Introduction
Apoptosis is an indispensable process in embryonic
development and tissue homeostasis (1) that is tightly regulated by
caspase-mediated signaling cascades. Caspases are part of a family
of cysteine proteases that participate in the cleavage of aspartic
acid-containing motifs (2). Among
the upstream initiator and downstream effector caspases, caspase-3
is the executioner caspase that triggers the cellular processes
resulting in the deconstruction of the cell (3). Various kinases and signaling adaptor
proteins have been suggested to be specific substrates of
caspase-3, thus it can be inferred that multiple aspects of cell
death and survival may be mediated through the cleavage of various
signaling molecules (4,5).
The highly conserved acidic nuclear phosphoprotein
32 kDa (ANP32) family of proteins are characterized by an
N-terminal leucine-rich repeat (LRR) domain and a C-terminal
low-complexity acidic region (LCAR) (6). These proteins have been demonstrated
to be involved in a wide array of physiological processes,
including cell apoptosis (7–12),
differentiation (13–16) and proliferation (17–19).
The majority of studies on the ANP32 family have focused on ANP32A,
which has been implicated in a variety of cellular functions. For
example, it has been demonstrated to enhance caspase-9 activation
via the promotion of apoptosome formation by mediating the
oncoprotein prothymosin (9).
Additionally, the apoptotic activity of ANP32A has been observed to
be crucial in tumor suppression (10), and it contributes to the apoptotic
response to chemotherapy in advanced non-small-cell lung cancer
(8). ANP32A-deficient mice are,
however, viable and fertile with no clear abnormalities, indicating
its functional redundancy, which may be due to an overlap of
functions with closely related family members (20). Compared with the effects of ANP32A
knockout, ANP32B knockout mice display a partially penetrant
perinatal lethality, suggesting a hierarchy of importance for the
mammalian ANP32 genes, with ANP32B as the most critical for normal
development (21).
In a previous study, ANP32B was observed to be a
direct substrate of caspase-3 and primarily cleaved at the sequence
of Ala-Glu-Val-Asp, following Asp-163. The reduced expression of
endogenous ANP32B enhances caspase-3 activation and apoptosis
(12). In the current study, in
order to determine the functional significance of ANP32B cleavage,
leukemic U937T cell lines with inducible expression of ANP32B(wild
type; WT), the uncleavable mutant ANP32B(D163A) and the N-terminal
fragment ANP32B(1–163) were generated, and the effect of ANP32B
cleavage on leukemic cell apoptosis was investigated.
Materials and methods
Cell lines
The U937T cells provided by Dr Tenen from the
Harvard Institute of Medicine (Harvard Medical School, Boston, MA,
USA) (22) were U937 cells stably
transfected with a pUHD-tetracycline-responsive transcription
activator (tTA) under the control of a tetracycline-inducible
promoter. The U937T cells were cultured in RPMI-1640 medium
supplemented with 10% fetal bovine serum (FBS), 1 mg/ml
tetracycline and 0.5 mg/ml puromycin (Sigma-Aldrich, St. Louis, MO,
USA).
Establishment of U937T stable
transformant
Full-length (wild type and D163A mutant) and
fragment (1-163) of ANP32B cDNA with flag tag, a polypeptide
protein tag with sequence motif DYKXXD, were amplified from the
plasmid which was generated as previously described (12), and then cloned and inserted into
the pTRE2hyg expression vector (BD Clontech, Palo Alto, CA, USA), a
tetracycline-responsive expression vector, in order to form the
corresponding plasmids. The sequence of the cDNA insert of the
plasmid was confirmed by sequencing (Applied Biosystems 3730/3730xl
DNA analyzer, Sangon Biotechnology, Shanghai, China). Following
which, the plasmids were transfected into the U937T cells at
passage 5 (provided by Dr DG Tenen at Harvard Institutes of
Medicine, Harvard Medical School, Boston, MA, USA) containing
stably transfected pUHD-tTA (BD Clontech, Palo Alto, CA, USA),
whose expression is turned off in the presence of tetracycline
(Calbiochem, San Diego, CA). In principle, the expression of
tetracycline-responsive transcription activator (tTA) and target
proteins should be extremely low in tetracycline-containing medium.
In the absence of tetracycline, tTA activates its own promoter to
produce more tTA which in turn induces the expression of target
proteins (23). To generate the
corresponding stable transformants, 1×107 U937T cells
were washed in RPMI-1640 medium and resuspended in 0.2 ml of
Isceve’s modified Dulbecco’s medium (Sigma-Aldrich) without FBS. A
total of 20 μg plasmid in 20 ml double-distilled H2O was
transferred to an electroporation cuvette with a 0.4-cm gap
(Bio-Rad Laboratories, Hercules, CA, USA). Electroporation was
performed using a Gene Pulser II Electroporation system (Bio-Rad
Laboratories) at 170 V and 960 mF. The samples were then
transferred to complete RPMI-1640 medium. Subsequent to a 24-h
resting period, 1 mg/ml tetracycline, 0.5 mg/ml puromycin and 500
mg/ml hygromycin B (Clontech Laboratories, Inc.) were added and
cells were incubated at 37°C in 5% CO2. The positive
polyclonal population (pool) was identified based on the induction
of the corresponding protein expression following tetracycline
withdrawal, and were named U937ANP32B(WT),
U937ANP32B(D163A) and U937ANP32B(1-163). An
empty plasmid-transfected cell line called U937empty was
used as the control. All cells were maintained in RPMI-1640 medium
supplemented with 10% FBS, 1 mg/ml tetracycline, 0.5 mg/ml
puromycin and 0.5 mg/ml hygromycin B.
Apoptosis assay
To induce apoptosis, ~3×105 cells/ml were
initially seeded and cultured in tetracycline-free Isceve’s
modified Dulbecco’s medium for 10 days to induce the expression of
the corresponding proteins. The cells were then incubated with 1 μM
etoposide (Enzo Life Sciences, Inc., Farmingdale, NY, USA) for an
additional 12 h. The annexin-V assay was performed using a flow
cytometer (BD FACSCanto II; BD Biosciences, San Jose, CA, USA)
according to the manufacturer’s instructions for the ApoAlert
Annexin-V Apoptosis kit (Clontech Laboratories, Inc.), in order to
count the number of annexin-V-positive apoptotic cells. Caspase-3
activity was determined by the Caspase 3 Assay kit, Colorimetric
(Sigma-Aldrich).
Western blots
The protein lysates were equally loaded onto a
10–12% SDS-PAGE gel using a mini gel apparatus (Bio-Rad
Laboratories, Inc.) and subsequently transferred to a
nitrocellulose membrane (Bio-Rad Laboratories). The membranes were
blocked with 5% nonfat dry milk solution in tris-buffered saline
(Beyotime) for 1 h at room temperature and then incubated in
primary antibody dissolved in the 5% nonfat dry milk solution at
4°C overnight. The following antibodies were used: Polyclonal
rabbit anti-human protein kinase Cδ (PKCδ) (C-20; SC-937; Santa
Cruz Biotechnology, Inc., Santa Cruz, CA, USA); polyclonal rabbit
anti-human cleaved caspase-3 (9661; Cell Signaling Technology,
Danvers, MA, USA); monoclonal mouse anti-human poly[ADP (adenosine
diphosphate)-ribose] polymerase (PARP; F-2; SC-8007; Santa Cruz
Biotechnology, Inc.); polyclonal rabbit anti-human Bcl-2 (sc-492;
Santa Cruz Biotechnology, Inc.) and polyclonal rabbit anti-human
ANP32B (10843-1-AP; ProteinTech Group, Inc., Chicago, IL, USA),
with monoclonal mouse anti-β-actin mAb (EMD Millipore, Billerica,
MA, USA) to confirm equal loading. Following washing with PBS, the
blots were incubated with horseradish peroxidase-conjugated
secondary antibody (Dako, Glostrup, Denmark) corresponding to the
primary antibody in 5% nonfat dry milk solution for 1 h at room
temperature. Proteins were then detected using a luminol detection
reagent (Santa Cruz Biotechnology, Inc.), and developed onto X-ray
films (Kodak, Rochester, NY, USA).
Immunofluorescent staining of cells
Cells were collected onto slides using a Shandon
Cytospin 4 Cytocentrifuge (Thermo Fisher Scientific, Waltham, MA,
USA) at 800 × g for 5 min and fixed with 4% formaldehyde (Beyotime)
for 10 min. Subsequent to permeabilization with 0.3% Triton X-100
(Beyotime) in phosphate-buffered saline (PBS; Beyotime) for 10 min,
cells were incubated with 1% bovine serum albumin (Beyotime) in PBS
for 2 h at room temperature, followed by overnight incubation with
the ANP32B antibody. Cells were stained with fluorescein
isothiocyanate-labeled anti-rabbit IgG (Dako) for 1 h. Cell nuclei
were stained with 4′,6-diamidino-2-phenylindole (Molecular Probes
Life Technologies, Eugene, OR, USA). Fluorescent signals were
measured using an MRC-1024 laser scanning confocal microscope
equipped with a Zeiss ×60 objective (Bio-Rad Laboratories) or an
Olympus BX51 fluorescence microscope (100x/1.30 oil objective lens;
Olympus Corporation, Tokyo, Japan).
Statistical analysis
The differences between the two groups were analyzed
for statistical significance using the nonparametric Wilcoxon rank
sum test. P<0.05 was considered to indicate a statistically
significant difference.
Results
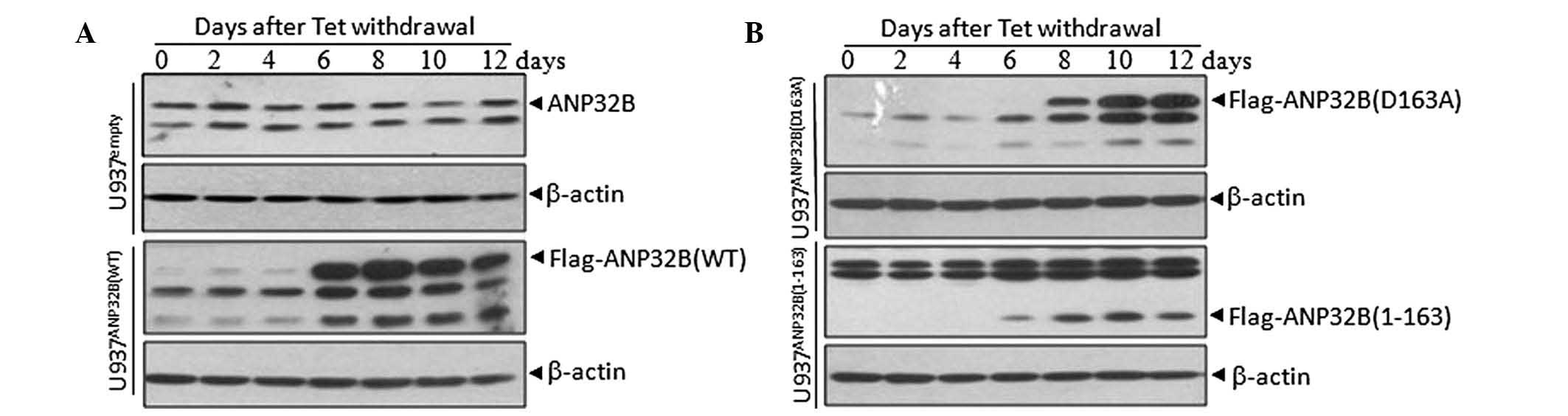
Establishment of cell lines with
inducible expression of ANP32B and its mutants
In previous studies, ANP32B has been demonstrated to
be a caspase-3 substrate during apoptotic induction, and is
primarily cleaved after Asp163 (12). To determine the functional
significances of ANP32B cleavage during apoptosis, leukemic U937T
cell lines with inducible expression of WT, D163A mutant and
N-terminal fragment (1-163) ANP32B, were generated by a Tet-Off
gene expression system. In the Tet-off system, gene expression is
turned on when tetracycline is removed from the culture medium.
U937Tempty, U937TANP32B(WT),
U937TANP32B(D163A) and U937TANP32B(1-163)
cells were all induced to express flag-tagged ANP32B or its
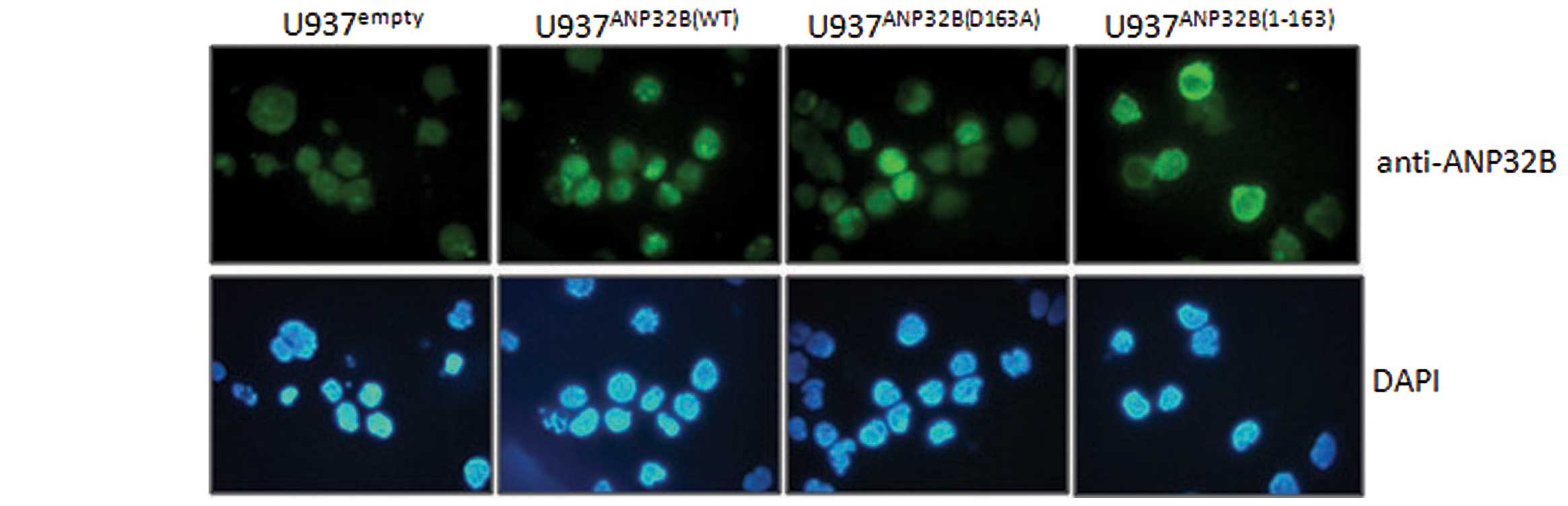
mutants, by tetracycline withdrawal for 6–12 days (Fig. 1). Immunofluorescent analysis with
the antibody against ANP32B indicated that ANP32B(WT) and
ANP32B(D163A) proteins were localized in the nuclei, while
ANP32B(1-163) was in the cytosol (Fig.
2). This was due to the absence of the nuclear localization
signal (NLS), which was mapped to amino acid residues KRKR at
position 239–242 in the C-terminal of this protein.
Uncleavable ANP32B(D163A) protein
amplifies apoptosis induction via caspase-3
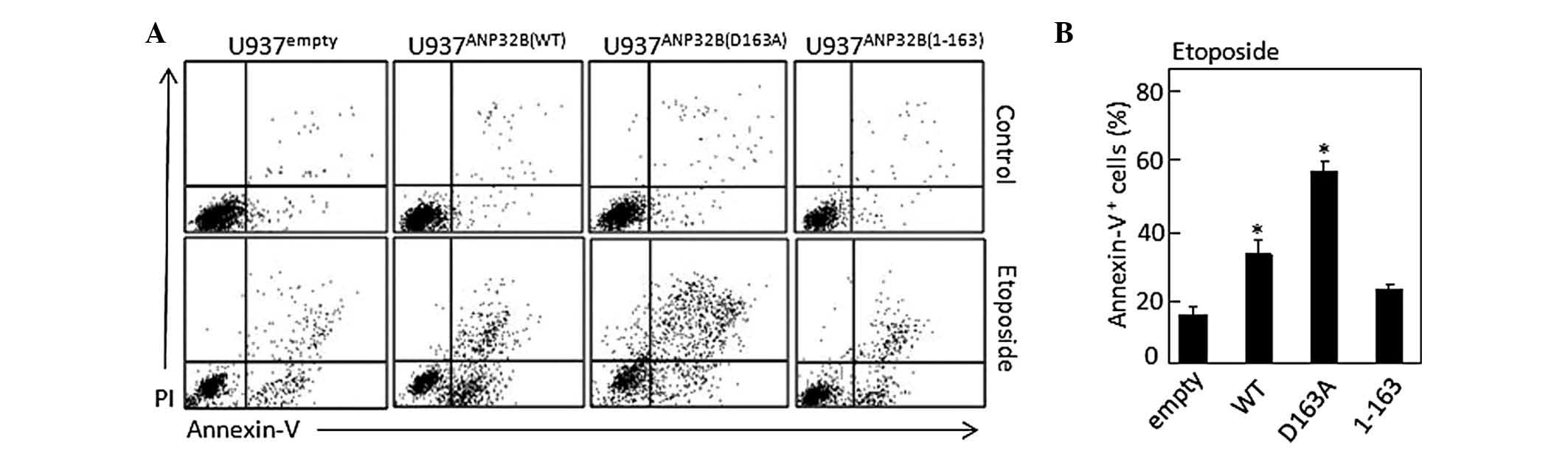
The contribution of caspase-3-mediated cleavage of
ANP32B to apoptosis was investigated by inducing the above cell
lines to undergo apoptosis and measuring the number of
annexin-V-positive cells using flow cytometry (Fig. 3A). The results demonstrated that
compared with U937Tempty cells (18.3±1.9%), apoptosis
was significantly increased in U937TANP32B(WT) cells
(37.2±2.7%; P<0.05) as well as highly significantly enhanced in
U937TANP32B(D163A) cells (58.5±2.1%; P<0.01), but was
not markedly altered in U937TANP32B(1-163) cells
(21.8±0.8%) (Fig. 3B). This
suggested that the uncleavable ANP32B(D163A) possesses
pro-apoptotic activity.
Uncleavable ANP32B(D163A) protein
enhances PKCδ and caspase-3 activity
The catalytic activity of PKCδ is established to be
important at the early stage of etoposide-induced apoptosis (prior
to caspase-3 activation) (24,25),
while activated caspase-3 produces an amplifying effect on PKCδ
activation (26). To further
confirm the roles of different ANP32B forms in apoptosis, the
proteolytic activation of PKCδ and caspase-3 was investigated, in
addition to the cleavage of PARP, a common substrate of activated
caspase-3 during apoptosis. As depicted in Fig. 4A, treatment with 1 μM etoposide
induced slight fragmentation of PKCδ, caspase-3 and PARP in
U937Tempty cells, while these fragmentations were
moderately and significantly enhanced in U937TANP32B(WT)
and U937TANP32B(D163A) cells, respectively, but remained
unchanged in U937TANP32B(1-163) cells. Caspase-3
activation was observed to be regulated similarly (Fig. 4B).
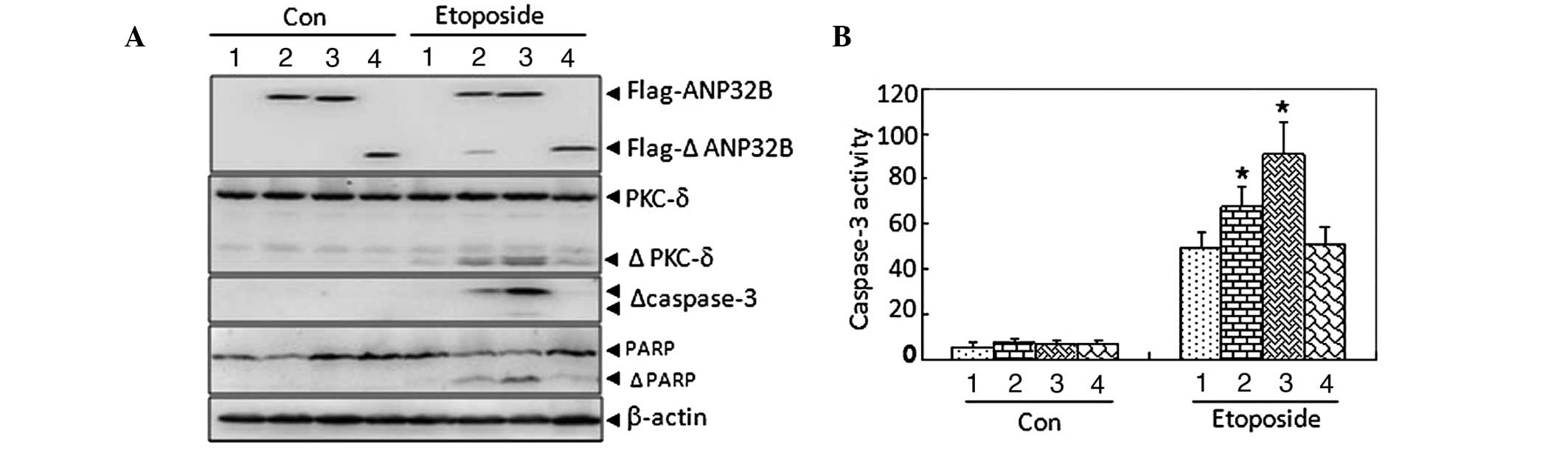 | Figure 4Effect of uncleavable ANP32B(D163A)
protein on etoposide-induced proteolytic cleavage of PKCδ and
caspase-3 activation. (A) Western blot analysis with β-actin as a
loading control. (B) Analysis of caspase-3 activity.
*P<0.05 vs. U937Tempty cells with the
corresponding treatment in an independent experiment with
triplicates. ANP32B, acidic leucine-rich nuclear phosphoprotein
32B; PKC-δ, protein kinase Cδ; WT, wild type; 1,
U937Tempty; 2, U937TANP32B(WT); 3,
U937TANP32B(D163A); 4, U937TANP32B(1-163);
PARP, poly[ADP (adenosine diphosphate)-ribose] polymerase; Con,
control; Δ, fragments of the corresponding protein, |
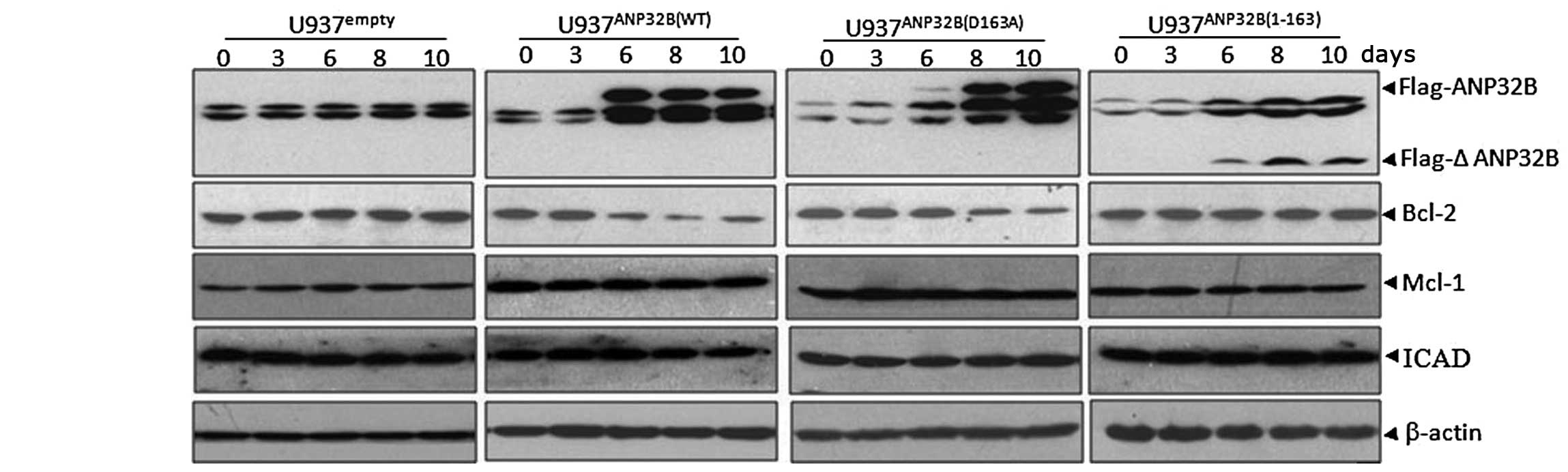
Inducible expression of uncleavable
ANP32B(D163A) protein reduces Bcl-2 protein levels
ANP32B has been reported to regulate gene
transcription as a histone chaperone in the nucleus (27), thus, the mechanisms underlying the
pro-apoptotic role of full-length ANP32B were investigated, via the
detection of several key apoptosis-associated genes, including
Bcl-2, Mcl-1 and inhibitor of caspase-activated DNase (ICAD) in the
inducible cell lines. The data indicated that the expression levels
of anti-apoptotic Bcl-2 protein were progressively reduced
subsequent to tetracycline removal, in parallel with the induction
of full-length ANP32B protein in U937TANP32B(WT) and
U937TANP32B(D163A) cells, while Mcl-1 and ICAD
expression remained unchanged (Fig.
5). In addition, the expression of Bcl-2 was not affected by
ANP32B(1-163), which was localized in the cytosol and had no effect
on apoptosis induction. The reduced expression of Bcl-2 protein is
hypothesized to contribute to the apoptosis-enhancing ability of
full-length ANP32B.
Discussion
Based on previous subcellular proteomic analysis
(28), ANP32B was identified as a
novel caspase-3 substrate primarily cleaved at the sequence of
Ala-Glu-Val-Asp, following Asp-163 (12). In order to ascertain whether the
cleavage of ANP32B contributes to apoptosis induction in leukemic
cells, leukemic U937T cell lines with inducible expression of
ANP32B(WT) along with two mutants, the D163A mutant and its
N-terminal fragment (1-163) were generated, which were treated with
etoposide following tetracycline withdrawal. Notably,
overexpression of either ANP32B(WT) or its uncleavable mutant
[ANP32B(D163A)] was observed to accelerate etoposide-induced
apoptosis and caspase-3 activation, whereas enforced expression of
the N-terminal fragment [ANP32B(1-163)] had no effect, suggesting
the LCAR of ANP32B may be critical for the apoptosis-inducing
activity of this protein. A previous study that supports this
theory demonstrated that ANP32A LCAR is able to directly promote
caspase-3 activation (29). In
addition, the cellular localization of ANP32B(1-163) lacking LCAR
was different from ANP32B(WT) and ANP32B(D163A). Consistent with
the study by Mutai et al (30), the current study demonstrated that
full length ANP32B protein (WT and D163A mutant) accumulated in the
nucleus, whereas the N-terminal fragment of ANP32B was localized in
the cytoplasm, due to the absence of the NLS. This may be the
primary cause of the varied apoptosis-inducing abilities of the
three transformants.
An anti-apoptotic function of ANP32B has been
previously reported, indicating that siRNA-mediated depletion of
ANP32B enhanced NSC606985- and etoposide-induced apoptosis
(12). However, the current study
observed that overexpression of ANP32B(WT) also enhanced
etoposide-induced apoptosis and the uncleavable ANP32B(D163A)
mutant significantly accelerated it, suggesting a pro-apoptotic
function of ANP32B. However, the explanation of this inconsistency
remains unclear. It is hypothesized that ANP32B may have
bidirectional roles in apoptotic induction. In support of this
theory, nuclear factor-κB has been reported to exhibit
anti-apoptotic and pro-apoptotic activity, via the induction of
different anti-apoptotic and pro-apoptotic genes (31). From the observation that ANP32B was
reported to regulate transcription factor activity as a histone
chaperone, it may be hypothesized that the bidirectional roles of
ANP32B potentially result from its regulation of certain
transcription factors with bidirectional roles in apoptosis
induction.
To further investigate the mechanism of the
pro-apoptotic activity of full-length ANP32B, the expression of
several apoptosis-associated genes were analyzed in these inducible
cell lines. As demonstrated in Fig.
5, the expression of ANP32B(WT) and its uncleavable mutant
[ANP32B(D163A)] resulted in progressively reduced expression of the
anti-apoptotic Bcl-2 protein, suggesting that ANP32B may exert its
pro-apoptotic role via the inhibition of Bcl-2 expression. However,
the mechanism of regulation of the Bcl-2 protein remains unclear
and requires further investigation.
Acknowledgements
The current study was supported by the Ministry of
Science and Technology (grant no. 2013CB910903), the Shanghai
Science and Technology Committee (grant no. 11ZR1421500) and the
National Natural Science Foundation of China (grant nos. 30971276,
31170783 and 31300679).
References
|
1
|
Fuchs Y and Steller H: Programmed cell
death in animal development and disease. Cell. 147:742–758. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Thornberry NA and Lazebnik Y: Caspases:
enemies within. Science. 281:1312–1316. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Utz PJ and Anderson P: Life and death
decisions: regulation of apoptosis by proteolysis of signaling
molecules. Cell Death Differ. 7:589–602. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Johnson CE and Kornbluth S: Caspase
cleavage is not for everyone. Cell. 134:720–721. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Timmer JC and Salvesen GS: Caspase
substrates. Cell Death Differ. 14:66–72. 2007. View Article : Google Scholar
|
|
6
|
Matilla A and Radrizzani M: The Anp32
family of proteins containing leucine-rich repeats. Cerebellum.
4:7–18. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Adegbola O and Pasternack GR:
Phosphorylated retinoblastoma protein complexes with pp32 and
inhibits pp32-mediated apoptosis. J Biol Chem. 280:15497–15502.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hoffarth S, Zitzer A, Wiewrodt R, et al:
pp32/PHAPI determines the apoptosis response of non-small-cell lung
cancer. Cell Death Differ. 15:161–170. 2008. View Article : Google Scholar
|
|
9
|
Jiang X, Kim HE, Shu H, et al: Distinctive
roles of PHAP proteins and prothymosin-alpha in a death regulatory
pathway. Science. 299:223–226. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Pan W, da Graca LS, Shao Y, et al:
PHAPI/pp32 suppresses tumorigenesis by stimulating apoptosis. J
Biol Chem. 284:6946–6954. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Schafer ZT, Parrish AB, Wright KM, et al:
Enhanced sensitivity to cytochrome c-induced apoptosis mediated by
PHAPI in breast cancer cells. Cancer Res. 66:2210–2218. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Shen SM, Yu Y, Wu YL, et al:
Downregulation of ANP32B, a novel substrate of caspase-3, enhances
caspase-3 activation and apoptosis induction in myeloid leukemic
cells. Carcinogenesis. 31:419–426. 2010. View Article : Google Scholar
|
|
13
|
Brody JR, Kadkol SS, Hauer MC, et al: pp32
reduction induces differentiation of TSU-Pr1 cells. Am J Pathol.
164:273–283. 2004. View Article : Google Scholar
|
|
14
|
Kular RK, Cvetanovic M, Silferd S, et al:
Neuronal differentiation is regulated by leucine-rich acidic
nuclear protein (LANP), a member of the inhibitor of histone
acetyltransferase complex. J Biol Chem. 284:7783–7792. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Puente LG, Carrière JF, Kelly JF and
Megeney LA: Comparative analysis of phosphoprotein-enriched myocyte
proteomes reveals widespread alterations during differentiation.
FEBS Lett. 574:138–144. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yu Y, Shen SM, Zhang FF, et al: Acidic
leucine-rich nuclear phosphoprotein 32 family member B (ANP32B)
contributes to retinoic acid-induced differentiation of leukemic
cells. Biochem Biophys Res Commun. 423:721–725. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Amasaki H, Ogawa M, Nagasao J, et al:
Distributional changes of BrdU, PCNA, E2F1 and PAL31 molecules in
developing murine palatal rugae. Ann Anat. 185:517–523. 2003.
View Article : Google Scholar
|
|
18
|
Sun W, Hattori N, Mutai H, et al: PAL31, a
nuclear protein required for progression to the S phase. Biochem
Biophys Res Commun. 280:1048–1054. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Fukukawa C, Tanuma N, Okada T, et al:
pp32/ I-1(PP2A) negatively regulates the Raf-1/MEK/ERK pathway.
Cancer Lett. 226:155–160. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Opal P, Garcia JJ, McCall AE, et al:
Generation and characterization of LANP/pp32 null mice. Mol Cell
Biol. 24:3140–3149. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Reilly PT, Afzal S, Gorrini C, et al:
Acidic nuclear phosphoprotein 32kDa (ANP32)B-deficient mouse
reveals a hierarchy of ANP32 importance in mammalian development.
Proc Natl Acad Sci USA. 108:10243–10248. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Boer J, Bonten-Surtel J and Grosveld G:
Overexpression of the nucleoporin CAN/NUP214 induces growth arrest,
nucleocytoplasmic transport defects, and apoptosis. Mol Cell Biol.
18:1236–1247. 1998.PubMed/NCBI
|
|
23
|
Boer J, Bonten-Surtel J and Grosveld G:
Overexpression of the nucleoporin CAN/NUP214 induces growth arrest,
nucleocytoplasmic transport defects, and apoptosis. Mol Cell Biol.
18:1236–1247. 1998.PubMed/NCBI
|
|
24
|
Matassa AA, Carpenter L, Biden TJ, et al:
PKCdelta is required for mitochondrial-dependent apoptosis in
salivary epithelial cells. J Biol Chem. 276:29719–29728. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Qi X and Mochly-Rosen D: The PKCdelta -Abl
complex communicates ER stress to the mitochondria - an essential
step in subsequent apoptosis. J Cell Sci. 121:804–813. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Song MG, Gao SM, Du KM, et al: Nanomolar
concentration of NSC606985, a camptothecin analog, induces
leukemic-cell apoptosis through protein kinase Cdelta-dependent
mechanisms. Blood. 105:3714–3721. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Munemasa Y, Suzuki T, Aizawa K, et al:
Promoter region-specific histone incorporation by the novel histone
chaperone ANP32B and DNA-binding factor KLF5. Mol Cell Biol.
28:1171–1181. 2008. View Article : Google Scholar :
|
|
28
|
Yu Y, Wang LS, Shen SM, et al: Subcellular
proteome analysis of camptothecin analogue NSC606985-treated acute
myeloid leukemic cells. J Proteome Res. 6:3808–3818. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hill MM, Adrain C, Duriez PJ, et al:
Analysis of the composition, assembly kinetics and activity of
native Apaf-1 apoptosomes. EMBO J. 23:2134–2145. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Mutai H, Toyoshima Y, Sun W, et al: PAL31,
a novel nuclear protein, expressed in the developing brain. Biochem
Biophys Res Commun. 274:427–433. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Karin M and Lin A: NF-kappaB at the
crossroads of life and death. Nat Immunol. 3:221–227. 2002.
View Article : Google Scholar : PubMed/NCBI
|