Introduction
The clinical manifestations of acute spinal cord
injuries (ASCIs) mainly appear as quadriplegia or paraplegia. In
addition to primary injuries, ASCIs are also caused by secondary
injuries, including ischemia, hypoxia, the inflammatory response,
free radical formation and excitotoxicity, which could take several
hours or days to manifest following the primary injuries. These
secondary injuries can in turn result in the formation of severe
spinal cord injuries (SCIs), for which no effective treatment is
available to date (1). Stem cell
transplantation is currently the most promising treatment method
for SCIs. In particular, bone marrow mesenchymal stem cells (BMSCs)
are considered highly promising in SCI clinical therapy and cell
transplantation (2,3). In addition to their wide availability
and relative ease of isolation and expansion, BMSCs are able to
differentiate into neurons (4).
However, transplanting BMSCs into the SCI site and inducing them to
differentiate into neurons remains a challenge and has yielded a
number of inconsistent results in animal models (5,6).
These results were associated with the deficiency of neurotrophic
factors in the local microenvironment during SCI repair. Therefore,
the importance of improving the local neurotrophic factor levels
and creating a gentle microenvironment for the damaged spinal cord
sites has become apparent in the study of SCIs.
Neurotrophin-3 (NT-3), a member of the neurotrophin
family, is involved in the neuroprotection and nutrition of various
neurons and thus promotes the survival and axonal regeneration of
damaged neurons (7). However, due
to its relatively short half-life (8) and its rapid metabolism when combined
with BMSCs, NT-3 cannot sustain its effects on SCIs. In order to
address this problem, a combined treatment involving the
transfection of neurotrophic factor genes into BMSC cells was
designed. This combination allows the sustained secretion of
neurotrophic factors, improves the survival of BMSCs and is
involved in induced neuronal differentiation in order to maximize
the therapeutic effects in SCIs. The current study aimed to
transfect the NT-3 gene in vitro into cultured Sprague
Dawley (SD) rat BMSCs using a lentivirus-mediated method in order
to determine the efficiency of NT-3 gene-BMSC transfection and NT-3
gene expression in BMSCs.
Materials and methods
Plasmid construction
The attB1-NT3-attB2 sequence was amplified using the
overlapping polymerase chain reaction (PCR) method. The primers
used were as follows: attB1-Kozak-NT3-P, forward 5-′ATG TCC ATC TTG
TTT TAT GTG ATA TTT C-3′ and reverse 5′-TCA TGT TCT TCC GAT TTT TCT
TGA C-3′. The PCR products were then subjected to 1% agarose gel
electrophoresis. Subsequently, the DNA was recycled with the
agarose gel electrophoresis recycling kit, according to the
manufacturer’s instructions (Qiagen Inc., Valencia, CA, USA) and
stored at −20°C.
Gateway cloning was performed to construct the entry
clone. pDown-NT3 was added to the pDonr221 plasmid, BP clonase and
attB1-NT3-attB2. The mixture was incubated at 25°C and was then
reacted with BP for 3 h. Proteinase K (Cyagen Biosciences Inc.,
Guagzhou, China) was added for 10 min to terminate the reaction.
Subsequently, the BP reaction products were transferred into the
Escherichia coli strain DH5α (Cyagen Biosciences Inc.).
Following this, 100 μl of the transformed product was coated onto a
lysogeny broth plate with 50 μg/ml Kan and subsequently incubated
overnight at 37°C. Colony PCR was used to screen the positive
plasmid clones, which were then used for positive clone
sequencing.
Gateway cloning was performed to construct the
recombinant target plasmid, PLV.Ex3d.P-neo-EF1A-NT3-internal
ribosome entry site (IRES)-enhanced green fluorescent protein
(EGFP). Minor extractions of the pUp-EF1A, pDown-NT3,
pTail-IRES-EGFP, LR clonase and PLV.Des3d.P-neo (Tiangen Biotech
(Beijing) Co., Ltd., Beijing, China) plasmids were performed, and
the extracts were incubated at 25°C. Subsequently, the incubated
samples were subjected to LR reaction for 3 h. Proteinase K was
added for 10 min to terminate the reaction. The LR reaction
products were then transferred into the E. coli strain DH5α.
Colony PCR was used to screen the positive plasmid clones, which
were then used for positive clone sequencing.
Lentivirus packaging
293FT cells (Cyagen Biosciences Inc.) in the
logarithmic growth phase and in good condition were obtained.
Following digestion and counting, the cells were seeded in a 10 cm
dish at a concentration of 5×106 cells/dish and then
incubated overnight at 37°C and 5% CO2. Prior to
transfection, the used culture solution was removed and 5 ml of 10%
fresh Dulbecco’s modified Eagle’s medium was added to prepare a
DNA-Lipofectamine 2000 complex (Invitrogen Life Technologies,
Carlsbad, CA, USA). The quantity of complex in the 10 cm dish was
used as a reference. The DNA-Lipofectamine 2000 complex was added
dropwise onto the 293FT cells and the dish was agitated to achieve
mixing. Subsequently, the mixture was incubated overnight at 37°C
in a 5% CO2-humidified incubator (NuAire, Inc.,
Plymouth, MN, USA). Approximately 48 h after the transfection, the
cell supernatant of the transfected 293FT cells (in the logarithmic
growth phase) was collected and concentrated, and then was added to
a 10 ml culture dish for continuous cultivation. After 72 h, the
cell supernatant was collected and reconcentrated and the virus was
subpacked. The sample was then stored at −80°C for use in the
determination of the biological virus titer. The virus titers were
preliminarily determined according to their fluorescent
expression.
Cultivation, passage and
lentiviral-stable transfection of SD rat BMSCs
An SD rat was sacrificed via cervical dislocation.
The whole body was then immersed in 75% ethanol for 30 min. The
bilateral femur and tibia were obtained under sterile conditions
and washed with the medium. Subsequently, the samples were prepared
into single-cell suspensions. The cell concentration was adjusted
and the sample was placed in an incubator at 37°C. Cell passage was
performed when 80–90% of the cells were fused. The present study
was conducted in strict accordance with the recommendations in the
Guide for the Care and Use of Laboratory Animals of the National
Institutes of Health (Bethesda, MD, USA). The animal use protocol
has been reviewed and approved by the Institutional Animal Care and
Use Committee of the Second Affiliated Hospital of Dalian Medical
University (Dalian, China).
Fifth-generation BMSCs were obtained for digestion
and were centrifuged at 168 × g, for 5 min. Subsequently, the
supernatant was discarded and the cell precipitate was resuspended.
Several samples were obtained for counting and the cells were
seeded into a six-well plate at a density of 1×105
cells/9.6 cm2. When the cells had completely adhered to
the wall, the medium was replaced with 1 ml of a fresh SD MSC
complete medium (Gibco-BRL, Carlsbad, CA, USA). The cells in two
wells were then added with 40 and 60 μl NT-3 and EGFP-carrying
lentivirus, respectively. Following complete mixing by agitation,
the samples were incubated at 37°C, 5% CO2 and 95%
relative humidity. Approximately 8 h after the transduction, the SD
BMSC complete medium was replaced and transduction was continued
for 48 h. Following this, transduction results for the samples were
determined using an inverted fluorescence microscope (Olympus,
Tokyo, Japan). Overgrown cells were inoculated onto a six-well
plate for drug screening using two 0.5 and 1.0 mg/ml G418 solutions
(Shanghai Fushen Biotechnology Co., Ltd., Shanghai, China). The
medium was replaced every 2 days. The expression of green
fluorescent protein (GFP) was observed using inverted fluorescence
microscopy based on the color of the medium and the cell state.
Following 7 days of screening, the complete medium without G418 was
replaced and cultivation was continued.
Quantitative PCR (qPCR)
RNA was extracted and its concentration and purity
were determined using an ultra microspectrophotometer (Thermo
Fisher Scientific, Inc., Waltham, MA, USA). Following reverse
transcription, the synthesized cDNA samples were subjected to qPCR
(Applied Biosystems, Foster City, CA, USA) using the following
primer sequences: NT3, forward 5′-ACC AGGTTA CAG TGT TGG GAG AG-3′
and reverse 5′-CGC TGA GAG TTC CAG TGT TTG TC-3′; rat GAPDH,
forward 5′-CCT TCC GTG TTC CTA CCC-3′ and reverse 5′-CAA CCT GGT
CCT CAG TGT AG-3′. The PCR reaction conditions were as follows:
predenaturation, 95°C for 60 sec; PCR cycles (×40) consisting of
95°C for 15 sec and then 60°C for 1 min; melting curve analysis at
95°C for 15 sec (100%), 70°C for 1 min (1%) and at 95°C for 30
sec.
Western blotting
Radioimmunoprecipitation assay lysate (Beyotime
Institute of Biotechnology, Shanghai, China) was used to extract
the total cellular proteins. A nucleic acid protein analyzer
(Thermo Fisher Scientific, Inc.) was then used to determine the
total concentrations of the sample proteins. Following
SDS-polyacrylamide gel electrophoresis, the proteins were
transferred and hybridized. The membrane was incubated overnight
with the appropriately diluted primary antibodies (rabbit anti-rat
NT-3 polyclonal antibody; Sigma-Aldrich, St. Louis, MO, USA) at
4°C. Subsequently, the membrane was incubated with the
appropriately diluted secondary antibodies (Abcam, Cambridge, MA,
USA) at room temperature for 1 h and stained using an enhanced
chemiluminescence kit (Thermo Fisher Scientific, Inc.). The
membrane was analyzed using a Fiuorchem HD2 gel-imaging analysis
system (ProteinSimple Inc., Santa Clara, CA, USA), and the
development and fixation of specific protein bands were
detected.
Observation indicators
Electrophoretic identification and sequencing
identification of the recombinant plasmid
PLV.Ex3d.P/neo-EF1A>NT3>IRES/EGFP was performed. Following
the lentiviral packaging, the expression of GFP and the positive
transfection efficiency were determined. The mRNA and protein
expression in the PLV.Ex3d.P/neo-EF1A/NT3/IRES/EGFP-transfected
BMSCs was determined.
Results
Electrophoresis of the PCR products of
the SD rat recombinant NT-3 gene
PCR amplification yielded the ~841 bp NT-3 gene
fragment attB1-NT3-attB2 (Fig.
1A). Gateway clone technology was used to generate the entry
clone pDown-NT3 (~1041 bp; Fig.
1B) via the BP reaction and to produce the recombinant target
plasmid PLV.Ex3d.P-neo-EF1A/NT3/-IRES/EGFP (~830 bp; Fig. 1C). The colony was screened for
positive clones. Sequencing of the recombinant target plasmid
demonstrated that the expected sequence was achieved. This result
indicated the successful construction of the recombinant target
plasmid PLV.Ex3d.P-neo-EF1A-NT3-IRES-EGFP.
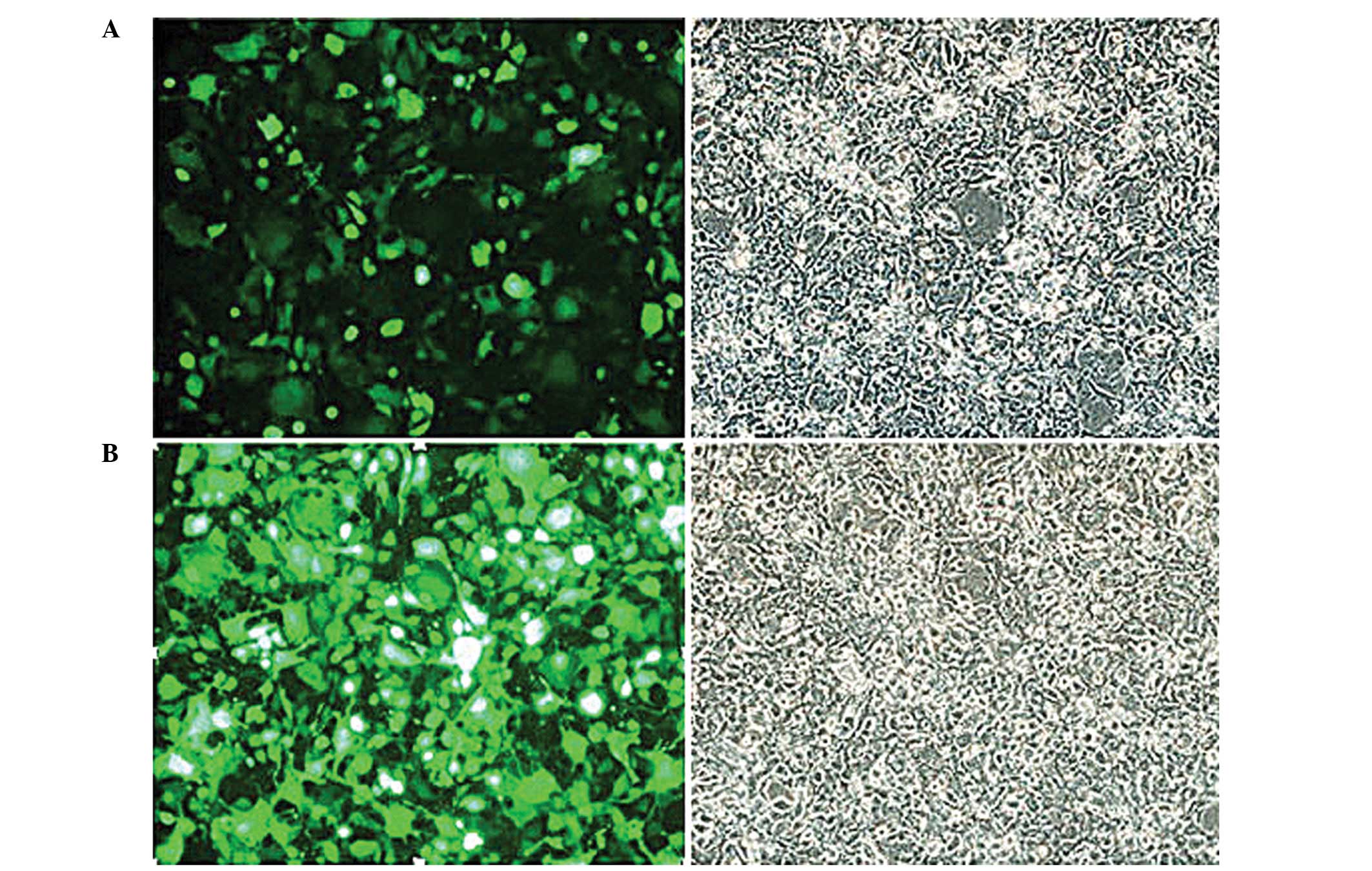
Results of the lentiviral packaging
A lentiviral packaging system was used to pack the
293FT cells in the logarithmic growth phase, as shown in the
fluorescence image (Fig. 2A). The
fluorescence image of pLV.Ex2d.P/neo-EF1A/EGFP in the control group
(Fig. 2B) shows the generation of
EGFP fluorescence proteins. This result indicated the successful
packaging of the lentivirus.
Determination of the fluorescence
expression of the lentivirus titer
The number of fluorescent cells decreased with
increasing dilution. At a stock lentivirus solution of
10−5 μl, the GFP remained expressed. Thus, the
preliminary lentivirus titer was determined as 3.5×10−8
TU/ml.
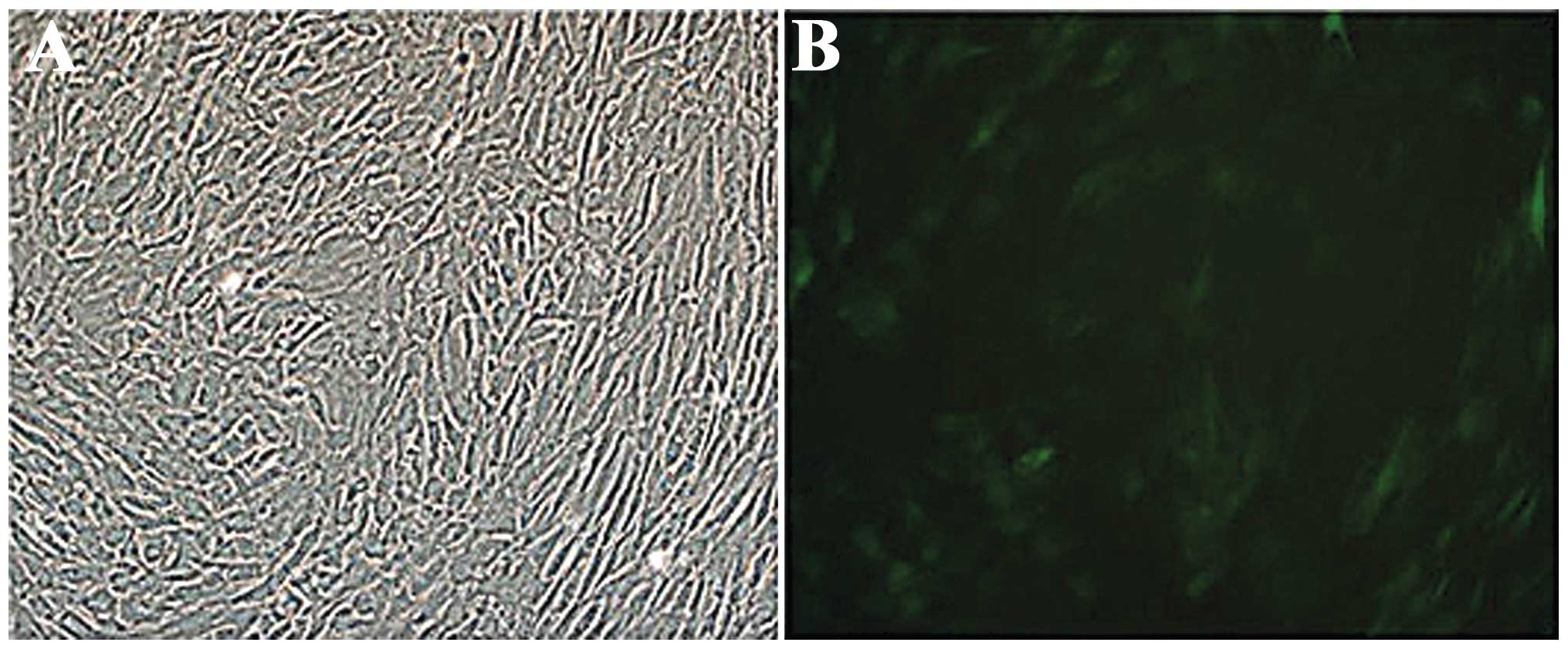
Establishment of stably transfected SD
BMSCs
The PLV Ex3d.P-neo-EF1A-NT3-IRES-EGFP-transfected
BMSCs were purified via drug screening and observed under an
inverted fluorescence microscope. The results demonstrated the
growth of a positive cell colony. Following the transfection, no
significant change was identified in the BMSC morphology and the
cells uniformly expressed GFP. The transfection efficiency was
60–70% (Fig. 3).
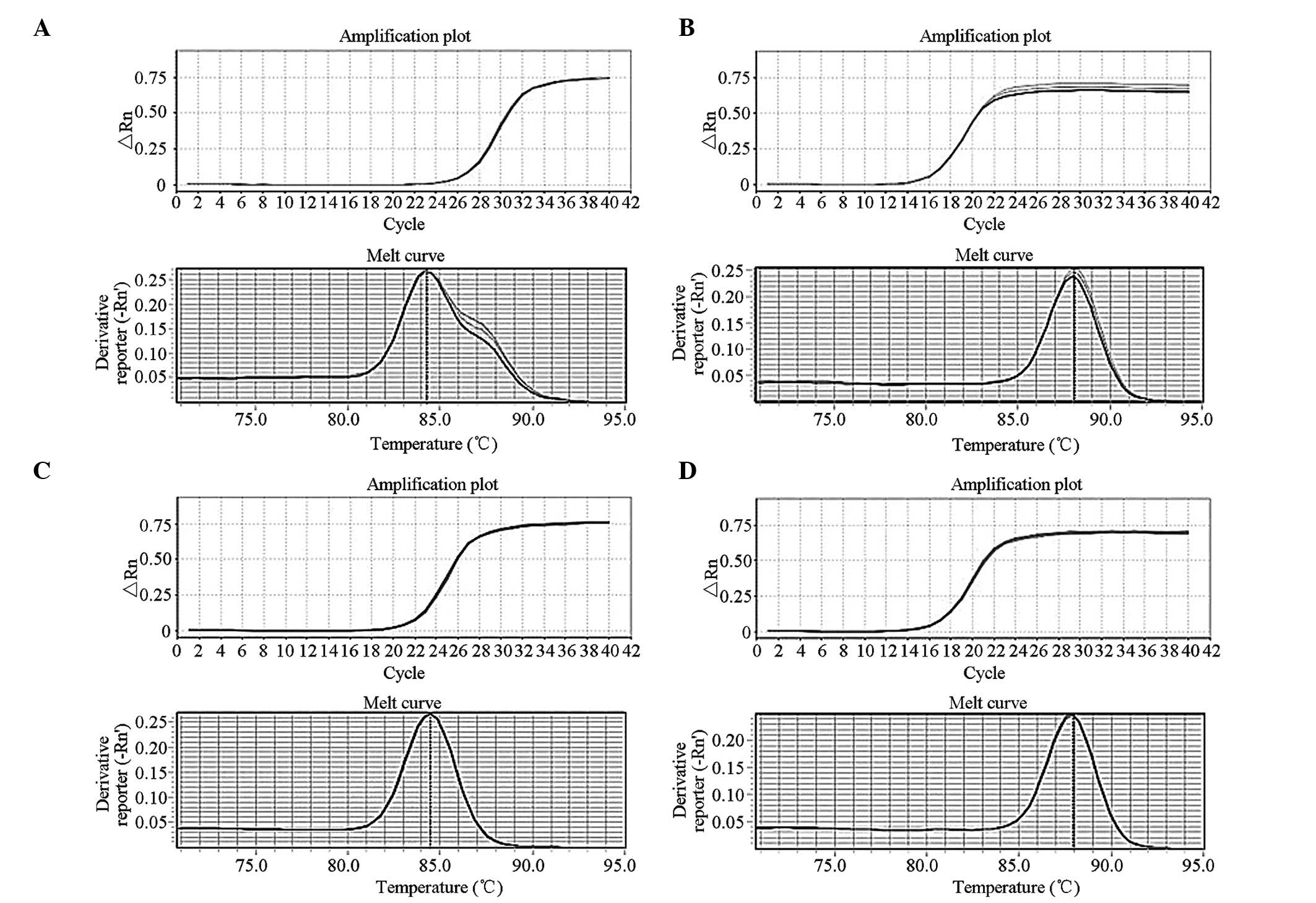
Determination of NT-3 expression
Western blotting and qPCR were mainly used to verify
the transcription and translation of NT-3 in the transfected cells,
particularly in terms of nucleic acid and protein levels. The mRNA
levels in the NT-3-transfected BMSCs significantly increased
(Fig. 4) compared with those of
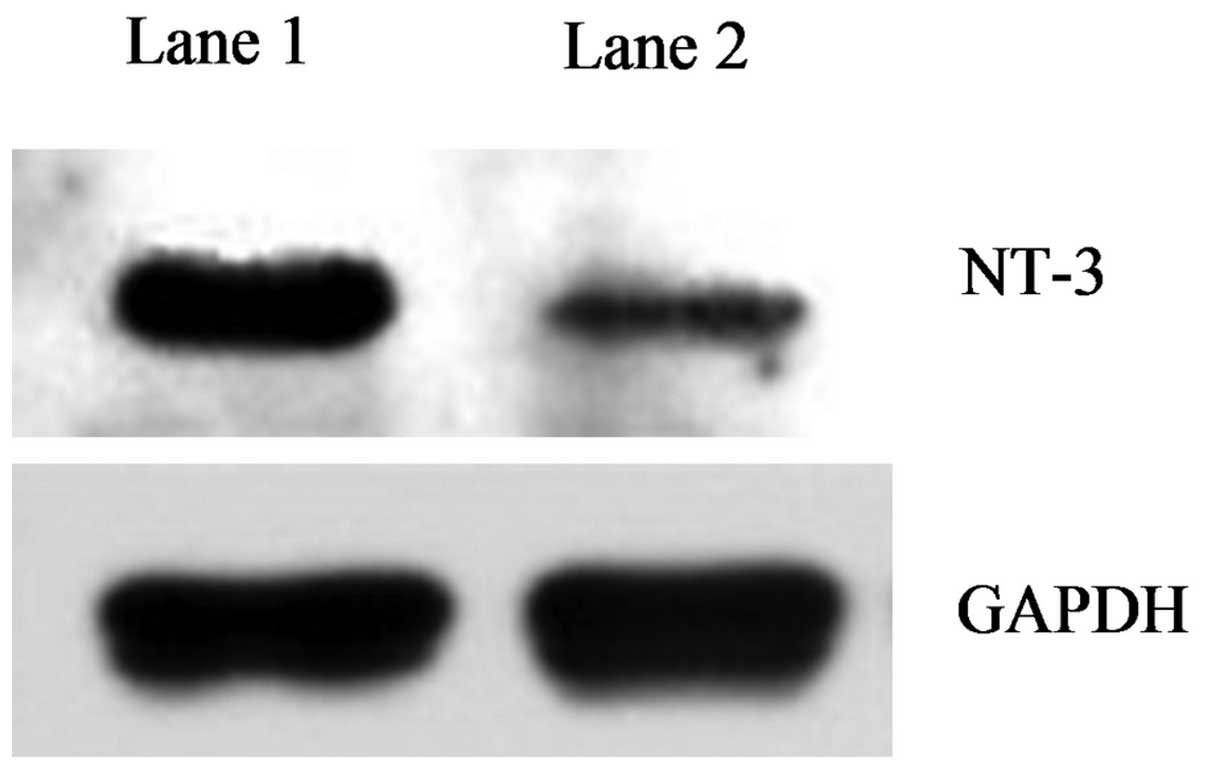
the untransfected group. Similar results were obtained from the
western blot analysis. The NT-3 protein band in the untransfected
BMSC group was significantly weaker than that in the transfected
group (Fig. 5). These results
indicated that the NT-3 protein expression in the NT-3-transfected
BMSC group was significantly higher than that in the untransfected
group.
The findings of the present study demonstrated that
the Gateway clone technique successfully constructed the
recombinant target plasmid PLV.Ex3d.P-neo-EF1A-NT3-IRES-EGFP and
efficiently transfected it into the SD rat BMSCs, which maintained
a high NT-3 expression for a long period.
Discussion
In SCI repair, deficiency in neurotrophic factors in
the damaged local microenvironment is generally considered an
important factor that determines functional recovery following SCI
(9). Therefore, increasing the
concentration of neurotrophic factors in the microenvironment may
promote and induce the regeneration of nerve cells and is a
potential therapeutic treatment strategy for SCIs. Zhou et
al (10) confirmed that NT-3
has a significant role in inhibiting neuronal apoptosis and
promoting axonal regeneration during SCI repair and is considered
the most effective of the neurotrophic factors. However, the
topical application of NT-3 in SCIs is clinically limited due to
its short half-life, immunogenicity, difficulty in passing through
the blood/spinal barrier and ineffective promotion in spinal
pathway reconstruction following syringomyelia. To overcome these
disadvantages, molecular biological methods were designed to
transfer the therapeutic NT-3 gene into a suitable cell vector and
allow the gene to maintain a stable expression in the cell. The
NT-3 gene-carrying cell vector was then replanted in the SCIs to
achieve sustained secretion and thus promote the recovery of
neurological functions following SCI. To a certain extent, these
methods overcome the limitations of NT-3 in SCI treatment.
Neurotrophic factors and their receptors are widely
present in the spinal cord. However, with the exception of the high
NT-3 levels during early spinal cord development (11), the levels of other neutrophic
factors are relatively low in the spinal cord. Among the receptor
tyrosine kinases, tyrosine kinase receptor C has a marked affinity
for NT-3 (12). As an important
member of neurotrophic factor families, NT-3 is involved in the
development and differentiation of neurons and can promote the
survival of damaged central neurons as well as the regeneration of
nerve fibers (13). In addition,
NT-3 promotes the proliferation of oligodendrocytes and the
myelination of regenerated axons and serves as a guide for damaged
axons (14).
BMSCs are fibroblast-colony units derived from the
bone marrow. These cells exhibit a high degree of self-renewal
capacity, marked proliferative capacity, high plasticity and
potential pluripotency (15).
Furthermore, BMSCs self-originate and thus have low immunogenicity,
are easily sourced and can stably express a variety of exogenous
target genes in vivo and in vitro (16). Due to these features, BMSCs have
been used in the cell therapy of various diseases and as vector
cells in gene therapy (17)
Brazelton et al (18) and
Mezey et al (19) found
that in vivo, exogenous BMSCs differentiate into glial cells
and neurons via different routes following migration into the mouse
brain. This finding suggests the feasibility of using transplanted
BMSCs to treat central nervous system damage. Hofstetter et
al (20) transplanted BMSCs to
treat a standard spinal cord injury in a rat model and demonstrated
that BMSCs can promote the recovery of hind limb functions. Several
studies attributed this neuroprotective effect of BMSCs to their
ability to differentiate into neurons, which replace diseased or
damaged neurons (21). However,
Mahmood et al (22)
attributed the ability of transplanted BMSCs to promote functional
recovery in SCIs to the secretion of neurotrophic factors by these
cells within the damaged tissues. These neurotrophic factors in
turn promote neuronal survival and induce the proliferation of
endogenous cells and the regeneration of nerve fibers. Further
studies demonstrated that BMSCs have high biocompatibility as well
as excellent immunoregulation and immune circumvention abilities.
Therefore, BMSCs have become the dominant vectors of neurotrophin
cells (23). In the quantitative
analysis of neurotrophic factors secreted by BMSCs, Crigler et
al (24) detected the
expression of neurotrophic factors, including brain-derived
neurotrophic factor and nerve growth factor but did not detect NT-3
expression.
In the present study, the lentivirus-mediated method
was used to transfect the NT-3 gene into SD rat BMSCs in order to
establish NT-3-carrying BMSC strains. Gateway clone technology was
used in the recombination and construction of the target plasmid.
This technique is a new universal vector construction method based
on the phage λ site-specific recombination system (25), which can rapidly clone heterologous
DNA fragments into different vectors with high specificity. The
recombination sites include attB, attP, attL and attR. The
recombination reactions include the BP recombination reaction, in
which attP-containing donor vectors react with attB-containing PCR
products, sequence-exchange reactions occur at the att sites and
entry clones are generated and the LR recombination reaction, in
which attL-containing entry clones react with attR-containing
target vector, a sequence-exchange reaction occurs at the att sites
and eukaryotic expression vectors of the target gene are generated.
Compared with traditional vector construction methods, including
restricted enzyme digestion and ligation, Gateway cloning is easy
and highly efficient. Furthermore, this method can significantly
reduce false positive results during vector construction (26), keep the developing and reading
frames unchanged and achieve fast and efficient transfers of DNA
fragments (27). In the present
study, the overlapping PCR amplification method was used to obtain
attB1-NT-3-attB2. Gateway cloning then successfully constructed the
recombinant target plasmid PLV.Ex3d.P-neo-EF1A-NT3-IRES-EGFP for
the viral packaging. Although adenoviral vectors have high
transfection rates, their corresponding immune response poses
certain risks in in vivo experiments (28). Therefore, a lentivirus was used as
the expression vector to prevent unwanted side effects. A
third-generation lentiviral packaging system was used to pack the
293FT cells into the viral particles, which were then transfected
with Lipofectamine 2000. The fluorescence image demonstrated that
EGFP was generated at a transfection rate >90%. The lentiviral
vector, which completed the viral packaging and carried NT-3/EGFP,
was transfected into the SD BMSCs. NT-3/EGFP-carrying SD BMSCs
strains were obtained following drug screening and purification.
qPCR and western blotting confirmed that the NT-3 gene transfected
into SD BMSCs maintained a stable expression. The recombinant
lentiviral vector significantly improved the gene transfection
efficiency of the BMSCs and the probability of target gene
integration into the host cell genome was significantly increased.
The efficient integration of a foreign gene into the host
chromosome resulted in a stable and lasting expression (29). The BMSCs stably and efficiently
expressed the foreign genes and thus overcame the disadvantages of
topical cytokine application.
In conclusion, the present study used a recombinant
lentiviral vector as the expression vector to transfect the target
gene PLV Ex3d.P/neo-EF1A-NT3-IRES-EGFP into the SD rat BMSC genome.
The transfected BMSCs successfully expressed NT-3 and EGFP. In
addition, the transfected BMSCs exhibited a stable NT-3 secretion
and the EGFP gene successfully traced and located the site of
target gene expression in vivo and in vitro. The
results of the present study provide a solid foundation for further
experimental studies on NT-3 gene-modified BMSC transplantation for
SCI treatment. The results may also serve as a reference for the
development of reliable NT-3 gene-modified BMSCs.
Acknowledgements
This study was supported by the Liaoning Provincial
Science and Technology Project Foundation (grant no.
2011225013).
References
|
1
|
Sahani V and Kessler JA: Stem cell
therapies for spinal cord injury. Nat Rev Neurol. 6:363–372. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sensebe L, Krampera M, Schrezenmeier H,
Bourin P and Giordano R: Mesenchymal stem cell for clinical
application. Vox Sang. 98:93–107. 2010. View Article : Google Scholar
|
|
3
|
Uccelli A, Moretta L and Pistoia V:
Mesenchyal stem cells in health and disease. Nat Rev Immunol.
8:726–736. 2008. View
Article : Google Scholar
|
|
4
|
Sanchez-Ramos J, Song S, Cardozo-Pelaez F,
et al: Adult bone marrow stromal cells differentiate into neural
cells in vitro. Exp Neurol. 164:247–256. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lu P, Jones LL and Tuszynski MH:
BDNF-expressing marrow stromal cells support extensive axonal
growth at sites of spinal cord injury. Exp Neurol. 191:344–360.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ohta M, Suzuki Y, Noda T, et al: Bone
marrow stromal cells infused into the cerebrospinal fluid promote
functional recovery of the injured rat spinal cord with reduced
cavity formation. Exp Neurol. 187:266–278. 2004. View Article : Google Scholar
|
|
7
|
Tuszynski MH, Grill R, Leonard LL, et al:
NT-3 gene delivery elicits growth of chronically injured
corticospinal axons and modestly improves functional deficits after
chronic scar resection. Exp Neurol. 181:47–56. 2003. View Article : Google Scholar
|
|
8
|
Ejstrup R, Kiilgaard JF, Tucker BA,
Klassen HJ, Young MJ and La Cour M: Pharmacokinetics of
intravitreal glial cell line-derived neurotrophic factor:
experimental studies in pigs. Exp Eye Res. 91:890–895. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Fawectt J: Repair of spinal cord injuries:
where are we, where are we going? Spinal Cord. 40:615–623. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhou L, Baumgartner BJ, Hill-Felberg SJ,
McGowen LR and Shine HD: Neurotrophin-3 expressed in situ induces
axonal plasticity in the adult injured spinal cord. J Neurosci.
23:1424–1431. 2003.PubMed/NCBI
|
|
11
|
Zhang W, Li Y, Wang ZJ, Zhou X, Ou KQ,
Zhou HL and Wang TH: Functional roles of intrinsic neurotrophin-3
in spinal neuroplasticity of cats following partial ganglionectomy.
Growth Factors. 28:351–358. 2010. View Article : Google Scholar
|
|
12
|
Ba YC, Dai P, Zhou HL, Liu J and Wang TH:
Spatiotemporal changes of NGF, BDNF and NT-3 in the developing
spinal cords of embryonic chicken. Neurochem Res. 35:273–278. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chapleau CA and Pozzo-Miller L: Divergent
roles of p75NTR and Trk receptors in BDNF’s effects on dendritic
spine density and morphology. Neural Plast.
2012:5780572012.PubMed/NCBI
|
|
14
|
Xian HQ, McNichols E, St Clair A and
Gottlieb DI: A subset of ES-cell-derived neural cells marked by
gene targeting. Stem Cells. 21:41–49. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang XJ and Li QP: The roles of
mesenchymal stem cells (MSCs) therapy in ischemic heart diseases.
Biochem Biophys Res Commun. 359:189–193. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lu L, Zhao C, Liu Y, et al: Therapeutic
benefit of TH-engineered mesenchymal stem cells for Parkinson’s
disease. Brain Res Brain Res Protoc. 15:46–51. 2005.PubMed/NCBI
|
|
17
|
Kan EM, Ling EA and Lu J: Stem cell
therapy for spinal cord injury. Curr Med Chem. 17:4492–4510. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Brazelton TR, Rossi FM, Keshet GI and Blau
HM: From marrow to brain: expression of neuronal phenotype in adult
mice. Science. 290:1775–1779. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Mezey E, Chandross KJ, Harta G, Maki RA
and McKercher SR: Turning blood into brain: cells bearing neuronal
antigens generated in vivo from bone marrow. Science.
290:1779–1782. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hofstetter CP, Schwarz EJ, Hess D,
Widenfalk J, El Manira A, Prockop DJ and Olson L: Marrow stromal
cells form guiding strands in the injured spinal cord and promote
recovery. Proc Natl Acad Sci USA. 99:2199–2204. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Black IB and Woodbury D: Adult rat and
human bone marrow stromal stem cells differentiate into neurons.
Blood Cells Mol Dis. 27:632–636. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Mahmood A, Lu D and Chopp M: Marrow stroma
cell transplantation after traumatic brain injury promotes cllular
proliferation within the brain. Neurosurgery. 55:1185–1193. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang Y, Liu J, Xu C, et al: Bone marrow
transplantation combined with mesenchymal stem cells induces immune
tolerance without cytotoxic conditioning. J Surg Res.
171:e123–e131. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Crigler L, Robey RC, Asawachaicham A,
Gaupp D and Phinney DG: Human mesenchymal stem cell subpopulations
express a variety of neuro-regulatory molecules and promote
neuronal cell survival and neuritogenesis. Exp Neurol. 198:54–64.
2006. View Article : Google Scholar
|
|
25
|
Landy A: Dynamic, structural, and
regulatory aspects of lambda site-specific recombination. Annu Rev
Biochem. 58:913–949. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Bernard P and Couturier M: Cell killing by
the F plasmid CcdB protein involves poisoning of DNA-topoisomerase
II complexes. J Mol Biol. 226:735–745. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Magnani E, Bartling L and Hake S: From
Gateway to MultiSite Gateway in one recombination event. BMC Mol
Biol. 7:462006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Su JG, Zhu ZY, Wang Y, Xiong F and Zou J:
The cytomegalovirus promoter-driven short hairpin RNA constructs
mediate effective RNA interference in zebrafish in vivo. Mar
Biotechnol. 10:262–269. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Fan JH and Li YJ: Cloning and sequence
analysis of the M gene of porcine epidemic diarrhea virus LJB/03.
Virus Genes. 30:69–73. 2005. View Article : Google Scholar : PubMed/NCBI
|