Introduction
Hypoxic-ischemic encephalopathy (HIE) is the primary
cause of cerebral damage and long-term neurological sequelae,
including cerebral palsy and mental retardation (1), in the perinatal period in term and
preterm infants (2). However, no
specific and successful neuroprotective strategies currently exist.
Brain protection in newborn infants remains a challenging priority
and represents a medical requirement that has yet to be met.
Recently, microRNAs (miRNAs), a large conserved
family of noncoding short RNAs, have been found to be novel
therapeutic tools for the treatment of ischemic hypoxia (IH).
Hypoxia and insufficient blood flow to the brain, known as brain
ischemia, lead to poor oxygen supply and consequently the death of
brain tissue, with a wide range of pathophysiological outcomes.
miRNAs have previously been reported to be induced by oxygen
deprivation. In particular, microRNA-210 (miR-210), which is
activated by the transcriptional factor hypoxia inducible factor-1α
(HIF-1α) (3), is a unique miR that
is evolutionarily conserved and ubiquitously expressed in a number
of hypoxic cell and tissue types (4–10).
HIF-1α is an important factor in the molecular mechanisms of oxygen
deprivation (11–16) and the development of injured and
normal brains. Hence, we hypothesized that miR-210 may be involved
in HIE, even though its exact role remains to be determined. Its
association with HIE may establish miR-210 as a potential
therapeutic target.
During normal brain development, redundant neurons
are removed via apoptosis; this is an important physiological
process that ensures the formation of appropriate neural networks.
However, following IH, this apoptotic component becomes
detrimental, leading to excessive neuronal loss. Therefore, in a
previous study (17), the effect
of miR-210 on the IH-induced apoptosis of neurons was investigated
in an ex vivo model of IH (18–20),
through the use of PC12 rat pheochromocytoma cells combined with
oxygen glucose deprivation (OGD). The results of the study
demonstrated that miR-210 overexpression suppressed cell apoptosis
by inhibiting caspase activity and regulating the balance between
the expression levels of Bcl-2 and Bax. The aim of the present
study was to further elucidate the effect of miR-210 on apoptosis
in PC12 cells via the silencing of its expression.
Materials and methods
Cell culture
PC12 rat pheochromocytoma cells were obtained from
the American Tissue Culture Collection (Rockville, MD, USA) and
cultured in Dulbecco’s modified Eagle’s medium (DMEM) supplemented
with 10% v/v horse serum (HS), 5% v/v fetal bovine serum (FBS) and
appropriate antibiotics in a humidified chamber (5% CO2
and 37°C), all of which were purchased from Invitrogen Life
Technologies (Carlsbad, CA, USA).
miRNA transfection
An hsa-miR-210 inhibitor (anti-miR210) or
anti-miR-negative control #1 (Ambion, Austin, TX, USA) (50 μl) was
diluted in OptiMEM I (Invitrogen, Carlsbad, CA, USA) to a
concentration of 100 nM. Subsequently, 50 μl of the solution was
mixed with 50 μl Lipofectamine 2000 (Invitrogen) in OptiMEM I (25X
dilution), incubated at room temperature for 20 min and then added
to each well of a 24-well plate (100 μl). PC12 cells (400 μl,
concentration 6.25×105 cells/ml) were added to each
well. The transfection mixture was incubated (5% CO2 and
37°C) for 24 h and the cells were either used immediately in assays
or the media was replaced (500 μl DMEM/10% v/v HS/5% v/v FBS) and
the mixture incubated further.
Oxygen glucose deprivation (OGD)
A mixture of 95% nitrogen and 5% CO2 was
passed through glucose-free DMEM to deoxygenate it. The DMEM was
used to wash the PC12 cells once and then to maintain them prior to
use. The cells were then placed in a modular incubation chamber
(Billups-Rothenberg, Del Mar, CA, USA) and flushed with 95%
nitrogen/5% CO2 for 4 min at a flow rate of 10 l/min.
The chamber was sealed and stored in an incubator for 4 h at 37°C.
Control cells were washed with glucose-containing DMEM and
incubated in a normoxic incubator for 4 h, as previously described
(17).
miRNA reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was prepared using TRIzol®
reagent (Invitrogen). miRNA was purified using the mirVana™ kit
(Applied Biosystems, Foster City, CA, USA), according to the
manufacturer’s instructions. Using specific miR-210 and an
endogenous control U6 stem-loop primer (Applied Biosystems, Foster
City, CA, USA), reverse transcription was performed using the
TaqMan® MicroRNA Reverse Transcription kit (Applied
Biosystems) according to the manufacturer’s instructions. Total RNA
(10 ng) was reverse transcribed to cDNA using the following
reagents: 1 mM dNTPs (with dTTP), 1 μl reverse transcriptase (50
U/μl) and RNase inhibitor (4 U) in the presence of specific miR-210
or U6 stem-loop reverse transcriptase primers in a 15-μl system
buffered with RT buffer and diethylpyrocarbonate (DEPC) water. The
RT thermal cycle program was as follows: 16°C for 30 min, 42°C for
30 min and 85°C for 5 min. The resulting cDNA was stored at −20°C.
The qPCR step was performed using a 7900HT Fast Real-Time PCR
system with a TaqMan® MicroRNA Assay kit (Applied
Biosystems). The total reaction volume was 20 μl and contained the
following components: miR-210 or U6 RT reaction product (1.33 μl),
1 μl 20X TaqMan® MicroRNA assay (for miR-210 or U6), 10
μl 2X TaqMan Universal PCR Master mix and 7.67 μl DEPC water
(Sigma-Aldrich, St. Louis, MO, USA). qPCR was then performed on the
96-well plate using the following protocol: 95°C for 10 min,
followed by 43 cycles of 95°C for 15 sec and 60°C for 1 min. The
relative miR-210 level was normalized to the expression of the
endogenous control U6 for each sample, calculated using the
2−ΔΔCt method. Experiments were performed in
triplicate.
Evaluation of apoptotic index
Cells were harvested using
trypsin/ethylenediaminetetraacetic acid (EDTA) (Sigma-Aldrich),
washed with phosphate-buffered saline (PBS; Sigma-Aldrich),
resuspended in 100 μl binding buffer (BioVision, Milpitas, CA, USA)
and stained with 5 μl Annexin V-fluorescein isothiocyanate (FITC)
and 1 μl propidium iodide (PI) at room temperature for 15 min
(Biovision). The fluorescence of FITC and PI was analyzed using
flow cytometry (Becton Dickinson, Mountain View, CA) following the
addition of 400 μl binding buffer.
Western blot analysis
Cells were washed with ice-cold PBS and lysed in
protein lysis buffer (50 mmol/l Tris, 150 mmol/l NaCl, 10 mmol/l
EDTA, 1% Triton X-100, 200 mmol/l NaF and 4 mmol/l sodium
orthovanadate-containing protease inhibitors, pH 7.5) for 1 h on
ice. Proteins were quantified using a bicinchoninic acid (BCA)
Protein Assay kit (Pierce, Rockford, IL, USA) according to the
manufacturer’s instructions. Following separation by 10% sodium
dodecyl sulfate-polyacrylamide gel electrophoresis, the proteins
(20 μg/lane) were electrophoretically transferred onto a
nitrocellulose membrane (Whatman, London, UK), which was blocked
with non-fat dry milk in buffer. The membrane was incubated with
mouse monoclonal antibodies against caspase-3, caspase-9, Bax and
Bcl-2 (Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA) and
goat anti-mouse IgG secondary antibodies conjugated with
horseradish peroxidase (Santa Cruz Biotechnology, Inc.) according
to the manufacturer’s instructions. Thereafter the proteins were
visualized with an electrochemiluminescence detection system (GE
Healthcare Bio-Sciences, Uppsala, Sweden) and analyzed using
Quantity One Analysis software (Bio-Rad Laboratories, Hercules, CA,
USA). β-actin (Cell Signaling Technology, Inc., Beverly, MA, USA)
was used as protein loading control.
Statistical analysis
All data are expressed as the mean ± the standard
error of the mean. Statistical analysis was performed using the
paired Student’s t-test of the SPSS statistical software package
v10.0 (SPSS, Inc., Chicago, IL, USA). P<0.05 was considered to
indicate a statistically significant difference.
Results
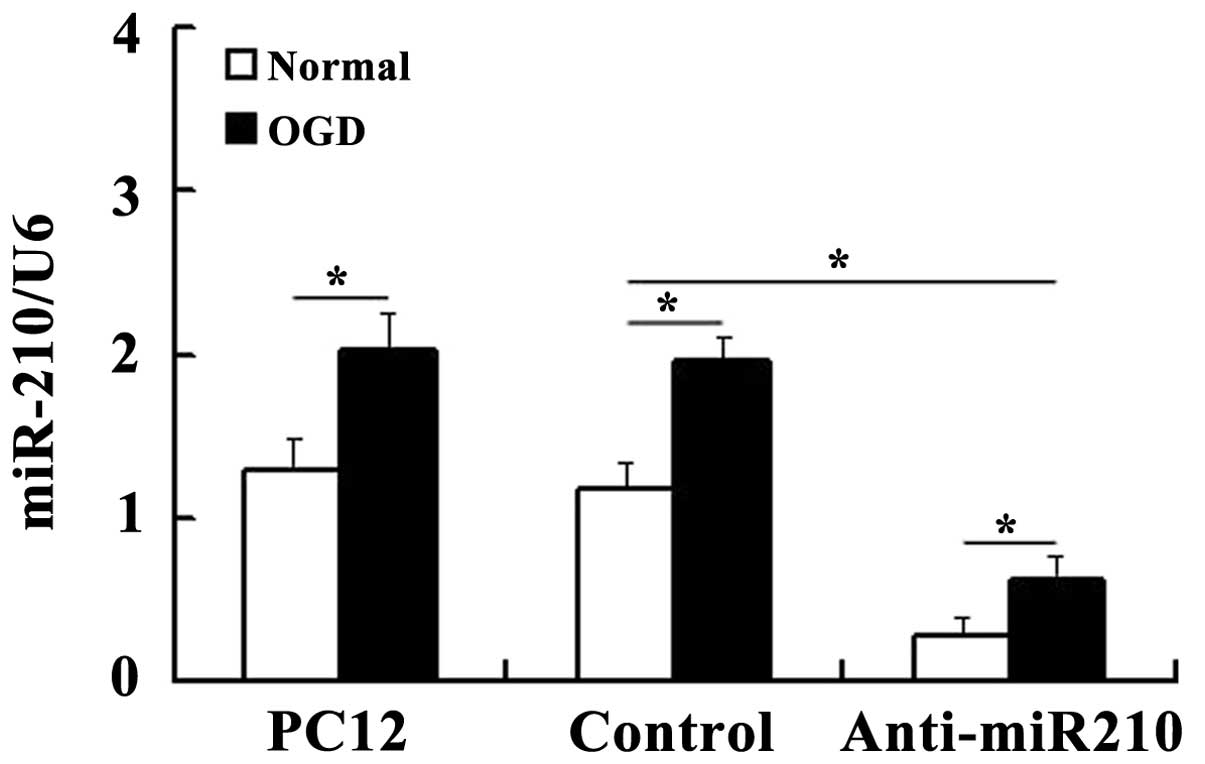
OGD significantly increases miR-210
expression levels
The expression levels of miR-210 were measured using
RT-qPCR analysis. U6 was used as the endogenous control as it was
the most stably expressed miR across all subjects in the control
and experimental groups (data not shown). Expression of miR-210 was
markedly downregulated in cells transfected with anti-miR210
compared with that in the U6 controls, which confirmed that the
miR-210 knockdown cells had been prepared successfully. Consistent
with previous results (17),
RT-qPCR revealed that miR-210 expression was significantly induced
in PC12 cells under hypoxic conditions (Fig. 1).
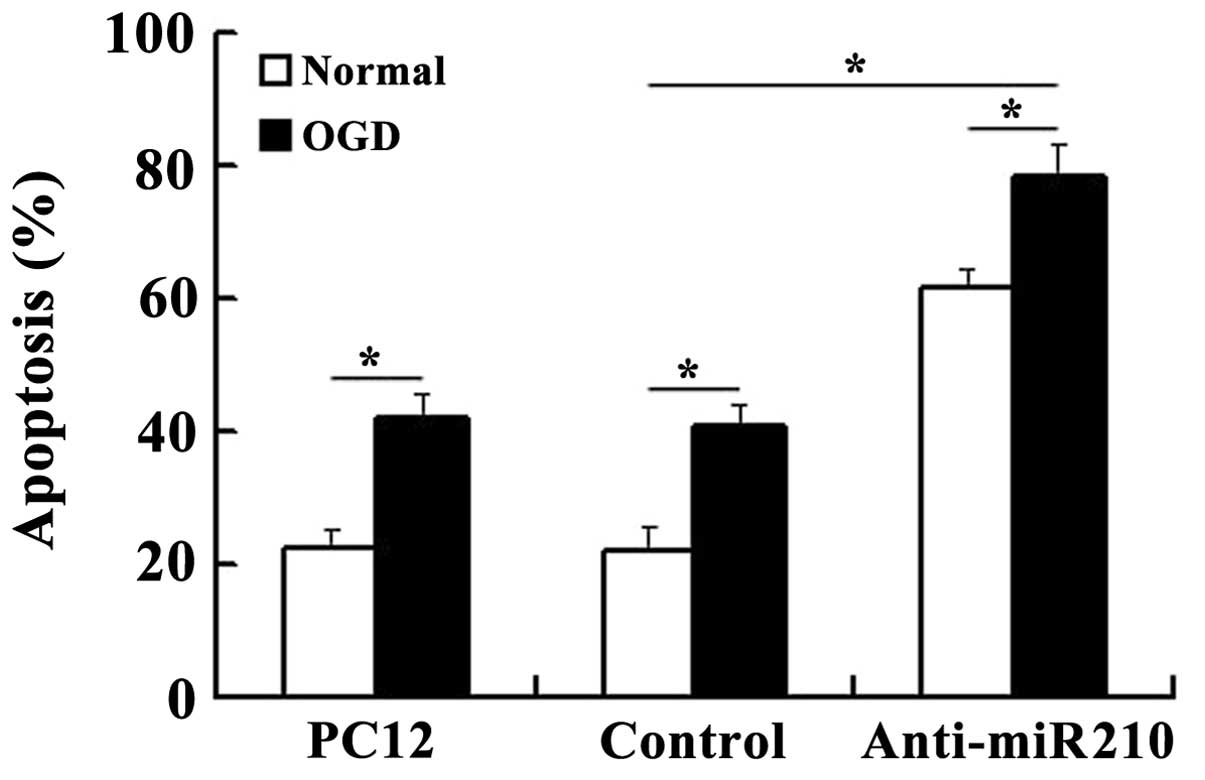
miR-210 knockdown increases the levels of
cell apoptosis
As shown in Fig. 2,
compared with the control cells that were not deprived of glucose
and maintained under normoxic conditions, the cells subjected to
OGD showed a higher level of cell death. However, miR-210 knockdown
increased the levels of apoptosis observed in cells, particularly
following subjection to OGD.
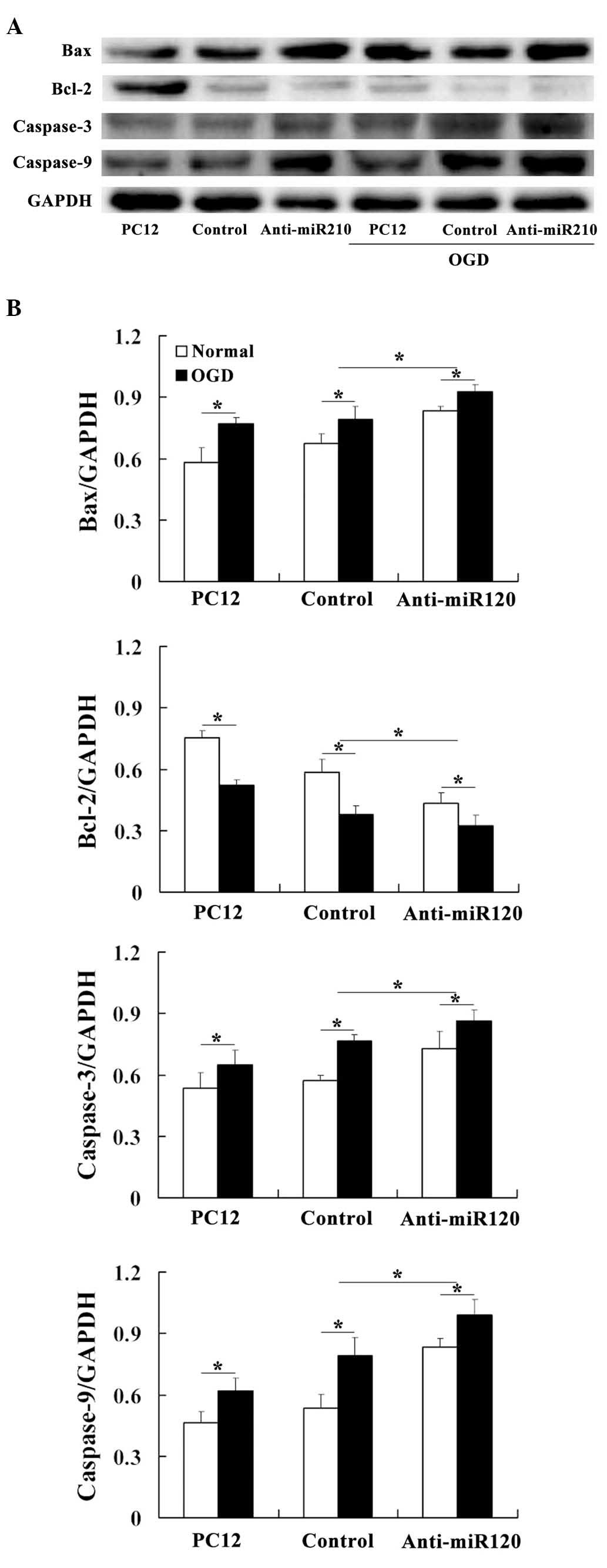
miR-210 alters the protein expression
levels of caspase-3, caspase-9, Bax and Bcl-2
The effects of miR-210 on the expression levels of
apoptosis-related proteins were assessed. Western blot analysis
demonstrated that the protein levels of Bax, caspase-3 and
caspase-9 were increased in the miR-210 knockdown cells compared
with those observed in the controls. By contrast, anti-apoptotic
Bcl-2 expression levels changed in an almost inverse manner
(Fig. 3).
Discussion
miRNAs are small noncoding RNAs that silence the
expression of target genes through either mRNA degradation or the
suppression of transcription procedures. While several studies have
demonstrated the roles of specific miRNAs in neuronal
differentiation, neurogenesis, neural cell specification and
neurodevelopmental function (21–23),
no studies are available on the importance of miRNAs in HIE to the
best of our knowledge. Recently, a specific group of
hypoxia-regulated miRNAs that are under the regulation of HIF-1α
were identified. miR-210, a direct transcriptional target of
HIF-1α, is one of the most hypoxia-sensitive miRNAs and has a
number of pleiotropic effects. miR-210 has been mechanistically
linked to the control of a wide range of cellular responses known
to influence normal developmental physiology, as well as a number
of hypoxia-dependent disease states, including tissue ischemia,
inflammation and tumorigenesis.
A previous study demonstrated that miR-210 has
neuroprotective effects through inhibiting apoptosis in a murine
model of HIE (24). Furthermore,
an OGD model was used to investigate the link between miR-210 and
IH injury in PC12 cells through the overexpression of miR-210
(17). To further elucidate the
effect of miR-210 on apoptosis in PC12 cells following exposure to
OGD, the present study used anti-miRNA transfection to downregulate
the expression of miR-210.
A previous study has shown that miR-210 expression
is upregulated in PC12 cells following a 4-h exposure to OGD
(17). Consistently, the current
study also demonstrated that OGD upregulates the expression of
miR-210 in both miR-210 knockdown and control cells. The induction
of miR-210 by hypoxia is consistent with studies involving other
types of cell, including embryo kidney cells, endothelial cells,
breast carcinoma cells, colonic adenocarcinoma cells and ovarian
epithelial cells (8,25,26).
Thus far, in all published studies, miR-210 is an miRNA that has
been consistently observed to be upregulated in normal and
transformed hypoxic cells to the best of our knowledge.
miRNAs are involved in a diverse range of biological
processes, including development, cell growth, apoptosis and
hematopoiesis. During normal brain development, redundant neurons
are removed via apoptosis; this is an important physiologic process
to ensure the formation of appropriate neural networks. However,
following IH, this apoptotic component becomes detrimental, leading
to excessive neuronal loss. Previous studies have revealed that
miR-210 treatment within the first 4 h post-OGD prevents cell
apoptosis (17). The present study
showed that anti-miR210 transfection induced apoptosis in both
normoxic and hypoxic conditions, which demonstrates that miR-210
modulates the cellular apoptotic response to OGD. In support of
this finding, the results revealed that miR-210 inhibition
upregulates expression levels of Bax, caspase-3 and caspase-9;
while downregulating Bcl-2 expression. Thus, it may be hypothesized
that the miR-210 inhibition-dependent upregulation of Bax,
caspase-3 and caspase-9 expression and downregulation of Bcl-2
expression may contribute to the modulation of the apoptotic
response to IH.
miRNA-based therapeutics are set to become the one
of the most significant commercial hotspots in the health care
market (27–31). Although there are a number of
challenges for miRNAs as therapeutic targets, including delivery,
potential off-target effects and safety, the strategy of
manipulating miRNAs in vivo to regulate disease-related
processes is already becoming a feasible therapeutic approach for
the future (32), due to the small
size of miRNAs and their high rate of transduction in eukaryotic
cells. Furthermore, the feasibility of miRNAs as therapeutics is
supported by ongoing clinical trials (33). In the present study, we hypothesize
that miR-210 may have a significant role in regulating apoptosis
following IH injury and may lead to a novel therapy for the
treatment of HIE.
In conclusion, the present study demonstrated that
the knockdown of miR-210, which is a highly upregulated miRNA
induced by OGD, contributes to OGD-induced neuronal apoptosis
through the induction of caspase activity and the regulation of the
balance between Bcl-2 and Bax expression levels. Further
investigation of the regulation, targets and physiological and/or
pathogenic effects of miR-210 in IH should be anticipated in the
future.
Acknowledgements
This study was supported by grants from the Science
and Technology Development Project of Nanjing, China (no.:
201001090), the Medical Science and Technology Development Project
of the Nanjing Health Bureau, China (no.: YKK10046) and the Nanjing
Sanitation Engineering of Young Talents during the 12th
Five-Year Plan Period.
References
|
1
|
Tioseco JA, Aly H, Essers J, Patel K and
El-Mohandes AA: Male sex and intraventricular hemorrhage. Pediatr
Crit Care Med. 7:40–44. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
du Plessis AJ and Volpe JJ: Perinatal
brain injury in the preterm and term newborn. Curr Opin Neurol.
15:151–157. 2002.PubMed/NCBI
|
|
3
|
Ivan M, Harris AL, Martelli F and
Kulshreshtha R: Hypoxia response and microRNAs: no longer two
separate worlds. J Cell Mol Med. 12:1426–1431. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chan SY and Loscalzo J: MicroRNA-210: a
unique and pleiotropic hypoxamir. Cell Cycle. 9:1072–1083. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chan SY, Zhang YY, Hemann C, Mahoney CE,
Zweier JL and Loscalzo J: MicroRNA-210 controls mitochondrial
metabolism during hypoxia by repressing the iron-sulfur cluster
assembly proteins ISCU1/2. Cell Metab. 10:273–284. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chen Z, Li Y, Zhang H, Huang P and Luthra
R: Hypoxia-regulated microRNA-210 modulates mitochondrial function
and decreases ISCU and COX10 expression. Oncogene. 29:4362–4368.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Favaro E, Ramachandran A, McCormick R, Gee
H, et al: MicroRNA-210 regulates mitochondrial free radical
response to hypoxia and krebs cycle in cancer cells by targeting
iron sulfur cluster protein ISCU. PloS One. 5:e103452010.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Huang X, Ding L, Bennewith KL, Tong RT, et
al: Hypoxia-inducible mir-210 regulates normoxic gene expression
involved in tumor initiation. Mol Cell. 35:856–867. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kushibiki T: Photodynamic therapy induces
microRNA-210 and -296 expression in HeLa cells. J Biophotonics.
3:368–372. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Pulkkinen K, Malm T, Turunen M, Koistinaho
J and Yla-Herttualä S: Hypoxia induces microRNA miR-210 in vitro
and in vivo ephrin-A3 and neuronal pentraxin 1 are potentially
regulated by miR-210. FEBS Lett. 582:2397–2401. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bacon AL and Harris AL: Hypoxia-inducible
factors and hypoxic cell death in tumour physiology. Ann Med.
36:530–539. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gordan JD and Simon MC: Hypoxia-inducible
factors: central regulators of the tumor phenotype. Curr Opin Genet
Dev. 17:71–77. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Gruber M and Simon MC: Hypoxia-inducible
factors, hypoxia, and tumor angiogenesis. Curr Opin Hematol.
13:169–174. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Harris AL: Hypoxia - a key regulatory
factor in tumour growth. Nat Rev Cancer. 2:38–47. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kim JW, Tchernyshyov I, Semenza GL and
Dang CV: HIF-1-mediated expression of pyruvate dehydrogenase
kinase: a metabolic switch required for cellular adaptation to
hypoxia. Cell Metab. 3:177–185. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Koumenis C: ER stress, hypoxia tolerance
and tumor progression. Curr Mol Med. 6:55–69. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Qiu J, Zhou XY, Zhou XG, Cheng R, Liu HY
and Li Y: Neuroprotective effects of microRNA-210 against
oxygen-glucose deprivation through inhibition of apoptosis in PC12
cells. Mol Med Rep. 7:1955–1959. 2013.PubMed/NCBI
|
|
18
|
Tabakman R, Lazarovici P and Kohen R:
Neuroprotective effects of carnosine and homocarnosine on
pheochromocytoma PC12 cells exposed to ischemia. Journal Neurosci
Res. 68:463–469. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Guo G and Bhat NR: p38alpha MAP kinase
mediates hypoxia-induced motor neuron cell death: a potential
target of minocycline’s neuroprotective action. Neurochem Res.
32:2160–2166. 2007.PubMed/NCBI
|
|
20
|
Tabakman R, Jiang H, Schaefer E, Levine RA
and Lazarovici P: Nerve growth factor pretreatment attenuates
oxygen and glucose deprivation-induced c-Jun amino-terminal kinase
1 and stress-activated kinases p38alpha and p38beta activation and
confers neuroprotection in the pheochromocytoma PC12 model. J Mol
Neurosci. 22:237–250. 2004. View Article : Google Scholar
|
|
21
|
Kim J, Krichevsky A, Grad Y, Hayes GD,
Kosik KS, Church GM and Ruvkun G: Identification of many microRNAs
that copurify with polyribosomes in mammalian neurons. Proc Nat
Acad Sci USA. 101:360–365. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kosik KS and Krichevsky AM: The elegance
of the microRNAs: A neuronal perspective. Neuron. 47:779–782. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Schratt GM, Tuebing F, Nigh EA, Kane CG,
Sabatini ME, Kiebler M and Greenberg ME: A brain-specific microRNA
regulates dendritic spine development. Nature. 439:283–289. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Qiu J, Zhou XY, Zhou XG, Cheng R, Liu HY
and Li Y: Neuroprotective effects of microRNA-210 on
hypoxic-ischemic encephalopathy. Biomed Res Int.
2013:3504192013.PubMed/NCBI
|
|
25
|
Fasanaro P, D’Alessandra Y, Di Stefano V,
Melchionna R, et al: MicroRNA-210 modulates endothelial cell
response to hypoxia and inhibits the receptor tyrosine kinase
ligand Ephrin-A3. J Biol Chem. 283:15878–15883. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Giannakakis A, Sandaltzopoulos R, Greshock
J, Liang S, Huang J, Hasegawa K, Li C, O’Brien-Jenkins A, Katsaros
D, Weber BL, Simon C, Coukos G and Zhang L: miR-210 links hypoxia
with cell cycle regulation and is deleted in human epithelial
ovarian cancer. Cancer Biol Ther. 7:255–264. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
van Rooij E, Liu N and Olson EN: MicroRNAs
flex their muscles. Trends Genet. 24:159–166. 2008.
|
|
28
|
Love TM, Moffett HF and Novina CD: Not
miR-ly small RNAs: big potential for microRNAs in therapy. J
Allergy Clin Immunol. 121:309–319. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Weiler J, Hunziker J and Hall J:
Anti-miRNA oligonucleotides (AMOs): ammunition to target miRNAs
implicated in human disease? Gene Ther. 13:496–502. 2006.
View Article : Google Scholar
|
|
30
|
Ørom UA, Kauppinen S and Lund AH: LNA
modified oligonucleotides mediate specific inhibition of microRNA
function. Gene. 372:137–141. 2006.PubMed/NCBI
|
|
31
|
Zhang B and Farwell MA: microRNAs: a new
emerging class of players for disease diagnostics and gene therapy.
J Cell Mol Med. 12:3–21. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Liu NK and Xu XM: MicroRNA in central
nervous system trauma and degenerative disorders. Physiol Genomics.
43:571–580. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhang C: Novel functions for small RNA
molecules. Curr Opin Mol Ther. 11:641–651. 2009.PubMed/NCBI
|