Introduction
Hepatocellular carcinoma (HCC) is the third most
common cause of cancer-associated mortality worldwide. The HCC
incidence rate is increasing more rapidly than most other types of
cancers due to the increasing prevalence of hepatitis B and C viral
infections (1,2). Therapies for HCC include surgery,
radiofrequency ablation and chemotherapy. At present, no drugs are
available to prevent or reduce tumor spread and/or recurrence,
which severely affects prognosis and survival. HCC is the only type
of cancer in which ‘conventional’ chemotherapeutics must be
avoided, due to the underlying liver damage. Thus, a novel drug for
the treatment of HCC is urgently required.
Tumor angiogenesis is a complex process in which new
blood vessels are formed in response to interactions between tumor
cells and endothelial cells (ECs) (3,4).
Angiogenesis has an important role in cancer growth and metastasis,
with tumors able to grow autonomously to a size of 2±3
mm3, but incapable of growing beyond this size without a
blood supply (5–9). Tumor cells continuously stimulate the
formation of new blood vessels to grow. New blood vessels that
surround tumor tissues provide a gateway for the tumor cells to
enter the vascular system and metastasize to distant organs. Thus,
the inhibition of angiogenesis has been proposed to suppress tumor
growth and metastasis (10). An
increasing number of studies have revealed that tumor cells secrete
angiogenic growth factors to stimulate EC proliferation and to
induce angiogenesis (6). Among
them, vascular endothelial growth factor (VEGF) is one of the most
potent angiogenic factors and it is overexpressed in numerous types
of human cancer (11).
Genetic studies have shown that Notch signaling has
a critical role in vascular formation during early embryonic
development (12–14). Notch signaling is an evolutionarily
conserved pathway, which regulates cell fate decisions during the
embryonic development of vertebrates and invertebrates (15). There are four mammalian Notch
genes, Notch1-4, and five ligands, jagged 1, jagged 2,
delta-like ligand (DLL) 1, DLL3 and DLL4 (12,16,17).
Notch signaling has been reported to have a crucial role in tumor
angiogenesis and is a novel target for anti-angiogenic therapy.
Studies have demonstrated that the activation of the Notch
signaling pathway inhibits angiogenesis. Activated Notch1 and
Notch4 have been reported to decrease the expression of VEGF
receptor (VEGFR) 2, resulting in decreased human EC proliferation
in response to VEGF (18). Human
ECs transduced with DLL4 have been shown to exhibit decreased
VEGF-induced proliferation and migration, as well as a significant
reduction in the expression of VEGFR2 and its coreceptor
neuropilin-1, suggesting that DLL4 is an inhibitor of angiogenesis
(19). In addition, inhibiting
jagged using an antisense oligomer in bovine ECs has been found to
induce tube formation and collagen gel invasion in response to
fibroblast growth factor, but not VEGF (20). Although Notch signaling has been
reported to inhibit sprouting angiogenesis in these studies
(18–20), most were performed in vitro.
Thus, in vivo tumor angiogenesis assays are required to
confirm these findings and to determine their physiological
relevance.
Rubus alceifolius Poir is a natural product,
which is used in Traditional Chinese Medicine in Southern regions
of Fujian Province, China, to treat various types of hepatitis. In
a previous study, our group identified that total alkaloids of
Rubus alceifolius Poir (TARAP) promoted apoptosis in human
HCC cells (21). However, the
specific mechanism underlying the anticancer effect of TARAP in
vivo has yet to be elucidated. In the present study, in order
to further investigate the anticancer activity of TARAP, the effect
of TARAP on tumor growth and its underlying molecular mechanisms
were investigated in a HCC xenograft mouse model.
Materials and methods
Materials and reagents
Dulbecco’s modified Eagle’s medium (DMEM), fetal
bovine serum (FBS), penicillin-streptomycin, Trypsin-EDTA and
TRIzol® reagent were purchased from Life Technologies
(Carlsbad, CA, USA). SuperScript® II reverse
transcriptase was purchased from Promega Corporation (Madison, WI,
USA). Antibodies against cluster of differentiation (CD) 31 (human,
pAb), VEGF-A (human, pAb), VEGFR2 (human, pAb), Notch (human, mAb),
Dll4 (human, pAb) and jagged 1 (human, pAb) were obtained from
Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA). All
antibodies were diluted 1:100. All other chemicals, unless
otherwise stated, were purchased from Sigma-Aldrich (St. Louis, MO,
USA).
TARAP preparation
TARAP preparation was performed as described
previously (21). The roots of
Rubus alceifolius Poir were collected in Anxi (China) and
were identified and authenticated at Fujian University of
Traditional Chinese Medicine (Fuzhou, China). The alkaloids were
then extracted as follows: The herb powder (1 g) was extracted
using 50 ml chloroform:methanol:ammonia solution (15:4:3) for 2 h
in an ice bath, sonicated for 30 min, brought to room temperature
and filtered. The filtered solution was collected and dessicated.
The resultant residue was dissolved in 2 ml of 2% sulfuric acid
solution and filtered. The filter paper and residue were re-washed
with 2 ml of 2% sulfuric acid solution and buffer solution (pH
3.6). The buffer was then added to make a final volume of 50 ml,
and the solution was saved for future use. Acid dye colorimetry was
used to measure the total alkaloid content. The total alkaloid
content was 0.81 mg alkaloid per gram of initial herb powder.
Cell culture and animals
HepG2 human HCC cells were purchased from the
American Type Culture Collection (Rockville, MD, USA). Cells were
grown in DMEM containing 10% (v/v) FBS, 100 U/ml penicillin and 100
μg/ml streptomycin in a humidified incubator at 37°C with 5%
CO2. Cells were subcultured to 80–90% confluency.
Six-week-old male BALB/c athymic nude mice were purchased from
Shanghai Si-Lai-Ke Experimental Animal Ltd. (Shanghai, China). Mice
were housed at five mice per cage in a pathogen-free room in an
environment with controlled temperature (22°C), humidity and a 12-h
light/dark cycle with free access to water and a standard
laboratory diet. All animal treatments were performed in accordance
with the Guidelines for Animal Experimentation of Fujian University
of Traditional Chinese Medicine (Fuzhou, China). The animal studies
were approved by the Fujian Institute of Traditional Chinese
Medicine Animal Ethics Committee (Fuzhou, China).
In vivo tumor xenograft study
HepG2 cells were seeded at a density of
4×106 cells mixed 1:1 with Matrigel™ and were
subcutaneously injected in the right flank of the mice to initiate
tumor growth. Seven days following xenograft implantation, when the
tumors reached 3 mm in diameter, the mice were randomly divided
into two groups (n=10) and were intragastrically administered 3
g/kg body weight TARAP or saline daily, five days a week for 21
days. At the end of the experiment, the animals were anaesthetized
with ether and the mice were sacrificed by cervical vertebra
dislocation, and the tumor tissue was removed. A section of the
tumor was fixed in buffered formalin and the remaining sections
were stored at −80°C for molecular analyses.
RNA extraction and reverse
transcription-polymerase chain reaction (PCR) analysis
Total RNA was isolated using TRIzol Reagent.
Oligo(dT)-primed RNA (1 μg) was reverse-transcribed using
SuperScript II reverse transcriptase according to the
manufacturer’s instructions. The cDNA product (1 μl) was added to
prepare the PCR reaction mixture containing 200 μM dNTPs, 1.5 mM
MgCl2 and 1.25 U Platinum Taq polymerase for VEGF-A,
VEGFR2, Notch, Dll4 or jagged1 with a Taq DNA polymerase. GAPDH was
used as an internal control. Samples were analyzed using gel
electrophoresis on a 1.5% agarose gel. DNA bands were analyzed
using a Bio-Rad Gel Doc 2000 System (Bio-Rad, Hercules, CA,
USA).
Immunohistochemistry (IHC)
Tumor samples were fixed with 10% formaldehyde for
12 h and processed for the generation of paraffin-embedded tumor
slides. Slides were subjected to antigen retrieval and endogenous
peroxidase activity was blocked using hydrogen peroxide. Sections
were then incubated with CD31, VEGF-A, VEGFR2, Notch, Dll4 and
jagged 1 antibodies diluted 1:100. Subsequent to washing with
phosphate-buffered saline (PBS), slides were incubated with
biotinylated secondary antibodies followed by incubation with
conjugated horseradish peroxidase-labeled streptavidin (Dako UK
Ltd., Ely, UK) and washed with PBS. Slides were then incubated with
diaminobenzidine as the chromogen, followed by counterstaining with
hematoxylin. Subsequent to staining, five high-power fields
(magnification, ×400) were randomly selected in each slide and the
average proportion of positive cells in each field was counted
using the true color multi-functional cell image analysis
management system (Image-Pro Plus; Media Cybernetics, Inc.,
Bethesda, MD, USA). To exclude any non-specific staining, PBS was
used to replace the primary antibody as a negative control.
Statistical analysis
All data are presented as the mean ± standard
deviation. Data were analyzed using the SPSS package for Windows
(Version 11.5; SPSS, Inc., Chicago, IL, USA). Statistical analysis
of the data was performed using the Student’s t-test. P<0.05 was
considered to indicate a statistically significant difference.
Results and Discussion
TARAP inhibits tumor angiogenesis in a
xenograft model
Deregulated angiogenesis has a crucial role in the
development of various diseases, including cancer (6,22,23).
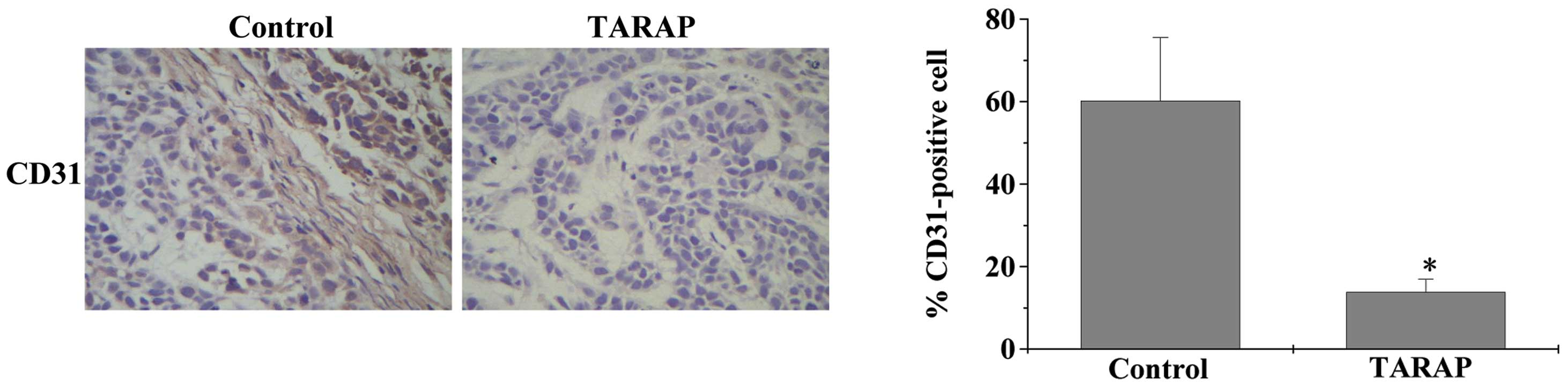
CD31 is a characteristic molecule in vascular ECs which is used as
an angiogenesis marker in vivo. In the present study,
immunohistochemical staining of CD31 was used to identify the
degree of angiogenesis. As shown in Fig. 1A and B, the microvessel density of
the HCC xenografts in the TARAP-treated mice was observed to be
reduced compared with that in the control mice, suggesting that
TARAP inhibited angiogenesis in vivo. Thus, TARAP may
mediate its antitumor activity through blocking angiogenesis.
TARAP inhibits the expression of VEGF-A
and VEGFR2 in HCC xenograft mice
In the process of tumor growth, interaction between
tumor cells and vascular ECs causes increases in tumor blood
vessels and promotes tumor growth (24,25).
VEGF-A is commonly expressed in a wide variety of types of human
tumor cells, is one of the most effective biological inducers of
angiogenesis and is associated with angiogenesis, growth and
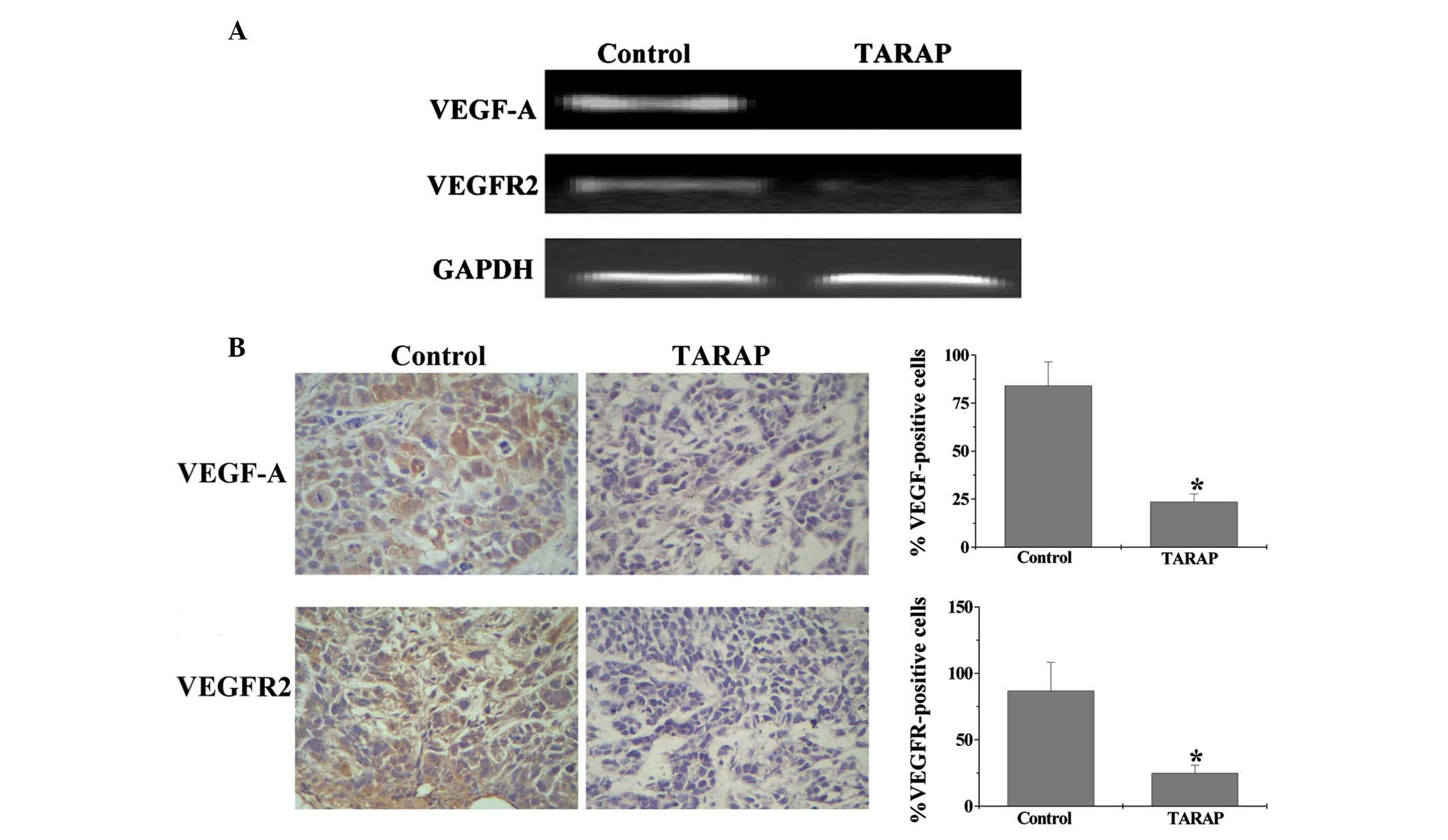
metastasis (26–29). To analyze the effect of TARAP on
tumor angiogensis, VEGF-A and VEGFR2 expression were assessed in a
xenograft model using PCR analysis and IHC. PCR analysis revealed
that TARAP treatment reduced the mRNA expression of VEGF-A and
VEGFR2 in the HCC xenografts (Fig.
2A). Furthermore, IHC showed that the protein expression of
VEGF-A and VEGFR2 was similar to their respective mRNA expression
levels. The percentage of VEGF-A- and VEGFR2-positive cells in the
control group was found to be 84±12.34 and 86.67±11.48%,
respectively, while that in TARAP-treated mice was 23.5±4.31 and
24.83±5.79%, respectively (Fig.
2B).
TARAP suppresses the Notch signaling
pathway in HCC xenograft mice
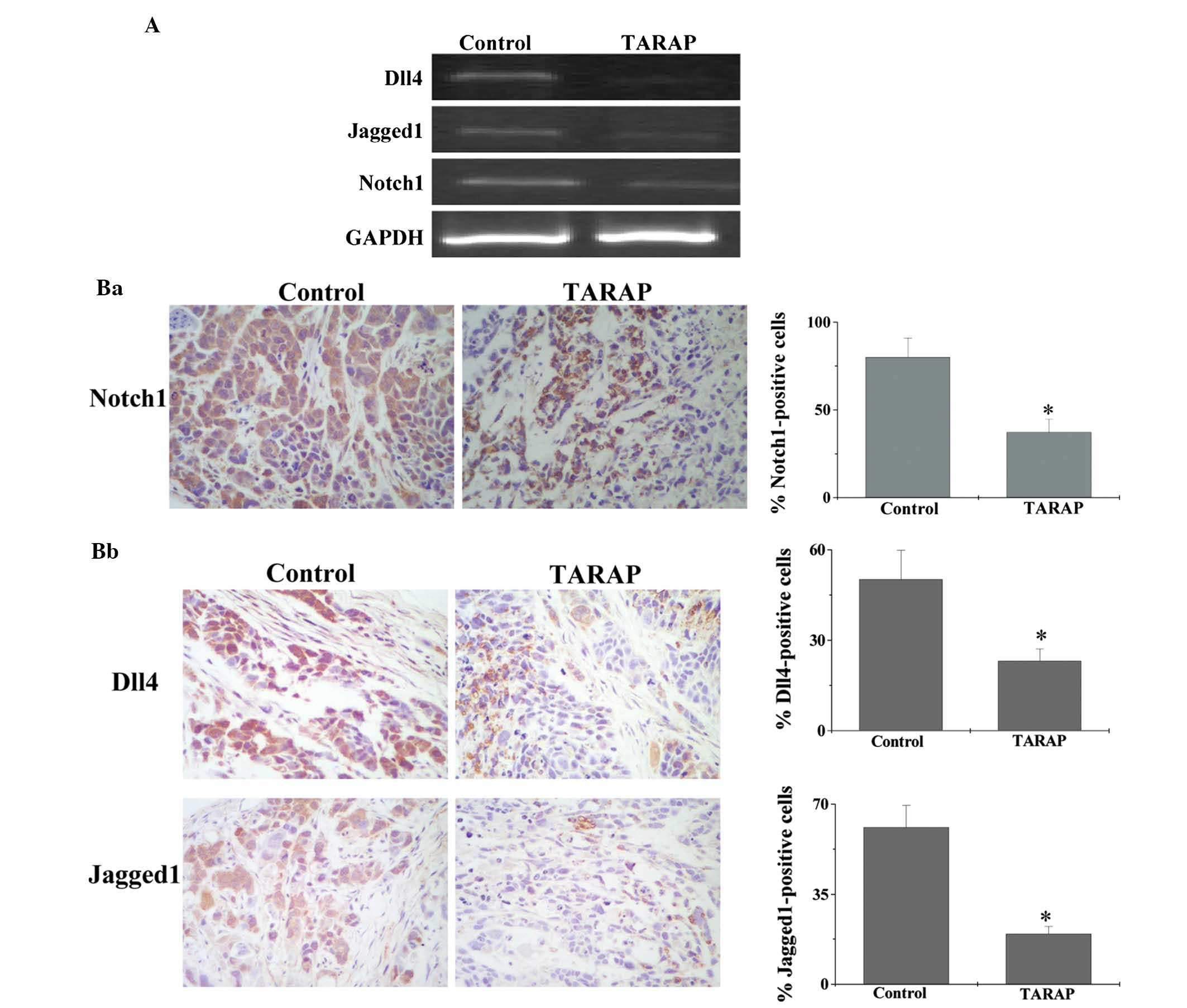
Tumor angiogenesis is tightly regulated by the Notch
signaling pathway; therefore, in the present study, the effect of
TARAP on the expression of key mediators of the Notch signaling
pathway was investigated using PCR and IHC analyses. As shown in
Fig. 3A, TARAP treatment was
observed to significantly reduce the mRNA expression of Notch, Dll4
and jagged 1 in tumor tissues. Furthermore, their protein
expression was significantly inhibited by TARAP treatment. The
percentages of Notch-, Dll4- and jagged 1-positive cells in the
control group were 80±10.96, 50.17±9.68 and 61±8.42%, respectively,
whereas those in TARAP-treated mice were 37.33±7.32, 23.17±3.99 and
19.5±3.1%, respectively (Fig. 3B).
These findings suggested that the inhibitory effect of TARAP on
tumor angiogenesis in vivo may be mediated through the
suppression of the Notch signaling pathway.
In conclusion, to the best of our knowledge, the
present study was the first study to show that TARAP inhibits tumor
angiogenesis through the suppression of the Notch signaling
pathway, which may in part explain its anticancer activity.
Acknowledgements
The present study was supported by grants from the
Nature Science Foundation of Fujian Province of China (nos.
2010J01191 and 2010J01194) and the Medical Originality Foundation
of Fujian Province of China (no. 2009-CX-18).
References
|
1
|
Tsai WL and Chung RT: Viral
hepatocarcinogenesis. Oncogene. 29:2309–2324. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Schütte K, Bornschein J and Malfertheiner
P: Hepatocellular carcinoma - epidemiological trends and risk
factors. Dig Dis. 27:80–92. 2009.PubMed/NCBI
|
|
3
|
Bikfalvi A: Significance of angiogenesis
in tumour progression and metastasis. Eur J Cancer. 31A:1101–1104.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Folkman J: What is the evidence that
tumors are angiogenesis dependent? J Natl Cancer Inst. 82:4–6.
1990. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ingber D, Fujita T, Kishimoto S, Sudo K,
Kanamaru T, Brem H and Folkman J: Synthetic analogues of fumagillin
that inhibit angiogenesis and suppress tumour growth. Nature.
348:555–557. 1990. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Folkman J and Shing Y: Angiogenesis. J
Biol Chem. 267:10931–10934. 1992.
|
|
7
|
Pluda JM: Tumor-associated angiogenesis:
mechanisms, clinical implications, and therapeutic strategies.
Semin Oncol. 24:203–218. 1997.PubMed/NCBI
|
|
8
|
Folkman J: Anti-angiogenesis: new concept
for therapy of solid tumors. Ann Surg. 175:409–416. 1972.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Brem H and Folkman J: Inhibition of tumor
angiogenesis mediated by cartilage. J Exp Med. 141:427–439. 1975.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Voest EE: Inhibitors of angiogenesis in a
clinical perspective. Anticancer Drugs. 7:723–727. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Jung YD, Ahmad SA, Liu W, Reinmuth N,
Parikh A, Stoeltzing O, Fan F and Ellis LM: The role of the
microenvironment and intercellular cross-talk in tumor
angiogenesis. Semin Cancer Biol. 12:105–112. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Krebs LT, Xue Y, Norton CR, et al: Notch
signaling is essential for vascular morphogenesis in mice. Genes
Dev. 14:1343–1352. 2000.PubMed/NCBI
|
|
13
|
Krebs LT, Shutter JR, Tanigaki K, Honio T,
Stark KL and Gridley T: Haploinsufficient lethality and formation
of arteriovenous malformations in Notch pathway mutants. Genes Dev.
18:2469–2473. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Xue Y, Gao X, Lindsell CE, et al:
Embryonic lethality and vascular defects in mice lacking the Notch
ligand Jagged1. Hum Mol Genet. 8:723–730. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Artavanis-Tsakonas S, Rand MD and Lake RJ:
Notch signaling: cell fate control and signal integration in
development. Science. 284:770–776. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Guo S, Liu M and Gonzalez-Perez RR: Role
of Notch and its oncogenic signaling crosstalk in breast cancer.
Biochim Biophys Acta. 1815:197–213. 2011.PubMed/NCBI
|
|
17
|
Li JL and Harris AL: Notch signaling from
tumor cells: a new mechanism of angiogenesis. Cancer Cell. 8:1–3.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Taylor KL, Henderson AM and Hughes CC:
Notch activation during endothelial cell network formation in vitro
targets the basic HLH transcription factor HESR-1 and downregulates
VEGFR-2/KDR expression. Microvasc Res. 64:372–383. 2002. View Article : Google Scholar
|
|
19
|
Williams CK, Li JL, Murga M, Harris AL and
Tosato G: Upregulation of the Notch ligand Delta-like 4 inhibits
VEGF-induced endothelial cell function. Blood. 107:931–939. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zimrin AB, Pepper MS, McMahon GA, Nguyen
F, Montesano R and Maciag T: An antisense oligonucleotide to the
notch ligand jagged enhances fibroblast growth factor-induced
angiogenesis in vitro. J Biol Chem. 271:32499–32502. 1996.
View Article : Google Scholar
|
|
21
|
Zhao J, Chen X, Lin W, et al: Total
alkaloids of Rubus aleaefolius Poir inhibit hepatocellular
carcinoma growth in vivo and in vitro via activation
of mitochondrial-dependent apoptosis. Int J Oncol. 42:971–978.
2013.
|
|
22
|
Folkman J: Angiogenesis in cancer,
vascular, rheumatoid and other diseases. Nat Med. 1:27–31. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Folkman J: Angiogenesis. Annu Rev Med.
57:1–18. 2006. View Article : Google Scholar
|
|
24
|
Yancopoulos GD, Davis S, Gale NW, et al:
Vascular-specific growth factors and blood vessel formation.
Nature. 407:242–248. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ishiwata I, Sudo T, Kiguchi K and Ishikawa
H: Tumor angiogenesis factors produced by cancer cells. Hum Cell.
12:37–46. 1999.PubMed/NCBI
|
|
26
|
Risau W: Mechanisms of angiogenesis.
Nature. 386:671–674. 1997. View
Article : Google Scholar : PubMed/NCBI
|
|
27
|
Jain RK: Tumor angiogenesis and
accessibility: role of vascular endothelial growth factor. Semin
Oncol. 29(6 Suppl 16): 3–9. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ferrara N: Role of vascular endothelial
growth factor in physiologic and pathologic angiogenesis:
therapeutic implications. Semin Oncol. 29(6 Suppl 16): 10–14. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Houck KA, Ferrara N, Winer J, et al: The
vascular endothelial growth factor family: identification of a
fourth molecular species and characterization of alternative
splicing of RNA. Mol Endocrinol. 5:1806–1814. 1991. View Article : Google Scholar : PubMed/NCBI
|