka, Japan). The terminal
deoxynucleotidyl transferase-mediated dUTP-biotin nick end-labeling
(TUNEL) assay kit was obtained from Nanjing Kaiji Biotechnology
Development Co. Ltd., (Nanjing, China). The antibody to chicken was
purchased from MedImmune, LLC (Gaithersburg, MD, USA). The mouse
anti-RV monoclonal antibody and caspase inhibitor
Z-Val-Ala-Asp-fluoromethylketone (Z-VAD-FMK) were purchased from
Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA). The rabbit
anti-Caspase-3, 8, 9 monoclonal antibodies, rabbit B-cell
lymphoma-2 (bcl-2)-associated X (bax) and mouse anti-bcl-2
monoclonal antibodies were purchased from Boster (Wuhan, China).
The horseradish peroxidase (HRP)-conjugated goat anti-rabbit,
HRP-conjugated goat anti-mouse antibody, Cy3-conjugated goat
anti-rabbit and the fluorescein isothiocyanate (FITC)-conjugated
goat anti-mouse antibody were purchased from CoWin Biotech, Co.,
Ltd. (Beijing, China). The HRP-AffiniPure rabbit anti-chicken was
purchased from EarthOx Life Science (Millbrae, CA, USA).
Reverse transcription PCR (RT-PCR)
analysis
Total RNA was extracted from lung adenocarcinoma
A549 cells by using TRIzol. The cDNA was synthesized with Oligo
(dT) primers (Takara Bio, Inc., Otsu, Japan). and Moloney murine
leukemia virus reverse transcriptase (Takara Bio, Inc.). The 1/10
cDNA was used for PCR amplification. The PCR protocol was as
follows: Initial denaturation at 94°C for 5 min, followed by 30
cycles at 94°C for 30 sec, annealing at 55°C (NDV) or 58°C for 30
sec, and extension at 72°C for 30 sec (RVG). The final extension
was performed by an incubation step at 72°C for 10 min. The PCR
products were subjected to electrophoresis in agarose gel
(Sigma-Aldrich, St. Louis, MO USA) and visualized with ethidium
bromide (Sigma-Aldrich). The bands were analyzed with Quantity One
software, version 4.62 (Bio-Rad, Hercules, CA, USA). All primers
used are listed in Table I.
 | Table IPrimers for polymerase chain reaction
amplification. |
Table I
Primers for polymerase chain reaction
amplification.
| Primer | Sequence | Product size
(bp) |
|---|
| rL-RVG |
| Upstream | 5′
AGCCGATGCTCACTACAAG 3′ | 175 |
| Downstream | 5′
CTGGAGGAGGGATGATTGC 3′ | |
| NDV |
| Upstream | 5′
CTGGACGGTTTGGTGGGAA 3′ | 462 |
| Downstream | 5′
TAATGCGACTGCGGGATGTG3′ | |
| GAPDH |
| Upstream | 5′
CAAGGTCATCCATGACAACTTTG3′ | |
| Downstream | 5′
GTCCACCACCCTGTTGCTGTAG3′ | |
Western blot analysis
Western blotting was performed to determine the
protein of NDV, RVG, bax, bcl-2 as well as caspase-3, -8 and -9
expression using the antibodies described above antibodies at a
dilution of 1:200. A549 cells were infected with rL-RVG and NDV at
a multiplicity of infection (MOI) of 10 (11). Whole cell lysates were prepared
with radioimmunoprecipitation assay buffer (KangChen Biotech Inc.,
Shanghai, China) containing protease inhibitor
(phenylmethanesulfonyl fluoride) and sodium orthovanadate, and 10
μl of each was loaded on a gel and the blot was run under reducing
conditions. The membrane was blocked for 1 h using 5% bovine serum
albumin (BSA) prior to incubation with chicken antibody against NDV
(1:300) and mouse antibody against RVG (1:200) overnight at 4°C.
Antibody binding was quantified using an anti-chicken
immunoglobulin (Ig)Y antibody or a goat anti-mouse IgG conjugated
to HRP at 1/10,000 dilution and visualized using an enhanced
chemiluminescence development solution, the Luminata TM Crescendo
Western HRP Substrate (Millipore, Middlesex, MA, USA).
Immunofluorescence analysis
Immunofluorescence analysis aimed to detect NDV and
RVG protein expression in infected A549 cells. A549 cells were
seeded in 24-well plates with 5×104 per well, and the
next day they were infected with NDV or rL-RVG at an MOI of 10
(11), while the negative group
was treated with phosphate-buffered saline (PBS). At 24 h
post-infection, cells were fixed in ice-cold 4% paraformaldehyde
(Solar Biotech Inc., Beijing, China) overnight at 4°C, then cells
were washed three times for 15 min each with ice-cold PBS. Cells
were blocked in 3% BSA at 4°C for 1 h. Following incubation with
chicken antibody against NDV (1:300) and mouse antibody against RVG
(1:200) for 1.5 h at room temperature, cells were then washed three
times with PBS. Next, cells were stained with Cy3-conjugated goat
anti-chicken (1:300) or FITC-conjugated goat anti-mouse secondary
antibody (1:100) for 30 min at room temperature, and were washed
three times with PBS. Finally, nuclei were stained with Hoechst
33342 (10 μg/ml; Sigma-Aldrich) for 10 min. Cells were analyzed
with a confocal laser microscope (Axio Observer A1; Zeiss,
Germany).
MTT assays
rL-RVG (109.8EID50/ml) and wild-type NDV strain
(109.8EID50/ml ) were diluted 103, 104, 105 and 106 times by
serum-free DMEM. A549 in the logarithmic growth phase were plated
at 1×104 cells/well in 96-well plates, incubated overnight,
infected by the diluted rL-RVG or NDV. DMEM containing 2% fetal
bovine serum (FBS) was added to the cells of the NDV and rL-RVG
groups on the following day. The PBS group was used as a negative
control. Then, 20 μl MTT (5 mg/ml) was added into each well. After
4 h of incubation, 150 μl DMSO solution was added to each well and
plates were agitated for 10 min. Finally, the absorbance was read
at 490 nm wavelength using standard spectrophotometer (FLX800;
Bio-tek instruments Inc., Shelburne, VT, USA). The above MTT
experiment was repeated three times and the tumor cell growth
inhibition rate was calculated. Simultaneously, an inverted phase
contrast microscope (ECLIPSE TS100; Nikon, Japan) was used to
observe the morphological changes of cells following infection with
rL-RVG and wild-type NDV following 24, 48 and 72 h.
Flow cytometric analysis
Flow cytometry was applied to examine early
apoptotic cells. A549 cells were infected with wild-type NDV and
rL-RVG in six-well plates containing 1×105 cells/well.
Infected cells were harvested 24, 48, 72 h later and washed twice
with ice-cold PBS. Cells were then resuspended in 1X binding buffer
from the BD Pharmingen FITC Annexin V Apoptosis Detection Kit I
(Becton Dickinson, Franklin Lakes, NJ, USA) at a concentration of
1×105 cells/ml. An aliquot of 100 μl of the solution was
transferred to a 5-ml culture tube. According to the manufacturer’s
instructions, 5 μl Annexin V-FITC and 5 μl propidium iodide (PI)
solution were added. Cells were then incubated for 20 min at room
temperature in the dark, followed by addition of 400 μl 1X binding
buffer to each tube. Infected cells were analyzed by flow cytometry
(Facscalibur, Becton Dickenson). All results were repeated three
times.
TUNEL assay
Cells in the logarithmic growth phase were infected
with the RVG and NDV and following 48 h cells were fixed.
Initially, the cover slips were in 24-well plates, and
subsequently, cells were grown on cover slips by micropipetter, add
rL-RVG and NDV, at 48 h postinfection, cells were fixed in 4%
formaldehyde for 20 min at room temperature (15–25°C). This was
followed by rinsing with PBS for 30 min and incubation with 3%
hydrogen peroxide in methanol for 10 min at room temperature. After
25-min washes with PBS, the samples were incubated with 0.1% Triton
X-100 and 0.1% sodium citrate in water for 30 min at room
temperature. For negative control, TdT was not added to the sample,
and for positive control, cells were treated by DNase I. After two
hours of washing with PBS, pretreated specimens were incubated with
50lL TdT labeling reaction buffer at 4°C overnight in dark and then
in a humidified at mosphere at 37°C for a further 2–3 hours.
Subsequently, the slides were incubated with 50lL streptavidin-HRP
for 60 min, followed by detection with 50lL diaminobenzidine
reagent for 10 min. Apoptosis was analyzed using the TUNEL assay
kit according to the manufacturer’s instructions. Cells were
observed and photographed under an optical microscope (ECLIPSE
TS100; Nikon, Japan). Areas were selected where the apoptotic cells
were distributed evenly, and the number of positive, cells out of
100 cells was counted. This was repeated three times The apoptotic
index (AI) was calculated by the number of apoptotic cells/(the
number of apoptotic cells + the number of non-apoptotic cells)
×100%.
Statistical analysis
Data were analyzed by SPSS 16.0 software
(International Business Machines, Armonk, NY, USA) and the results
are expressed as mean ± standard deviation. To compare differences
between multiple groups, statistical significance was analyzed
using Student’s t-test for data from the MTT assay, and one-way
analysis of variance for TUNEL and flow cytometry data. P<0.05
was considered to indicate a statistically significant difference
between values.
Results
Expression of viral genes and
proteins
RT-PCR was performed to assess RVG and NDV gene
expression in the A549 cells following infection with rL-RVG and
NDV. Gene-specific PCR products were detected, rather than full
genes. The analysis showed that the RVG gene (~175 bp) was
expressed in A549 cells following infection with rL-RVG. In the PBS
and NDV groups, the RVG gene was no expressed (Fig. 1Aa). Furthermore, the NDV HN gene
(~462 bp) was expressed following infection with rL-RVG and NDV. In
the PBS group, the HN gene was not expressed (Fig. 1Ab). The expression levels of RVG
and NDV protein were examined by western blotting and
immunofluorescence assays. The results showed that RVG protein was
expressed in A549 cells following infecttion with rL-RVG. However,
in the PBS and NDV groups, RVG protein was no expressed. At the
same time, NDV protein was detected in A549 cells following
infection with rL-RVG and NDV, and none of the proteins was
expressed in the PBS group (Fig. 1B
and C).
 | Figure 1Expression of the virus in A549 cells.
(A) a, Expression of RVG-mRNA; b, expression of HN-mRNA. Lanes: 1,
GAPDH in PBS group; 2, GAPDH in NDV group; 3, GAPDH in rL-RVG
group; M, maker; 4, rL-RVG group; 5, NDV group; 6, PBS group; 7,
blank. (B) Expression of RVG and NDV protein in A549 cells by
western blot analysis. Expression of RVG protein was present in the
rL-RVG group, and NDV protein was expressed in the rL-RVG and NDV
groups. The proteins were not expressed in the PBS group. (C)
Immunofluorescence analysis of RVG and NDV protein expression
(magnification, ×200). The RVG protein (green, top row) was only
expressed in the rL-RVG group, while the NDV protein (red, bottom
row) was expressed in the rL-RVG and NDV groups. Magnification,
×200. rL-RVG, LaSota NDV strain expressing rabies virus
glycoprotein; NDV, Newcastle disease virus; PBS, phosphate-buffered
saline. |
Effects of the virus rL-RVG on the
proliferation of cancer cells
It was shown that the viability of A549 cells
decreased with increasing virus concentration and incubation time
following infection. rL-RVG-infected and NDV-infected cells
exhibited morphological changes were observed, which were more
obvious in the rL-RVG group than in the NDV group. By contrast, no
changes were observed in the PBS group (Fig. 2A).
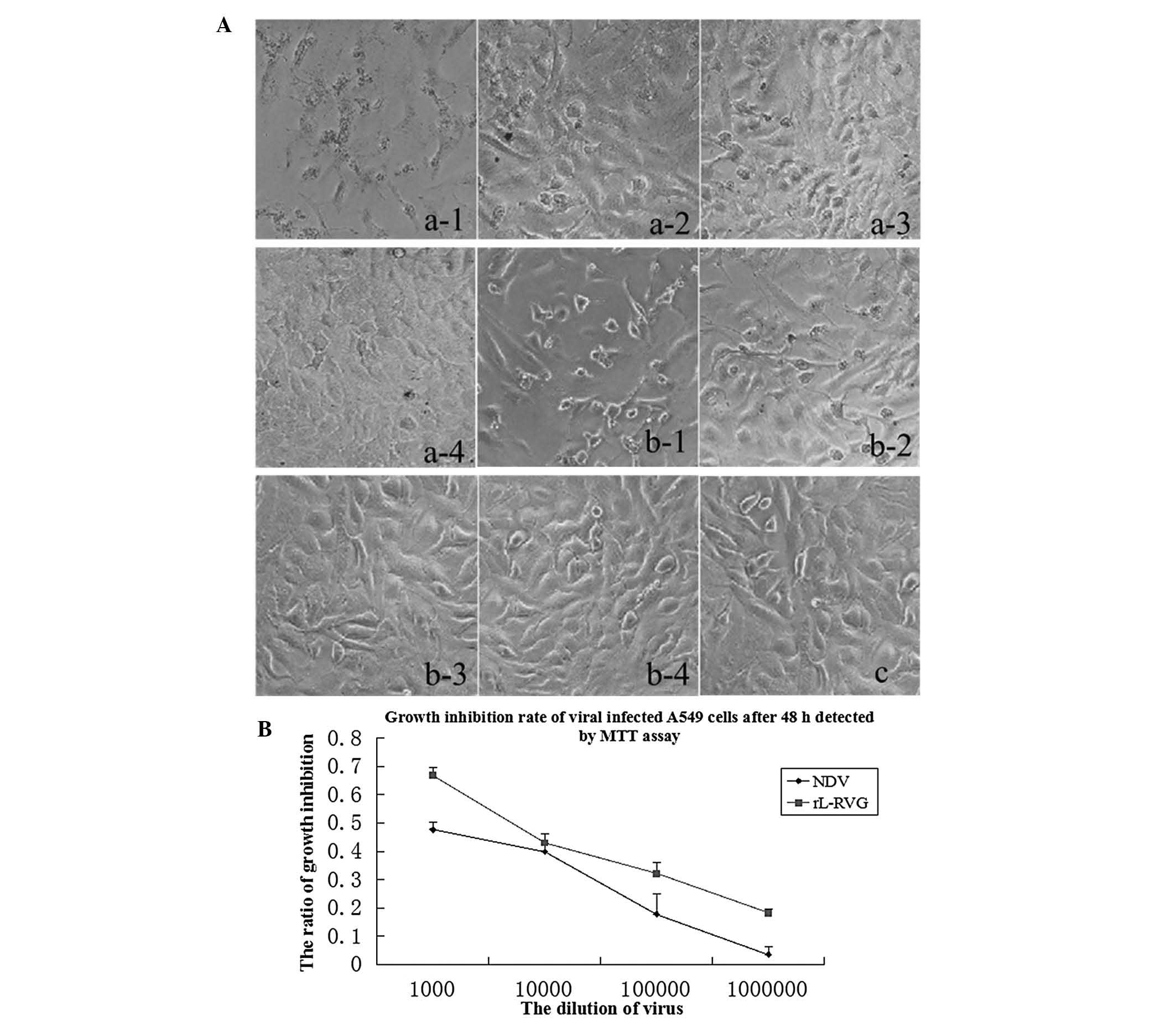 | Figure 2Morphological changes of A549 cells
following infection with virus. (A) Images of A549 cells infected
with virus on 48 h (magnification, 200×). a-1, 10−3
rL-RVG group; a-2, 10−4 rL-RVG group; a-3,
10−5 rL-RVG group; a4, 10−6 rL-RVG group;
b-1, 10−3 NDV group; b-2, 10−4 NDV group;
b-3, 10−5 NDV group; b-4, 10−6 NDV group; c,
phosphate-buffered saline group. The adherent ability of the cells
decreased. Magnification, ×200. (B) Growth inhibition rate of viral
infected A549 cells following 48 h determided by MTT assay. rL-RVG,
LaSota NDV strain expressing rabies virus glycoprotein; NDV,
Newcastle disease virus. |
The effect of different concentrations of
recombinant Newcastle disease virus rL-RVG and NDV on the growth of
A549 cells following 48 and 72 h was assessed by MTT assay. The
results showed that the inhibition rates in the rL-RVG group were
significantly higher than those infected with the wild-type NDV
strain, and the inhibition rate increased with time following
infection. The inhibition rate of A549 cells following 48 h is
shown in Fig. 2B and Table II.
| Table IIViral infection of A549 cells 48 h MTT
results of cell growth inhibition rate. |
Table II
Viral infection of A549 cells 48 h MTT
results of cell growth inhibition rate.
| Dilution | NDV, % | rL-RVG,
%a | t-value | P-value |
|---|
| 10−3 | 47.70±0.27 | 67.87±0.25 | 16.053 | 0.004 |
| 10−4 | 39.80±0.31 | 43.03±0.29 | 14.422 | 0.005 |
| 10−5 | 17.83±0.41 | 32.13±0.72 | 5.339 | 0.033 |
| 10−6 | 3.60±1.41 | 18.20±0.27 | 15.522 | 0.004 |
NDVs induce apoptosis in cancer
cells
Oncolytic NDV can induce apoptosis in a variety of
tumor cells. In the present study, the effect of RVG expression on
apoptosis induction by the recombinant virus was investigated using
Annexin V/PI double staining (Fig.
3A). The percentage of cells that underwent apoptosis was
higher in rL-RVG-infected than that in NDV-infected A549 cells at
48 h post-infection. Of note, the percentage of cells that
underwent apoptosis was significantly different between the rL-RVG
and NDV groups (P<0.05) (Fig.
3B). The induction of A549 cell apoptosis by rL-RVG and NDV was
dose- and time-dependent (data not shown). This suggested that
rL-RVG was more potent in inducing tumor cell apoptosis than its
parent virus, which indicated that RVG enhanced the oncolytic
properties of the parental virus.
 | Figure 3Apotopic effect induced by virus. (A)
Flow cytometric analysis of cells in early apoptosis following 48 h
of infection. a, rL-RVG 10−3 dilution group; b, NDV
10−3 dilution group; c, PBS group; d, rL-RVG
10−5 dilution group; e, NDV 10−5 dilution
group. (B) Quantification of early apoptotic cells from flow
cytometry experiment. a, 10−3 dilution; b,
10−5 dilution. One-way analysis of variance for
apoptotic cell percentages showed significant differences between
rL-RVG and NDV groups (P<0.05). (C) Terminal deoxynucleotidyl
transferase dUTP nick end labeling analysis of apoptosis in A549
cells following different infections, with arrows indicating
apoptotic cells. Magnification, ×200. rL-RVG, LaSota NDV strain
expressing rabies virus glycoprotein; NDV, Newcastle disease virus;
PBS, phosphate-buffered saline. |
The TUNEL assay of treated A549 cells showed that
the occurrence of apoptosis was easily detected following infection
with rL-RVG or NDV. As shown in Table III, compared with the PBS group,
the apoptotic rate was higher in the rL-RVG and NDV groups.
However, compared with the NDV and PBS groups, the apoptotic rate
was higher in the rL-RVG group, and there was a significant
difference between the rL-RVG and NDV groups (P<0.05) (Fig. 3C and Table III).
| Table IIIApoptotic indexes of A549 cells
infected by rL-RVG and NDV. |
Table III
Apoptotic indexes of A549 cells
infected by rL-RVG and NDV.
| Group | AI (n=10) | F-value | P-value |
|---|
| rL-RVG | 0.265±0.035a | | |
| NDV | 0.066±0.001b | 278.62 | <0.001 |
| PBS | 0.023±0.001 | | |
NDVs increase caspase expression
The intrinsic and extrinsic pathways converge
downstream at caspase-3. To examine whether the apoptosis induced
by rL-RVG depends on the caspase pathway, caspase-3 activation was
assessed in A549 cells following infection with rL-RVG or NDV. As
shown in Fig. 4Aa and Ba,
accumulation of cleaved caspase-3 in NDV-infected A549 cells was
observed at 24, 48 and 72 h. In addition, cleaved caspase-3 levels
in the rL-RVG group were higher than those in the NDV group. It was
assessed whether the extrinsic apoptotic pathway was involved in
tumor cells infected with rL-RVG or NDV. As illustrated in Fig. 4Ab and Bb, caspase-8 was activated
at 24, 48 and 72 h in A549 cells infected with rL-RVG or NDV. In
addition, caspase-8 activation in the rL-RVG group was higher than
that in the NDV group at 24, 48 and 72 h. These findings
demonstrated that rL-RVG infection induced the activation of
caspase-8 in tumor cells. Next, to determine the time course of
induction of the initiator caspase of the intrinsic apoptotic
pathway, A549 cells were infected with rL-RVG and NDV strains for
24, 48 and 72 h, respectively. Western blot analysis was used to
examine the activation of caspase-9. As shown in Fig. 4Ac and Bc, significant activation of
caspase-9 was induced at 24, 48 and 72 h. It is important that the
expression of caspase-9 in the rL-RVG group was higher than that in
the NDV group, which indicated that rL-RVG induced apoptosis via
the intrinsic pathway and the oncolytic effects were more evident
than those of NDV.
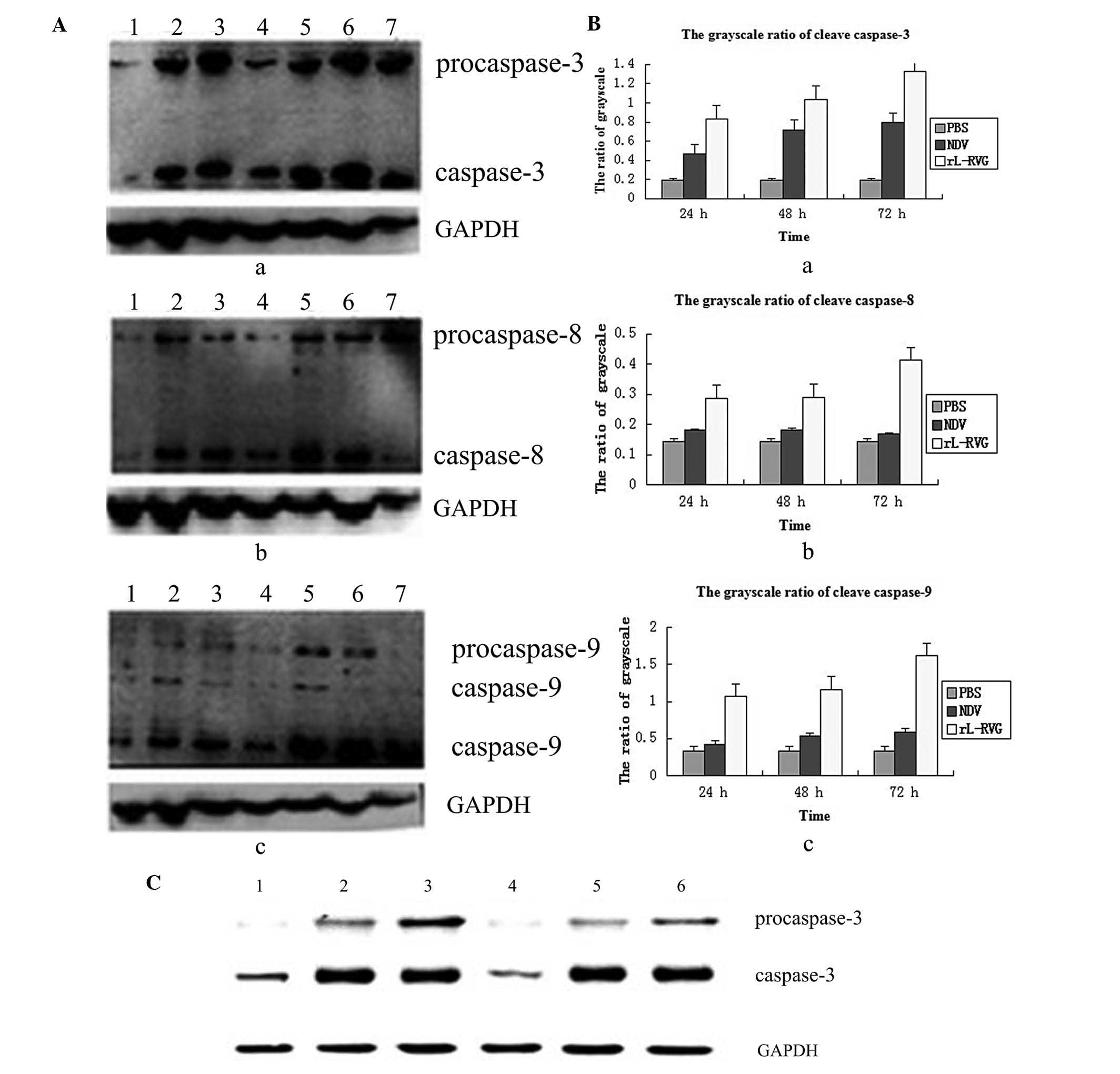 | Figure 4Expression of caspase induced by
virus. (A) Analysis of pro-apoptotic proteins in A549 cells
following infection. a, expression of caspase-3; b, expression of
caspase-8; c, expression of caspase-9. 1, PBS group; 2, NDV group
24 h; 3, NDV group 48 h; 4, NDV group 72 h; 5, rL-RVG group 24 h;
6, rL-RVG group 48 h; 7, rL-RVG group 72 h. (B) Grayscale ratio of
caspase. a, caspase-3 expression in the rL-RVG group was
significantly different from that in the other groups at any
time-point. b, caspase-8 expression in the rL-RVG group was
significantly different from that in the other groups, but there
were no significant differences between the NDV and PBS groups. c,
caspase-9 expression in the rL-RVG group was significantly
different from that in the other groups at any time-point. (C)
Following addition of pan-caspase inhibitor Z-VAD-FMK, the
expression of the caspase-3 protein was obviously reduced. 1, PBS
group; 2, NDV group; 3, rL-RVG group; 4, PBS + Z-VAD-FMK group; 5,
NDV + Z-VAD-FMK group; 6, rL-RVG + Z-VAD-FMK group. Z-VAD-FMK,
Z-Val-Ala-Asp-fluoromethylketone; rL-RVG, LaSota NDV strain
expressing rabies virus glycoprotein; NDV, Newcastle disease virus;
PBS, phosphate-buffered saline. |
To examine whether rL-RVG-induced apoptosis depended
on caspase activation, A549 cells were pretreated with the
broad-specificity caspase inhibitor Z-VAD-FMK and infected by
rL-RVG or NDV. The pan-caspase inhibitor Z-VAD-FMK was able to
effectively inhibit rL-RVG-triggered cytolysis at 24 h, indicating
that rL-RVG-mediated cytotoxicity was caspase-dependent. This
inhibitory effect was stronger than that of the NDV group, as shown
in Fig. 4C.
Expression of bcl-2 and bax
To examine whether the alteration of bcl-2 and bax
were induced by rL-RVG and NDV infection, the levels of bcl-2 and
bax were detected by western blot analysis (Fig. 5A). The levels of bcl-2 in
rL-RVG-infected and NDV-infected cells were not significantly
affected at 24, 48 and 72 h. However, the expression of bax in
rL-RVG-infected cells was higher than that in NDV-infected cells at
24, 48 and 72 h. Bcl-2 has been shown to inhibit the bax
oligomerization process. It has been proposed that the balance of
the bax/bcl-2 ratio serves as a regulator of cell survival vs.
death (14). The ratio of
bax/bcl-2 is shown in Fig. 5B. The
bax/bcl-2 ratio was significantly upregulated in rL-RVG-infected
cells at 24, 48 and 72 h. Thus, the pro-apoptotic protein dominates
in the apoptotic pathway.
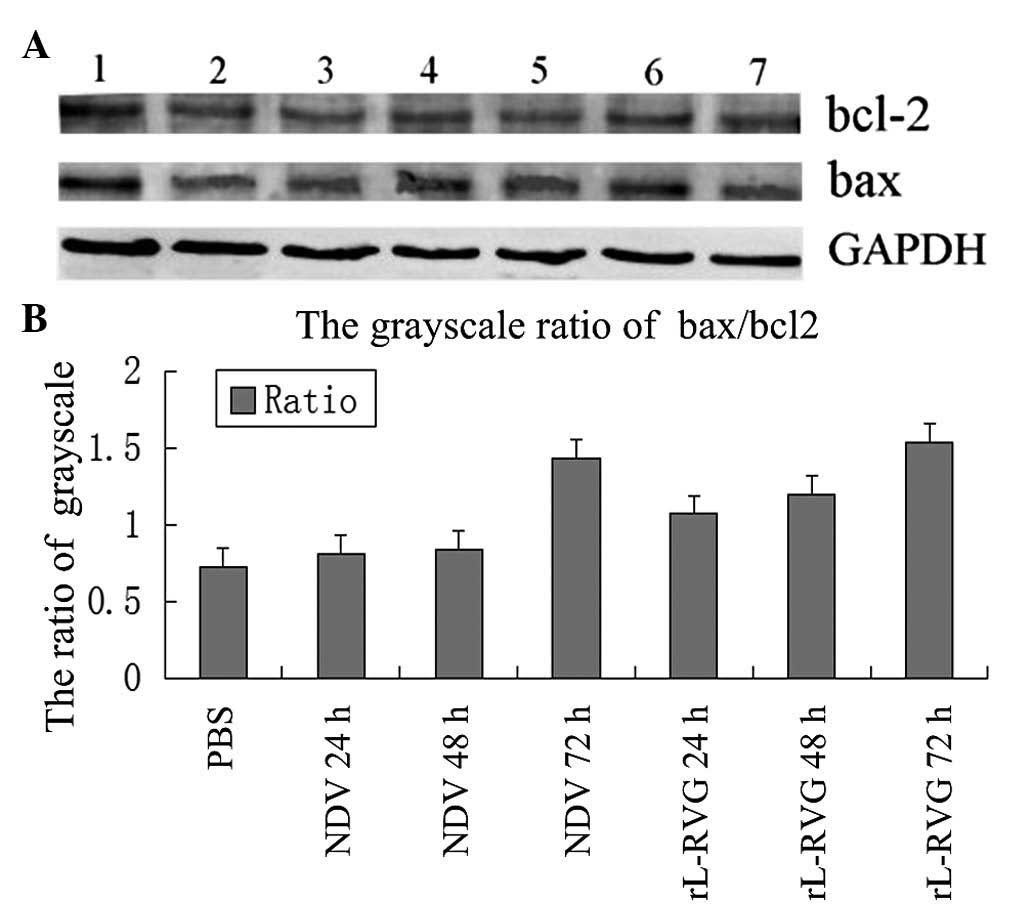 | Figure 5Expression of bax/bcl-2 in A549 cells
induced by virus. (A) Expression of bax and bcl-2. 1, PBS group; 2,
NDV group 24 h; 3, NDV group 48 h; 4, NDV group 72 h; 5, rL-RVG
group 24 h; 6, rL-RVG group 48 h; 7, rL-RVG group 72 h. (B) The
bax/bcl-2 ratio of the rL-RVG group was higher than that of the NDV
group at 24, 48 and 72 h following infection. bcl-2, B-cell
lymphoma 2; bax, bcl-2-associated X; rL-RVG, LaSota NDV strain
expressing rabies virus glycoprotein; NDV, Newcastle disease virus;
PBS, phosphate-buffered saline. |
Discussion
Oncolytic therapy is a novel biological treatment.
Oncolytic viruses can destroy harmful cells by infecting and
replicating in cancer cells while leaving normal cells unaffected
(15). In theory, the ability of
the NDV to kill tumor cells has higher efficiency and lower side
effects. Since the early 1950s, the NDV has been proven to be a
potential antitumor agent with promising results in clinical
trials. Numerous different studies focus on breeding effective
anti-tumor NDV strains and have produced 73-T (16), PV701 (17), MTH-68/H (18), HUJ (19) and several other potential
anti-neoplastic NDV strains (20).
Previous studies have shown that tumor cell growth was markedly
inhibited and cell apoptosis was induced by using adenovirus or
fowlpox viruses as vectors to express apoptin (21,22).
However, these gene therapy vectors did not have inherent
anti-tumor effects (23). By
applying a reverse genetics technique, it is possible to produce a
recombinant NDV. Compared with other vectors, NDV not only has the
ability of self-replication in tumor cells, but is also is a very
promising oncolytic agent. A study by Zhao et al (24) from 2008 supported the application
of a recombinant NDV/IL-2 virus as an anti-tumor agent.
The recombinant Newcastle disease virus rL-RVG,
which was kindly provided by the Harbin Veterinary Research
Institute (Harbin, China) (13),
was infected into A549 cells. PCR, western blot and
immunofluorescence analysis results showed that the rL-RVG was
stably expressed in A549 cells. In particular, RVG expression
enabled the NDV vector to spread from cell to cell and enhance its
replication in cell culture (13).
Western blot analysis showed that the NDV protein bands of the
rL-RVG group were stronger than those of the the NDV wild-type
strain group, and immunofluorescence results showed that more NDV
protein was present in the rL-RVG group than in the NDV group,
which may be due to RVG protein increasing the activity of NDV.
This may explain why rL-RVG had a higher oncolytic effect than the
wild-type NDV. Further study is required to understand the
underlying mechanism in detail.
Following infection of rL-RVG into A549 cells, cell
morphology and growth were strongly affected in the in vitro
experiment. The MTT assay indicated that compared with the PBS
group, the rL-RVG and NDV groups exerted a toxic effect in a time-
and concentration-dependent manner. Moreover, the toxic effect of
rL-RVG was stronger than that of the wild-type NDV strain. It would
be worthwhile exploring the underlying mechanisms leading to the
different potencies of the rL-RVG and wild-type NDV in further
studies.
Cell death commonly occurs due to cell necrosis and
apoptosis. The TUNEL experiment demonstrated a higher apoptotic
rate in the rL-RVG group as compared with that in the NDV and PBS
groups. Moreover, flow cytometry results showed that compared with
the PBS group, the rL-RVG and NDV groups exhibited a greater number
of early apoptotic cells, and the percentage of early apoptotic
cells in the rL-RVG group was higher than that in the NDV group
(P<0.05). All the results indicated that rL-RVG induced
apoptosis in A549 lung cancer cells. Therefore, it is concluded
that oncolysis of of tumor cells by rL-RVG was associated with its
ability to induce apoptosis.
Cell apoptosis is a basic biological function which
is crucial for the body to maintain a dynamic cell balance and
stability in the tissue. Studies have revealed delineation of
apoptotic pathways activated in NDV-infected cells (25). Multiple studies have investigated
the mechanisms underlying NDV-induced apoptosis. NDV can induce
apoptosis by activating both extrinsic and intrinsic apoptotic
pathways (26). Elankumaran et
al (27) have shown that in
tumor cells infected with NDV, apoptosis was triggered via the
extrinsic pathway, which was induced by caspase-8 activation
(27). However, such processes do
not occur in all tumor cells. In the present study, cleavage of
caspase-8 was also detected in the rL-RVG and NDV groups. Moreover,
levels of caspase-8 in the rL-RVG group were higher than those in
the wild-type NDV group. It may therefore be suggested that
compared with the NDV strain, rL-RVG induced stronger extrinsic
apoptosis. At the same time, intrinsic apoptotic pathways were
activated in the rL-RVG and NDV groups, which were induced by
caspase-9 activation. The caspase-3 inhibitor Z-VAD-FMK was able to
effectively inhibit NDV-triggered cytolysis following 24 h. This
demonstrated that rL-RVG-mediated and NDV-mediated cytotoxicity
were caspase-dependent, and that rL-RVG induced higher levels of
apoptosis than NDV.
By changing the outer membrane permeability and
mitochondrial activation, members of the bcl-2 protein family were
regulated to control cell apoptosis. The bcl-2 family members can
be divided into two categories according to their structure and
function: (1) Apoptosis-promoting members, such as Bax, which have
the ability to promote the opening of the mitochondrial
permeability transition pore, releasing caspase and promoting cell
apoptosis (28); and (2)
inhibitors of apoptosis, such as bcl-2, which protect the integrity
of the mitochondrial outer membrane and thereby prevent cytochrome
C release. Western blot analysis showed that the expression of
anti-apoptotic bcl-2 in the rL-RVG group did not change, but bax
expression was increased. Bcl-2 inhibits the bax oligomerization
process, and the ratio of bax/bcl-2 regulates cell survival vs.
death (14). The bax/bcl-2 ratio
was higher in the rL-RVG group than that in the NDV group at 24, 48
and 72 h. Accordingly, whether the involvement of viral protein or
soluble intermediate factors was responsible for the upregulation
of bax and downregulation of bcl-2 expression requires further
investigation.
The anti-cancer activity of rL-RVG in lung cancer
may have additional underlying mechanisms. For instance, the
198–214 amino acid sequence of rabies virus glycoprotein is highly
homologous with the 30–56 sequence in λ-bungatotoxin, which is able
to combine with nicotinic acetylcholine receptors (nAChRs). nAChRs
are considered to be combined with the rabies virus glycoprotein.
Plummer et al (29) showed
that α7-nAChR was widely expressed in lung cancer cells, including
squamous carcinoma, adenocarcinoma, large cell carcinoma and small
cell carcinoma. This therefore suggested that rL-RVG may be able to
be absorbed by tumor cells due to its affinity to α7-nAChR provided
by RVG.
In conclusion, rL-RVG is a novel type of oncolytic
virus and has a higher oncolytic efficacy than the previously
reported wild-type NDV. The underlying mechanism of action of the
recombinant rL-RVG strain proceeded via the induction of apoptosis
in NSCLC A549 cells. This induction of apoptosis was a
caspase-dependent process. The present study provided a strong
foundation for vaccine studies; however, the detailed mechanism
underlying the anti-cancer activity of rL-RVG requires further
study. The present study provided strong evidence for the potential
of rL-RVG to be used for clinical treatment of cancer.
Acknowledgements
The authors would like to thank Professor Zhigao Bu
and Dr Jinying Ge from the State Key Laboratory of Veterinary
Biotechnology, Harbin Veterinary Research Institute, Chinese
Academy of Agricultural Sciences, Harbin, China for suggestions
regarding the experiments performed. The authors would also like to
thank Dr Yan Tao and Aihua Gong from Jiangsu University (Jiangsu,
China) for kindly providing technical support.
References
|
1
|
Jemal A, Tiwari RC, Murray T, et al:
American Cancer Society: Cancer statistics, 2004. CA Cancer J Clin.
54:8–29. 2004. View Article : Google Scholar
|
|
2
|
DeSantis C, Naishadham D and Jemal A:
Cancer statistics for African Americans, 2013. CA Cancer J Clin.
63:151–166. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhang J: The treatment of lung carcinoma
in aged people. Chin J Geriatr. 4:316–318. 2001.
|
|
4
|
Vähä-Koskela MJ, Heikkilä JE and Hinkkanen
AE: Oncolytic viruses in cancer therapy. Cancer Letters.
254:178–216. 2007.
|
|
5
|
Lam HY, Yeap SK, Rasoli M, et al: Safety
and clinical usage of newcastle disease virus in cancer therapy. J
Biomed Biotechnol. 2011:7187102011.PubMed/NCBI
|
|
6
|
Yusoff K and Tan WS: Newcastle disease
virus: macromolecules and opportunities. Avian Pathol. 30:439–455.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Römer-Oberdörfer A, Werner O, Veits J,
Mebatsion T and Mettenleiter TC: Contribution of the length of the
HN protein and the sequence of the F protein cleavage site to
Newcastle disease virus pathogenicity. J Gen Virol. 84(Pt 11):
3121–3129. 2003.PubMed/NCBI
|
|
8
|
Wu Y, Zhang X, Wang X, Wang L, Hu S, Liu X
and Meng S: Apoptin enhances the oncolytic properties of Newcastle
disease virus. Intervirology. 55:276–86. 2012.PubMed/NCBI
|
|
9
|
Silverstein SC and Marcus PI: Early stages
of newcastle disease virus-hela cell interaction: an electron
microscopic study. Virology. 23:370–380. 1964. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Johnson N, McElhinney LM, Smith J, Lowings
P and Fooks AR: Phylogenetic comparison of the genus lyssavirus
using distal coding sequences of the glycoprotein and nucleoprotein
genes. Arch Virol. 147:2111–2123. 2002. View Article : Google Scholar
|
|
11
|
Préhaud C, Lay S, Dietzschold B and Lafon
M: Glycoprotein of nonpathogenic rabies viruses is a key
determinant of human cell apoptosis. J Virol. 77:10537–10547.
2003.PubMed/NCBI
|
|
12
|
Ge J, Wang X, Tao L, et al: Newcastle
disease virus-vectored rabies vaccine is safe, highly immunogenic,
and provides long-lasting protection in dogs and cats. J Virol.
85:8241–8252. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Reed LJ and Muench H: A simple method of
estimating fifty percent endpoints. Am J Hyg. 27:493–497. 1938.
|
|
14
|
Molouki A, Hsu YT, Jahanshiri F, Rosli R
and Yusoff K: Newcastle disease virus infection promotes bax
redistribution to mitochondria and cell death in HeLa cells.
Intervirology. 53:87–94. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ali R, Alabsi AM, Ali AM, et al: Cytolytic
effects and apoptosis induction of Newcastle disease virus strain
AF2240 on anaplastic astrocytoma brain tumor cell line. Neurochem
Res. 36:2051–2062. 2011. View Article : Google Scholar
|
|
16
|
Batliwalla FM, Bateman BA, Serrano D, et
al: A 15-year follow-up of AJCC stage III malignant melanoma
patients treated postsurgically with Newcastle diseasevirus (NDV)
oncolysate and determination of alterations in the CD8 T cell
repertoire. Mol Med. 4:783–794. 1998.
|
|
17
|
Pecora AL, Rizvi N, Cohen GI, et al: Phase
I trial of intravenous administration of PV701, an oncolytic virus,
in patients with advanced solid cancers. J Clin Oncol.
20:2251–2266. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Csatary LK, Moss RW, Beuth J, Töröcsik B,
Szeberenyi J and Bakacs T: Beneficial treatment of patients with
advanced cancer using a Newcastle disease virus vaccine (MTH-68/H).
Anticancer Res. 19:635–638. 1999.PubMed/NCBI
|
|
19
|
Freeman AI, Zakay-Rones Z, Gomori JM, et
al: Phase I/II trial of intravenous NDV-HUJ oncolytic virus in
recurrent glioblastoma multiforme. Mol Ther. 13:221–228. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ravindra PV, Tiwari AK, Sharma B and
Chauhan RS: Newcastle disease virus as an oncolytic agent. Indian J
Med Res. 130:507–513. 2009.PubMed/NCBI
|
|
21
|
Li X, Jin N, Mi Z, et al: Antitumor
effects of a recombinant fowlpox virus expressing apoptin in vivo
and in vitro. Int J Cancer. 119:2948–2957. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Pietersen AM, van der Eb MM, Rademaker HJ,
et al: Specific tumor-cell killing with adenovirus vectors
containing the apoptin gene. Gene Ther. 6:882–892. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wu Y, Zhang X, Wang X, et al: Apoptin
enhances the oncolytic properties of Newcastle disease virus.
Intervirology. 55:276–286. 2012.PubMed/NCBI
|
|
24
|
Zhao H, Janke M, Fournier P and
Schirrmacher V: Recombinant Newcastle disease virus expressing
human interleukin-2 serves as a potential candidate for tumor
therapy. Virus Res. 136:75–80. 2008. View Article : Google Scholar
|
|
25
|
Ravindra PV, Tiwari AK, Ratta B, et al:
Induction of apoptosis in Vero cells by Newcastle disease virus
requires viral replication, de-novo protein synthesis and caspase
activation. Virus Res. 133:285–90. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zamarin D and Palese P: Oncolytic
Newcastle disease virus for cancer therapy: old challenges and new
directions. Future Microbiol. 7:347–367. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Elankumaran S, Rockemann D and Samal SK:
Newcastle disease virus exerts oncolysis by both intrinsic and
extrinsic caspase-dependent pathways of cell death. J Virol.
80:7522–7534. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hirsch T, Marzo I and Kroemer G: Role of
the mitochondrial permeability transition pore in apoptosis. Biosci
Rep. 17:67–76. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Plummer HK III, Dhar M and Schuller HM:
Expression of the alpha7 nicotinic acetylcholine receptor in human
lung cells. Respir Res. 6:292005. View Article : Google Scholar : PubMed/NCBI
|














