Introduction
Henoch-Schönlein purpura (HSP) is a small vessel
vasculitis syndrome and is the most common vasculitis in children.
The typical clinical features of HSP include non-thrombocytopenic
purpura, joint pain, stomach pain, gastrointestinal bleeding and
renal system dysfunction (1). In
addition, HSP has also been rarely associated with the following
clinical manifestations: Thrombosis, vomiting, adult human
immunodeficiency virus infection, intestinal perforation, cerebral
hemorrhage, intestinal lesions, neuropathy, ischemic stroke,
cerebrovascular disease, skin rash, joint swelling and pulmonary
hemorrhage (1–7). The kidneys are the most commonly
affected organs in children with HSP and their involvement
primarily causes morbidity and mortality (8).
Numerous studies have demonstrated the role of
polymorphisms of various genes in the susceptibility, progression
and prognosis of HSP and HSP with nephritis (HSPN). Variations in
C1GALT1, CTLA-4 exon 1 +49A/G, interleukin (IL) 8 gene 2767 A/G,
human leukocyte antigen (HLA)-A, HLA-B, mediterranean fever E148Q,
tumor necrosis factor-α and polymorphisms at −2123 and −825 in the
p-selectin gene have been reported to be associated with HSP and/or
HSPN (9–15). However, several other genetic
polymorphisms, including TLR-2 Arg753Gln, TLR-4 Asp299Gly, TLR-4
Thr399Ile, CD18 AvaII, toll-like receptor 4, IL-6 receptor, IL-6
signal transducer and the −173 G/C polymorphism in the macrophage
migration inhibitory factor gene are not associated (16–20).
Paired box 2 (PAX2) is an essential nuclear
transcription factor in the development of the human embryonic
kidney and its expression gradually decreases with the maturity of
nephrons (21). Human PAX2 is
located at 10q24–25, covering ~70 kb of 12 exons (22). The PAX2 Mutation Database
(http://www.hgu.mrc.ac.uk/Softdata/PAX2) has reported
four single nucleotide polymorphisms (SNPs) within the PAX2 coding
region, including 291C>T at exon 3, 798C>T and 909A>C at
exon 8 and 932C>T at exon 9. The nonsense mutation of 932C>T
causes the 331st amino acid Alanine to mutate to Valine, while the
three other SNPs are all sense mutations.
Mutations in the PAX2 gene have been associated with
renal developmental abnormalities, urinary tract complications and
oncogenic processes (23).
Increased apoptosis has been reported in the fetal kidneys of
PAX21Neu +/− mutant mice (24). In children, PAX2 and hepatocyte
nuclear factor-1β were identified as the common cause of renal
aplasia and hypodysplasia (25).
An earlier study demonstrated the expression of PAX2 in the
podocytes of renal glomeruli and tubular epithelial cells of
children with HSPN, suggesting the potential involvement of PAX2 in
the progression of HSPN (26). A
study by Yi et al investigated the distribution of
polymorphisms in the PAX2 gene and examined the development of
renal pathology. The study concluded that the 410CT/1521AC PAX2
genotype improved the prognosis of HSPN (27).
However, the association between polymorphisms of
various exons of the PAX2 gene and HSPN remain to be fully
elucidated. Therefore, the present study aimed to analyze the
polymorphism sites of the 12 exons of PAX2 and to investigate the
association between various polymorphisms in the PAX2 gene and the
susceptibility and clinical characteristics of HSP and/or HSPN.
Subjects and methods
Study population
The present study was conducted at the Second
Xiangya Hospital of Central South University (Changsha, China). The
study protocol was reviewed and approved by the Ethics Committee of
The Second Xiangya Hospital of Central South University and parents
of all the children involved provided written informed consent
prior to surgery. Between June 2003 and February 2007, a total of
118 children with HSP were recruited from the Department of
Paediatrics, The Second Xiangya Hospital of Central South
University. The conditions of all cases were diagnosed as HSP
rather than other systemic diseases, including thrombocytopenic
purpura or systemic lupus erythematosus. In addition, 100 healthy
children, with no history of HSP, were enrolled in the present
study as controls.
After >6 months follow up, the subjects were
divided into two groups (HSPN and HSP) based on the results of
urinary sediment erythrocyte count and determination of urine
protein concentration at 24 h. An erythrocyte count >500,000 at
12 h and a protein concentration >150 mg at 24 h were regarded
as positive for HSPN (28). For
the HSPN group (n=80), the primary course of disease was between 1
day and 24 months (mean course: 1.7±4.2 months). After a follow-up
of 7–48 months until June 2007, the course of disease was 6–70
months (mean course: 18.5±14.7 months). For the HSP (without
nephritis) group (n=38), the course of disease was 4–45 months
(mean course: 9.2±10.3 months). Renal needle biopsy was performed
on 64 cases in the HSPN group. Based on the standards established
by the International Study of Kidney Diseases in Children (29), the cases were classified as
follows: Grade II (n=12), grade III (n=43), grade IV (n=8) and
grade V (n=1).
DNA extraction
Fresh blood (5 ml in 2% ethylenediaminetetraacetic
acid (Junrui Biotechnology Co., Ltd., Shanghai, China) was obtained
from each subject. The genomic DNA was extracted from peripheral
venous blood using a salting-out method (30)
Polymerase chain reaction (PCR)
analysis
The sequences of exons 1–12 of PAX2 were amplified
by PCR. The primers (Shanghai Bioengineering Co, Ltd., Shanghai,
China) were designed within the region of each exon based on a
previous study (31,32) using Primer Premier 5.0 software
(Premier Biosoft International, Palo Alto, CA, USA). PCR was
performed in a 25 μl reaction volume containing 50 ng/ml genomic
DNA, 2.5 μl 10X PCR buffer (containing 1.5 mM magnesium chloride),
2 μl 500 μM deoxyribonucleotides, 1 μl 5 μM primer and 0.25 μl
5U/μl Taq DNA polymerase (MBI Fermentas, Vilnius, Lithuania).
Amplification was performed under the following conditions:
Sequencing thermal cycling parameters included 96°C for 3 min and
39 cycles of 96°C for 45 sec, appropriate annealing temperature for
the particular primer pair for 1 min (Table I), 72°C for 1 min and final
extension at 72°C for 10 min using the PCR System EDC-810 (Eastwin
Life Science Inc., Beijing, China). The PCR products (~5 μl) were
then subjected to 1.5% agarose gel electrophoresis.
 | Table ISequences and annealing temperatures
for the primers of the paired box 2 gene. |
Table I
Sequences and annealing temperatures
for the primers of the paired box 2 gene.
| Exon | Primer sequence
(5′-3′) | Product length
(bp) | Annealing temperature
(°C) |
|---|
| 1 | F:
GTTCACTCATCCTCCCTCCCCCACC | | |
| R:
GGAGCCGGGCGCGGGTACTC | 179 | 62.0 |
| 2 | F:
CTGTGTGTGGGGTGTTGTGTT | | |
| R:
AAGGCGTCTCTCCCGGGACAGCTGC | 246 | 61.8 |
| 3 | F:
TGACCGGCTTTCCCGGCGCA | | |
| R:
GAGGAAGCTGGAGTCCAGCC | 259 | 62.0 |
| 4 | F:
CGGAATAGGAGTGGCATTTGA | | |
| R:
CTCTAGGTGGGATCTGGTTT | 182 | 60.0 |
| 5 | F:
TGATGCCATTTCCTCCTTCC | | |
| R:
GCCACACCTCTTCCCTCCT | 175 | 61.0 |
| 6 | F:
TCTCAGTGTTTGTCTGTCTC | | |
| R:
TGAATGTTCTCTAACCGCCA | 175 | 59.0 |
| 7 | F:
ACGCCCCGAGTGTCCATGT | | |
| R:
CTGCCGGCTCCTTTCTACT | 250 | 58.8 |
| 8 | F:
CCTTTCTCTGTGCGTGCATCAATAGA | | |
| R:
GGCACCCTCCACTGAACGCAG | 231 | 59.0 |
| 9 | F:
CCCTTCCCCTTTGTGTTTTT | | |
| R:
AGGCAGCTGCAGCATTGTC | 151 | 60.0 |
| 10 | F:
CCCCTCCCTGCAAACCAC | | |
| R:
CGCTGTGAGGGCCATGAC | 150 | 61.5 |
| 11 | F:
GCAGGCGTCACATCCCCACTC | | |
| R:
CCGGCCACCAGGTGGCGTA | 148 | 62.5 |
| 12 | F:
TGTTTGTCCTCTGACCCAGC | | |
| R:
CAGACGGGGACGATGTGGA | 187 | 59.0 |
Denatured high performance liquid
chromatography (DHPLC)
The PCR product (4 μl) was mixed with the same
volume of known wild-type sample. The mixture was denatured at 95°C
for 3 min, and the temperature was then decreased by 1°C per second
until it reached 45°C, in order to form a mixture of heterogeneous
and homologous double-stranded DNA. The mixture was then separated
using a WAVE DNA Fragment Analysis System (Transgenomic Inc., Santa
Clara, CA, USA) with a linear acetonitrile gradient elution,
consisting of buffer A (0.1 M triethylamine acetate; TEAA) and
buffer B (0.1 M TEAA in 25% acetonitrile) at a constant flow rate
of 0.9 ml/min. The optimal temperature for the DNA fragment was
provided by the software WaveMaker 4.0 (Transgenomic Inc.). The
mutation type was determined by the elution peaks.
DNA sequence analysis
The PCR products demonstrating abnormal peaks and
sections of products with normal peaks were sequenced using a
3730xl DNA Analyzer (Applied Biosystems, Foster City, CA, USA) for
determination of the PAX2 genotypes.
Statistical analysis
All statistical analyses were performed using SPSS
version 10.0 (SPSS, Inc., Chicago, IL, USA). The allelic and
genotype frequencies of the PAX2 gene polymorphisms in the two
groups were calculated, and reached genetic equilibrium using the
Hardy-Weinberg law (p2+2pq+q2), indicating that each group
consisted of an independent population. The data are expressed as
the mean ± standard deviation. To determine the extent to which the
subjects in the present study fit within the Hardy-Weinberg law, an
allele or genotype comparison between the two groups and linkage
disequilibrium analysis were conducted using a χ2 test
or Fisher’s analysis, respectively. P<0.05 was considered to
indicate a statistically significant difference.
Results
Demographic characteristics
Of the 118 children with HSP and HSPN, 76 patients
were male with a mean age of 9.8±1.9 years and 42 were female with
a mean age of 10.6±2.2 years. Among the 100 healthy children, 58
were male with a mean age of 9.5±2.0 years and 42 were female with
a mean age of 9.5±1.9 years. The demographic characteristics of
subjects in the three groups (including controls) are listed in
Table II.
| Table IIDemographic characteristics of
subjects enrolled. |
Table II
Demographic characteristics of
subjects enrolled.
| Variable | HSP
n, (%)
n=38 | HSPN
n, (%)
n=80 | Control
n, (%)
n=100 | P-value |
|---|
| Gender |
| Male | 22 (57.89) | 54 (67.50) | 58 (58.00) | 0.379 |
| Female | 16 (42.11) | 26 (32.50) | 42 (42.00) | |
| Age | 10.18±2.84a | 9.97±1.58a | 9.46±1.92a | 0.472 |
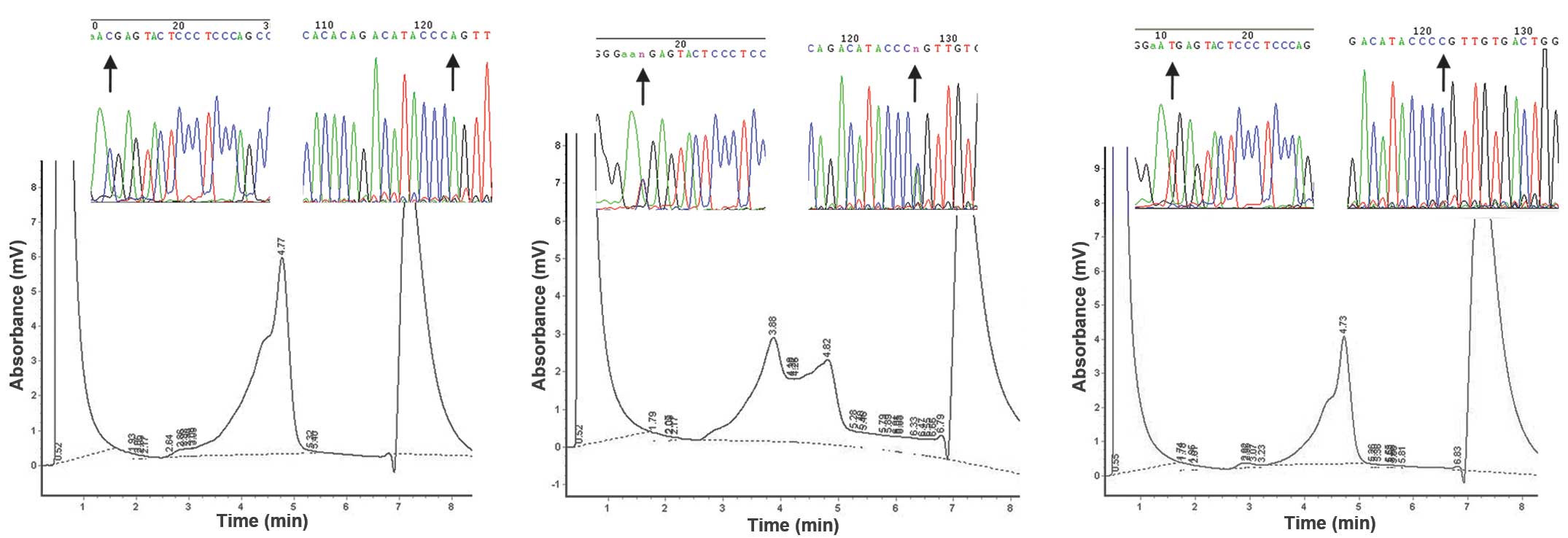
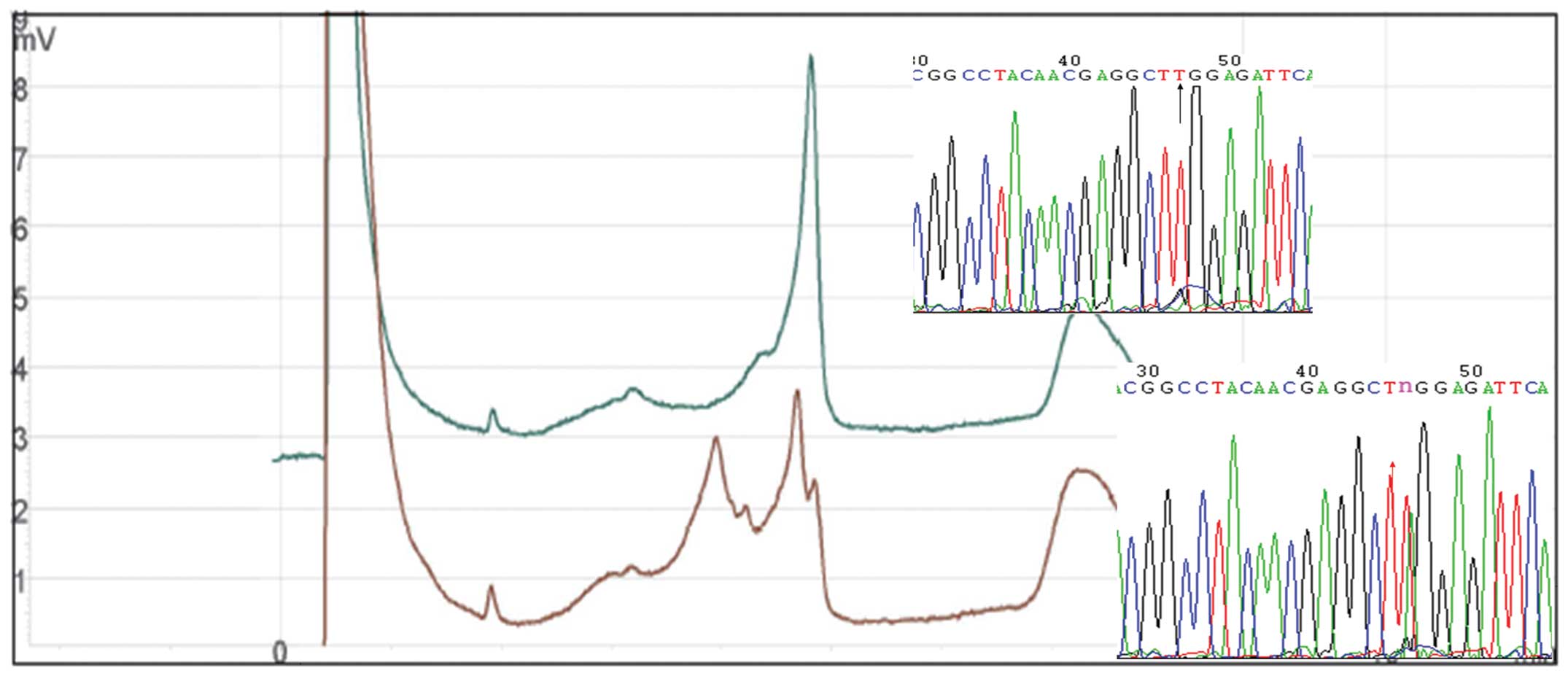
DHPLC screening of the PAX2 gene and DNA
sequencing
The DHPLC profiles of all subjects revealed exons
1–7, 9, 10 and 12 with a single peak, while exons 8 and 11 had a
distinct profile with multiple peaks (one single-peak and one
double-peak). The products with abnormal DHPLC elution curves were
sequenced and products with normal DHPLC elution curves were
randomly selected as references. The DNA sequence results of exons
1–7, 9, 10 and 12 and the single-peaked products of exons 8 and 11
were identical to those published in GenBank (http://www.ncbi.nlm.nih.gov/nuccore/NM_003988.3). The
results identified the presence of two SNPs in exon 8 (798C>T
and 909A>C; Fig. 1) and one SNP
in exon 11 (1164T>A) of the PAX2 gene (Fig. 2).
Linkage disequilibrium of 798C>T and
909A>C in exon 8
Two PAX2 gene alleles, 798C>T and 909A>C, were
not randomly distributed. In the 218 subjects, 798C>T was always
present with 909A>C. The correlation coefficient of polymorphism
combination was 1 (γ=1), the actual frequency of haplotype
798C/909A was 0.93 and the linkage equilibrium drift was Δ=0.065.
The difference was significant as evaluated by the χ2
test, indicating the complete linkage disequilibrium between
798C>T and 909A>C polymorphisms (Table III).
| Table IIIDistribution of the paired box 2
alleles and linkage disequilibrium in healthy children. |
Table III
Distribution of the paired box 2
alleles and linkage disequilibrium in healthy children.
| Actual (expected)
value of allele compound |
|---|
|
|
|---|
| Allele | 909A | 909C |
|---|
| 798C | 186 (173.0) | 0 (13.0) |
| 798T | 0 (13.0) | 14 (1.0) |
|
χ2=200 | P<0.001 |
| γ=1 | Δ=0.065 |
PAX2 gene exon 8 and 11 allele and
genotype frequency distribution in normal controls and HSP
patients
Due to the complete linkage disequilibrium between
798C>T and 909A>C polymorphisms, 798C>T was selected on
behalf of the polymorphism of exon 8. No significant difference in
the allele and genotype frequency distribution of the PAX2 gene
exon 8 (798C>T) or 11 (1164T>A) was identified between the
HSP and the control groups (P>0.05; Table IV).
| Table IVPaired box 2 gene exon 8 and 11
alleles, genotype frequency (%) and distribution in healthy
controls and HSP patients. |
Table IV
Paired box 2 gene exon 8 and 11
alleles, genotype frequency (%) and distribution in healthy
controls and HSP patients.
| Exon | Genotype or
allele | HSP (n=118)
(%) | Control (n=100)
(%) | χ2 | P-value |
|---|
| Exon 8 |
| 798 C>T |
| CC | 94 (79.66) | 87 (87.00) | 2.257 | 0.324 |
| CT | 23 (19.49) | 12 (12.00) | | |
| TT | 1 (0.85) | 1 (1.00) | | |
| C | 211 (89.41) | 186 (93.00) | 2.506 | 0.113 |
| T | 25 (10.59) | 14 (7.00) | | |
| Exon 11 |
| 1164T>A |
| TT | 116 (98.31) | 99 (99.00) | 0.193 | 0.661 |
| TA | 2 (1.69) | 1 (1.00) | | |
| AA | 0 (0.00) | 0 (0.00) | | |
| T | 234 (99.15) | 199 (99.50) | 0.091 | 0.662 |
| A | 2 (0.85) | 1 (0.50) | | |
Allele and genotype frequency
distribution of the PAX2 gene exon 8 and 11 in normal controls, HSP
and HSPN patients
Based on the results of the urinary sediment
erythrocyte count and 24 h urine protein determination, the HSP
group was further divided into HSP without nephritis (HSP, n=38)
and HSP with nephritis (HSPN, n=80). The genotype frequency of exon
8 798C>T in the HSPN group (20/80, 25%) was higher when compared
with the HSP (3/38, 7.89%) group and the control (12/100, 12%)
group (P<0.05). However, no significant difference was
identified in the genotype frequency of exon 8 798C>T between
the HSP group and the control group (P>0.05; Table V). In addition, no significant
difference was identified in the allele and genotype frequency of
exon 8 and 11, with the exception of 798C>T for exon 8, among
controls, HSP and HSPN patients (P>0.05; Table V).
| Table VPaired box 2 exon 8 and 11 alleles,
genotype frequency (%) and distribution in normal controls, HSP
patients without nephritis and HSP patients with nephritis
(HSPN). |
Table V
Paired box 2 exon 8 and 11 alleles,
genotype frequency (%) and distribution in normal controls, HSP
patients without nephritis and HSP patients with nephritis
(HSPN).
| | Group | | |
|---|
| |
| | |
|---|
| Exon | Genotype or
allele | HSP without
nephritis (n=38) (%) | HSPN (n=80)
(%) | Control (n=100)
(%) | χ2 | P-value |
|---|
| Exon 8 |
| 798 C>T |
| CC | 34 (89.47) | 60 (75.00) | 87 (87.00) | | |
| CT | 3 (7.89)a | 20 (25.00) | 12 (12.00)a | | |
| TT | 1 (2.63) | 0 (0.00) | 1 (1.00) | | |
| C | 71 (93.42) | 140 (87.50) | 186 (93.00) | 3.934 | 0.140 |
| T | 5 (6.58) | 20 (12.50) | 14 (7.00) | | |
| Exon 11 |
| 1164 T>A |
| TT | 38 (100.00) | 78 (97.50) | 99 (99.00) | 1.379 | 0.502 |
| TA | 0 (0.00) | 2 (2.50) | 1 (1.00) | | |
| AA | 0 (0.00) | 0 (0.00) | 0 (0.00) | | |
| T | 76 (100.00) | 158 (98.80) | 199 (99.50) | 1.369 | 0.504 |
| A | 0 (0.00) | 2 (1.20) | 1 (0.50) | | |
Correlation between the PAX2 gene
polymorphism (exon 8 798C>T) and the clinical characteristics of
children with HSP
The correlation between various clinical features
(gender, age, skin purpura, joint involvement, gastrointestinal
symptoms and kidney lesions) of the 118 children with HSP and the
frequency distribution of the PAX2 gene polymorphism exon 8
798C>T were analyzed. No significant differences were observed
in the features compared with the controls (P>0.05), with the
exception of kidney lesions (P<0.05; Table VI).
| Table VICorrelation between the paired box 2
gene polymorphism exon 8 798C>T and the clinical characteristics
of children with HSP. |
Table VI
Correlation between the paired box 2
gene polymorphism exon 8 798C>T and the clinical characteristics
of children with HSP.
| | Genotype | | |
|---|
| |
| | |
|---|
| | CC | CT | TT | | |
|---|
| |
| | |
|---|
| Organ damage | Number, n | n | % | n | % | n | % |
χ2a | P-valuea |
|---|
| Joint | 64 | 55 | 85.94 | 8 | 12.50 | 1 | 1.56 | 0.038 | 0.846 |
| Gastrointestinal
tract | 77 | 69 | 89.61 | 8 | 10.39 | 0 | 0.00 | 0.283 | 0.594 |
| Kidney | 80 | 60 | 75.00 | 20 | 25.00 | 0 | 0.00 | 4.257 | 0.039 |
| Control | 100 | 87 | 87.00 | 12 | 12.00 | 1 | 1.00 | | |
Correlation between the PAX2 gene
polymorphism (exon 8 798C>T) and pathological lesions of
children with HSPN
Compared with the overall genotype frequency (20/80,
25%) of exon 8 798C>T in the HSPN group, the children with HSP
and genotype 798C>T were more susceptible to nephrotic syndrome
(41.67%) and proteinuria (41.18%) and were less susceptible to
hypertension (18.92%), however, the difference was not
statistically significant (all P>0.05; Table VII).
| Table VIICorrelation between the paired box 2
gene polymorphism 798C>T in exon 8 and the clinical
characteristics and pathological lesions of children with
Henoch-Schönlein purpura with nephritis. |
Table VII
Correlation between the paired box 2
gene polymorphism 798C>T in exon 8 and the clinical
characteristics and pathological lesions of children with
Henoch-Schönlein purpura with nephritis.
| | Genotype | | |
|---|
| |
| | |
|---|
| | CC | CT | | |
|---|
| |
| | |
|---|
| Kidney lesion | Number, n | n | % | n | % |
χ2a | P-valuea |
|---|
| Patient
numbers | 80 | 60 | 75.00 | 20 | 25.00 | | |
| Microscopic
hematuria | 58 | 42 | 72.41 | 16 | 27.59 | 0.117 | 0.773 |
| Gross
hematuria | 72 | 55 | 76.39 | 17 | 23.61 | 0.040 | 0.842 |
| Small quantity of
proteinuria | 34 | 26 | 76.47 | 8 | 23.53 | 0.028 | 0.827 |
| Large quantity of
proteinuria | 17 | 10 | 58.80 | 7 | 41.20 | 1.826 | 0.177 |
| Hypertension | 37 | 30 | 81.10 | 7 | 18.90 | 0.567 | 0.468 |
| Nephrotic
syndrome | 12 | 7 | 58.30 | 5 | 41.70 | 1.465 | 0.226 |
| Nephritis
syndrome | 44 | 35 | 79.60 | 9 | 20.40 | 0.327 | 0.567 |
Discussion
The present study involved the analysis of PAX2 gene
polymorphisms in children with HSP and in gender and age-matched
healthy children in Hunan Province using DHPLC. The polymorphism
sites of the 12 exons of PAX2 were identified and the association
between various polymorphisms in the PAX2 gene and the
susceptibility and clinical characteristics of HSP and/or HSPN were
investigated. The results of the present study may be clinically
significant for the determination of the susceptibility towards the
development of kidney lesions in children with HSP.
No studies were available on the association between
the PAX2 haplotype 798C>T/909 A>C and SNP 1164T>A with the
susceptibility and clinical pathophysiological characteristics of
HSP. To the best of our knowledge, this is the first study to
identify a low frequency SNP of exon 11 (1164T>A) in the PAX2
gene in Han children. In addition, the 798C>T/909A>C PAX2
genotype did not increase the susceptibility to HSP, but it was
likely to increase the susceptibility of kidney involvement.
Therefore, this finding may be significant in the diagnosis of
HSPN.
In all 218 subjects, the exons 1–7, 9, 10 and 12 of
PAX2 demonstrated a single peak, indicating no mutation. This
result was inconsistent with those of Gelb et al, in which
SNPs, including 291C>T at exon 3 and 932C>T at exon 9, were
identified (22). This may be due
to geographic and ethnic differences. In the present study, two
SNPs of the PAX2 gene were identified in exon 8 as a complete
linkage haplotype 798C>T/909A>C. The frequency of the allele
798T and 909C was 7% in the control group. However, the study by
Shim et al demonstrated allelic exchange of the PAX2 gene
and no linkage haplotype in American children (33). The difference from the results of
the present study may also be explained by ethnic differences. A
T>A polymorphism at exon 11 was first reported with a frequency
of 0.5%. In the version of NM003988, a mutated allele 1164A
(Leu388->Leu388) was reported in the present study. No
significant differences were identified in this genotype frequency
distribution between patients with HSP and the control group,
suggesting that the PAX2 gene polymorphism had no correlation with
the susceptibility to, or pathophysiological characteristics of
HSP. However, the detailed mechanism underlying this may be
revealed by further studies.
No significant difference was identified in the
frequency of allele 798C/909A between the HSP and the control
groups (P>0.05), nor in the joint or gastrointestinal organ
damage groups (P>0.05). However, the frequency of genotype
798CT/909AC in the HSPN group was significantly higher than that in
the control or the HSP without nephritis group (P<0.05). These
data indicated that the PAX2 gene polymorphism neither predisposed
individuals to develop HSP, nor to develop joint and
gastrointestinal tract symptoms in HSP, although the PAX2 gene
polymorphism may be associated with the development of kidney
lesions in HSP.
Although no difference was identified in the
disruption of haplotype 798C 909A and 798T 909C in the PAX2 gene
between the HSP and the control group, the frequency of the
genotype 798C>T in the HSPN group was significantly higher than
that in the control group (P=0.039). However, no significant
difference in frequency was demonstrated between the HSP and the
control groups. These results indicated that the risk for HSP was
not increased in children with genotype 798C>T/909A>C,
however, it may increase susceptibility to the development of
kidney lesions in children with HSP. Ballinger et al
identified that 97% of children with HSP demonstrate kidney lesions
within 3 months (34). As the
follow-up of all subjects in the present study lasted >6 months,
the SNP results may be valuable in determining the susceptibility
to the development of kidney lesions in children with HSP.
The study by Yi et al reported that the
1410C>T/1521A>C genotype in PAX2 may predispose to kidney
damage in Chinese children with HSP (27). Kidney damage in HSPN patients is
characterized by hematuria, proteinuria, hypertension, nephrotic
syndrome or nephritic syndrome. Although differences were
identified in the distribution of the PAX2 polymorphism among the
clinical phenotypes, they were statistically insignificant. This
suggested there was no correlation between genotype and the various
clinical manifestations of HSPN or the progression or prognosis of
renal pathology. A larger study population is required in future
studies.
In conclusion, the present study identified a
polymorphism of exon 11 in the PAX2 gene (1164T>A mutation).
Geographic and ethnic disparities may have caused different PAX2
gene polymorphisms. The PAX2 heterozygous genotype 798C>T does
not increase susceptibility to HSP, but it can be used clinically
as a HSP screening indicator in children with a high risk of renal
involvement.
References
|
1
|
Li L, Zhang J, Zhang Y and Ji H:
Thrombosis warning in children suffering from henoch-schonlein
purpura. Indian J Dermatol. 58:4092013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chen O, Zhu XB, Ren P, Wang YB, Sun RP and
Wei DE: Henoch Schonlein Purpura in children: clinical analysis of
120 cases. Afr Health Sci. 13:94–99. 2013.PubMed/NCBI
|
|
3
|
Zaid M, Tan K, Smitasin N, Tambyah PA and
Archuleta S: Henoch-Schönlein purpura associated with adult human
immunodeficiency virus infection: case report and review of the
literature. Ann Acad Med Singapore. 42:358–360. 2013.PubMed/NCBI
|
|
4
|
Wang HL, Liu HT, Chen Q, Gao Y and Yu KJ:
Henoch-Schonlein purpura with intestinal perforation and cerebral
hemorrhage: a case report. World J Gastroenterol. 19:2574–2577.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ghosh K, Chatterjee A, Sau JT and Dey S:
Stroke and skin rash: A rare case of Henoch-Schonlein purpura. Ann
Indian Acad Neurol. 15:307–309. 2012. View Article : Google Scholar
|
|
6
|
Fukushima K: Severe intestinal lesions and
neuropathy associated with Henoch-Schonlein purpura. Intern Med.
52:2009–2010. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ren X, Zhang W, Dang W, et al: A case of
anaphylactoid purpura nephritis accompanied by pulmonary hemorrhage
and review of the literature. Exp Ther Med. 5:1385–1388.
2013.PubMed/NCBI
|
|
8
|
Kawasaki Y, Ono A, Ohara S, et al:
Henoch-Schonlein purpura nephritis in childhood: pathogenesis,
prognostic factors and treatment. Fukushima J Med Sci. 59:15–26.
2013. View Article : Google Scholar
|
|
9
|
An J, Lu Q, Zhao H, Cao Y, Yan B and Ma Z:
A study on the association between C1GALT1 polymorphisms and the
risk of Henoch-Schonlein purpura in a Chinese population. Rheumatol
Int. 33:2539–2542. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang JJ, Shi YP, Huang Y, Wu C and Li XC:
Association of tumor necrosis factor-alpha gene polymorphisms with
Henoch-Schonlein purpura nephritis in children. Zhongguo Dang Dai
Er Ke Za Zhi. 15:88–90. 2013.(In Chinese). PubMed/NCBI
|
|
11
|
Wang JJ, Shi YP, Yue H, Chun W and Zou LP:
CTLA-4 exon 1 +49A/G polymorphism is associated with renal
involvement in pediatric Henoch-Schönlein purpura. Pediatr Nephrol.
27:2059–2064. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Tabel Y, Mir S and Berdeli A: Interleukin
8 gene 2767 A/G polymorphism is associated with increased risk of
nephritis in children with Henoch-Schönlein purpura. Rheumatol Int.
32:941–947. 2012. View Article : Google Scholar
|
|
13
|
Ren SM, Yang GL, Liu CZ, et al:
Association between HLA-A and -B polymorphisms and susceptibility
to Henoch-Schönlein purpura in Han and Mongolian children from
Inner Mongolia. Genet Mol Res. 11:221–228. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
He X, Lu H, Kang S, et al: MEFV E148Q
polymorphism is associated with Henoch-Schönlein purpura in Chinese
children. Pediatr Nephrol. 25:2077–2082. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li J and Liu H: P-selectin gene -825
polymorphism is associated with risk of Henoch-Schonlein purpura
nephritis. Clin Exp Rheumatol. 29:S1342011.PubMed/NCBI
|
|
16
|
Soylu A, Kizildağ S, Kavukçu S, et al:
TLR-2 Arg753Gln, TLR-4 Asp299Gly, and TLR-4 Thr399Ile polymorphisms
in Henoch Schonlein purpura with and without renal involvement.
Rheumatol Int. 30:667–670. 2010. View Article : Google Scholar
|
|
17
|
Nalbantoglu S, Tabel Y, Mir S and Berdeli
A: Lack of association between macrophage migration inhibitory
factor gene promoter (−173 G/C) polymorphism and childhood
Henoch-Schönlein purpura in Turkish patients. Cytokine. 62:160–164.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
He X, Li Y, Kang S, et al: The CD18 AvaII
polymorphic site not associated with Henoch-Schönlein purpura. Clin
Exp Rheumatol. 29:S117–S120. 2011.PubMed/NCBI
|
|
19
|
Torres O, Palomino-Morales R,
Miranda-Filloy JA, Vazquez-Rodriguez TR, Martin J and Gonzalez-Gay
MA: Lack of association between toll-like receptor 4 gene
polymorphism and Henoch-Schönlein purpura. Clin Exp Rheumatol.
28:1102010.
|
|
20
|
López-Mejías R, Sevilla Pérez B, Genre F,
et al: No evidence of association between functional polymorphisms
located within IL6R and IL6ST genes and Henoch-Schönlein purpura.
Tissue Antigens. 82:416–419. 2013. View Article : Google Scholar
|
|
21
|
Torban E, Eccles MR, Favor J and Goodyer
PR: PAX2 suppresses apoptosis in renal collecting duct cells. Am J
Pathol. 157:833–842. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Gelb A, Manligas G, Gharaybeh S and
Schimmenti L: Identification of two novel polymorphisms
(g.903C>T and g.1544C>T) in the PAX2 gene. Hum Mutat.
17:1552001. View Article : Google Scholar
|
|
23
|
Harshman LA and Brophy PD: PAX2 in human
kidney malformations and disease. Pediatr Nephrol. 27:1265–1275.
2012. View Article : Google Scholar
|
|
24
|
Porteous S, Torban E, Cho NP, et al:
Primary renal hypoplasia in humans and mice with PAX2 mutations:
evidence of increased apoptosis in fetal kidneys of Pax2(1Neu) +/−
mutant mice. Hum Mol Genet. 9:1–11. 2000. View Article : Google Scholar
|
|
25
|
Thomas R, Sanna-Cherchi S, et al: HNF1B
and PAX2 mutations are a common cause of renal hypodysplasia in the
CKiD cohort. Pediatr Nephrol. 6:897–903. 2011. View Article : Google Scholar
|
|
26
|
Zu CP, Yi ZW and Dang XQ: Significance of
PAX2 expression in pathological kidney of children with
Henoch-Schoenlein purpura nephritis. Journal of Clinical Research.
22:1514–1517. 2005.
|
|
27
|
Yi ZW, Fang XL, Wu XC, et al: Role of PAX2
gene polymorphisms in Henoch-Schonlein purpura nephritis.
Nephrology (Carlton). 11:42–48. 2006. View Article : Google Scholar
|
|
28
|
Loghman-Adham M: Evaluating proteinuria in
children. Am Fam Physician. 58:1145–1152. 1158–1149.
1998.PubMed/NCBI
|
|
29
|
No authors listed. Primary nephrotic
syndrome in children: clinical significance of histopathologic
variants of minimal change and of diffuse mesangial
hypercellularity. A Report of the International Study of Kidney
Disease in Children. Kidney Int. 20:765–771. 1981. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Miller SA, Dykes DD and Polesky HF: A
simple salting out procedure for extracting DNA from human
nucleated cells. Nucleic Acids Res. 16:12151988. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Tellier AL, Amiel J, Delezoide AL, et al:
Expression of the PAX2 gene in human embryos and exclusion in the
CHARGE syndrome. Am J Med Genet. 93:85–88. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Schimmenti LA, Cunliffe HE, McNoe LA, et
al: Further delineation of renal-coloboma syndrome in patients with
extreme variability of phenotype and identical PAX2 mutations. Am J
Hum Genet. 60:869–878. 1997.PubMed/NCBI
|
|
33
|
Shim HH, Nakamura BN, Cantor RM and
Schimmenti LA: Identification of two single nucleotide
polymorphisms in exon 8 of PAX2. Mol Genet Metab. 68:507–510. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Ballinger S: Henoch-Schonlein purpura.
Curr Opin Rheumatol. 5:591–594. 2003. View Article : Google Scholar
|