Introduction
Atherosclerosis is a chronic inflammatory and
fibroproliferative disease. Effective results have not been
achieved by controlling and intervening with the associated risk
factors, including smoking, dyslipidemia, hypertension and diabetes
(1–3). Therefore, these risk factors may not
fully explain the occurrence and development of atherosclerosis and
other targets of atherosclerosis require identification.
MicroRNAs (miRNAs) are small (~23 nt), non-coding
RNAs that regulate gene expression at the post-transcriptional
level. MiRNAs have been shown to have an important role in the
development and progression of atherosclerosis (4–6).
They are expressed in a tissue-specific manner and are associated
with cell proliferation, apoptosis and differentiation (7,8).
MiRNAs have been previously implicated in atherosclerotic plaque
formation, caused by hyperlipidemia and hypertension (9,10).
MiRNAs have also been directly associated with anti-atherosclerotic
signals in vascular smooth muscle and endothelial cells (11). The exact mechanisms of the role of
miRNAs in atherosclerosis remain to be elucidated.
MiR-142-5p is a member of the miR-142 miRNA family
which have known roles in cancer, immune diseases and embryonic
stem cells (12,13). However, the expression of
miR-142-5p in atherosclerotic plaque and its roles in
atherosclerosis are currently unclear.
In the present study, the expression levels of
miR-142-5p were detected in murine atherosclerotic plaques and
human macrophages. The present study also aimed to identify
miR-142-5p target genes, and its effects on apoptosis in
macrophages.
Materials and methods
Materials
The following reagents, kits, primers and cells were
used and sourced from the following companies: Anti-rabbit
transforming growth factor-β2 (TGF-β2) monoclonal antibody
(ProteinTech Group, Inc., Chicago, IL, USA); anti-mouse GAPDH
monoclonal antibody (SunShineBio, Nanjing, China); miR-142-5p
primers (Exiqon Co., Copenhagen, Denmark); TGF-β2 primers (ShangDe
Biomedical Engineering, Shanghai, China); total RNA extraction kit
(TRIzol®; Invitrogen Life Technologies, Carlsbad, CA,
USA); THP-1 human monocytes, smooth muscle and endothelial cells
(American Type Culture Collection, Manassas, VA, USA). Other
reagents used throughout the present study were obtained from the
Key Laboratory of Cardiovascular Remodeling and Function Research,
Qilu Hospital, Shandong University, (Jinan, China).
Animals
Apolipoprotein E−/− (apoE−/−) mice were purchased
from WeiTong LiHua Co. (Beijing, China). The animal experiments
were approved by the Institutional Animal Care and Use Committee of
Shandong University (Jinan, China).
Animal model of atherosclerosis
The eight-week-old male apoE−/− mice were fed a
high-fat diet, that consisted of a standard diet plus 2%
cholesterol and 5% lard oil, for two weeks, following a three day
standard diet. The mice were then intraperitoneally injected with
0.08% sodium pentobarbital (40 mg/kg; Beijing OuHe Technology Co.,
Ltd., Beijing, China) and underwent surgery. Carotid
atherosclerotic plaques were induced in the mice using perivascular
constrictive collars, which were placed on the right common carotid
artery, as described by previous methods (14). The mice were divided into three
groups (n= 12/group): Control, stable plaque, and vulnerable
plaque. The mice then received a high-fat diet for a further 12
weeks. The vulnerable plaque group underwent Pisa syndrome noise
interference for four weeks during the 12 week period. In brief,
experimental mice were kept in a 50 ml plastic pipe with a covered
end and a through hole and then subjected to 110 dB noise
stimulation intensity for 30 sec (Beijing Great Wall Radio Factory,
Beijing, China), every 5 min for 6 h/day. All of the mice were
sacrificed by cervical dislocation and blood samples (1 ml) were
obtained from the abdominal vein and stored at −80°C, until further
use. Sections of the carotid arteries were cut in optimal cutting
temperature compound medium, and stored in liquid nitrogen, until
further use.
Gene microarray analysis for miR-142-5p
expression
The isolated carotid artery sections were removed
from liquid nitrogen and the vascular peripheral tissue was placed
on ice. The expression levels of miR-142-5p, in the atherosclerotic
plaques, were determined using a miRNA microarray assay with rat
miRNA array probes (Kangchen Bio-tech Inc., Shanghai, China).
Cell culture and transfection
Primary human endothelial cells and human
macrophages were obtained from the American Type Culture Collection
(Manassas, VA, USA) and were cultured in RPMI-1640 medium (Gibco
Life Technologies, Carlsbad, CA, USA) supplemented with 10% fetal
bovine serum (FBS; Beijing Solarbio Science & Technology Co.,
Ltd., Beijing, China) at 37°C in a 5% CO2 atmosphere.
Human smooth muscle cells were cultured in a medium of 2% FBS, 1%
smooth muscle cell growth supplement and 1% penicillin/streptomycin
(Xiang Bo Biological Technology Co., Guangzhou, China) under
identical conditions. The media were refreshed every 2–3 days. The
cells from passages 3–5 were used for the following experiments.
Following culture, cell aliquots were transferred to freezing tubes
with cell freezing medium and stored at −80°C overnight and then
preserved in a liquid nitrogen tank until further used. Cell were
seeded onto six-well plates, when cell confluence reached 70% 5 μl
diethylpyrocarbonate (1:10 dilution; Beijing Solarbio Science &
Technology Co., Ltd.,) of mir-142-5p inhibitor (Shanghai GenePharma
Co., Ltd., Shanghai, China) was added to 250 μl Opti-MEM (Gibco
Life Technologies) culture medium and incubated at room temperature
for 5 min. Subsequently, 2.5 μl Lipofectamine 2000 (Shanghai Yijie
Biotechnology Co., Ltd., Shanghai, China) was added to 250 μl
Opti-MEM culture medium and incubated at room temperature for 5
min. The liposome suspension was then added to the mir-142-5p
inhibitor liquid and incubated at room temperature for 15 min. The
suspension was then added to the cells and incubated for 6 h, the
medium was then replaced and the cells were cultured for a further
48 h. To study the miR-142-5p expression levels, the cells were
divided into two treatment groups: Control and oxidized low-density
lipoprotein treated (ox-LDL; 90 μl, 50 mg/ml, for 24 h). To study
TGF-β2 expression levels in the macrophages, the cells were divided
into five groups: Control, control + ox-LDL, miR-142-5p inhibitor
transfection + ox-LDL, miR-142-5p mimic transfection, and negative
control (NC) + ox-LDL. To investigate the effects of miR-142-5p on
the rate of apoptosis of human macrophages, the cells were divided
into seven groups: Control, control + ox-LDL, miR-142-5p inhibitor
transfection + ox-LDL, TGF-β2 inhibitor transfection + miR-142 -5p
inhibitor + ox-LDL, miR-142-5p mimic transfection, TGF-β2 inhibitor
+ ox-LDL, N.C + ox-LDL.
Quantitative polymerase chain
reaction
Total RNA was extracted from the human endothelial
cells, smooth muscle cells and macrophages using
TRIzol®. A total of 1 μg RNA, from each group, was
reverse transcribed using the PrimeScript RT Reagent kit (Takara
Bio Inc., Otsu, Japan), and the qPCR was performed using the
Bio-Rad IQ5 Real-Time PCR Detection system (Bio-Rad Laboratories
Inc., Hercules, CA, USA). All reagents used for qPCR were from this
detection system unless otherwise stated. The reaction system
consisted of 10 μl SYRB Green Mix (Takara Bio, Inc.), 2 μl miRNA
primer mix and 8 μl diluted cDNA. The following primer sequences
were used: TGF-β2 forward, 5′-ACAAAATAGACATGCCGCCC-3′, and reverse,
5′-GATGGCATCAAGGTACCCACAG-3′; Hsa-miR-142-5p forward,
5′-AACTCCAGCTGGTCCTTAG-3′, and reverse, 5′-TCTTGAACCCTCATCCTGT-3′;
Hsa-miR-142-5p inhibitor were, 5′-AGUAGUGCUUUCUACUUUAUG-3′;
Hsa-miR-142-5p mimic forward, 5′-CAUAAAGUAGAAAGCACUACU-3′, and
reverse, 5′-UAGUGCUUUCUACUUUAUGUU-3′. The housekeeping genes U6 or
β-actin were used as internal controls. Primer sequences were as
follows: U6 forward, 5′-CTCGCTTCGGCAGAC-3′ and reverse,
5′-AACGCTTACGAATTT -3′; β-actin forward,
5′-CGTGCGTGACATTAAGGAGA′-3′ and reverse,
5′-CACCTTCACCGTTCCAGTTT-3′. The cycling conditions were as follows:
Initial denaturation at 95°C for 10 min followed by 40 cycles of
95°C for 10 sec, 56°C for 10 sec and 72°C for 30 sec. qPCR
concluded with 65°C for 30 sec and 70°C for 30 sec. Changes in the
gene expression levels were calculated using the cycle threshold
(Ct) comparison method, by the formula 2−ΔΔCt.
Western blotting
The cell lysates from the treated macrophages were
prepared, as described previously (15). The Bio-Rad Protein Assay Reagent
kit (Bio-Rad Laboratories Inc.) was used to measure protein
concentrations. The protein samples (20 μg) were separated using
10% SDS-PAGE, at 90 V for 1 h and transferred electrophoretically
to polyvinylidene fluoride membranes (Millipore, Bellerica, MA,
USA), at 110 mA for 0.5 h. The membranes were then blocked with 5%
milk for 2 h at room temperature, and incubated with the primary
antibodies at 4°C overnight. The membranes were washed three times
with tris-buffered saline containing Tween® (10
min/wash), and then incubated with a secondary horseradish
peroxidase-labeled antibody at room temperature for 1.5 h. The
signals were visualized using an Enhanced Chemiluminescence
substrate (GE Healthcare Life Sciences, Chalfont, UK).
Apoptosis detection
The number of apoptotic human macrophages was
quantified using the Annexin V-PE Apoptosis Detection kit (Beyotime
Institute of Biotechnology, Hainen, China). The apoptotic cells
were calculated as number of apoptotic cells/total cell number ×
100%.
Statistical analysis
The data analysis was carried out using SPSS version
17.0 (SPSS Inc., Chicago, IL, USA). The data are presented as the
means ± standard error of the mean. Statistical comparisons were
performed using a paired student’s t test and an analysis of
variance. A P<0.05 was considered to indicate a statistically
significant difference.
Results
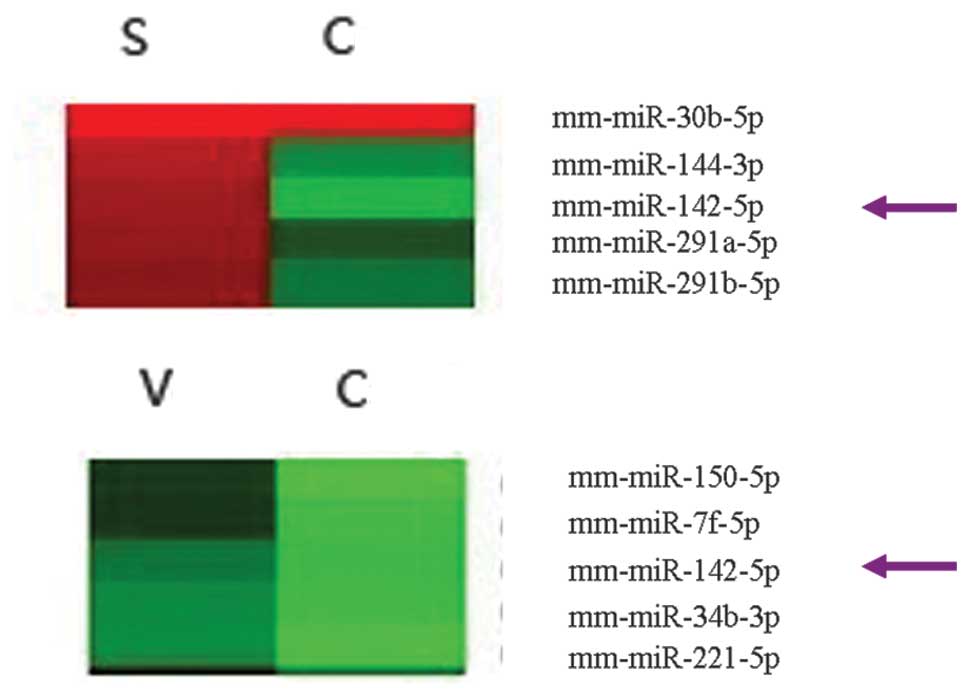
Expression levels of miR-142-5p are
upregulated in the atherosclerotic plaques of mice
The expression levels of miR-142-5p were 6.84-fold
higher in the mice with stable plaques, and was 2.69-fold higher in
the mice with vulnerable plaques, as compared with the controls
(Fig. 1).
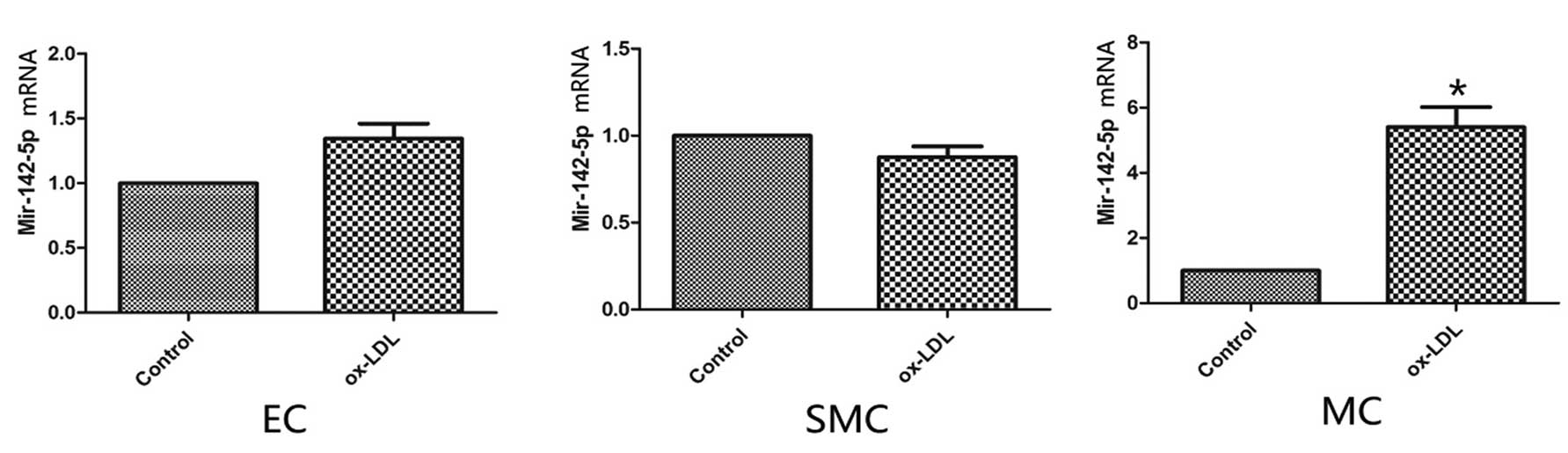
Expression levels of miR-142-5p are
upregulated in human macrophages treated with ox-LDL
The expression levels of miR-142-5p in the human
macrophages treated with ox-LDL were upregulated, as compared with
the control macrophages (P<0.05). However, there were no marked
differences from the controls in either the endothelial or smooth
muscle cells (Fig. 2).
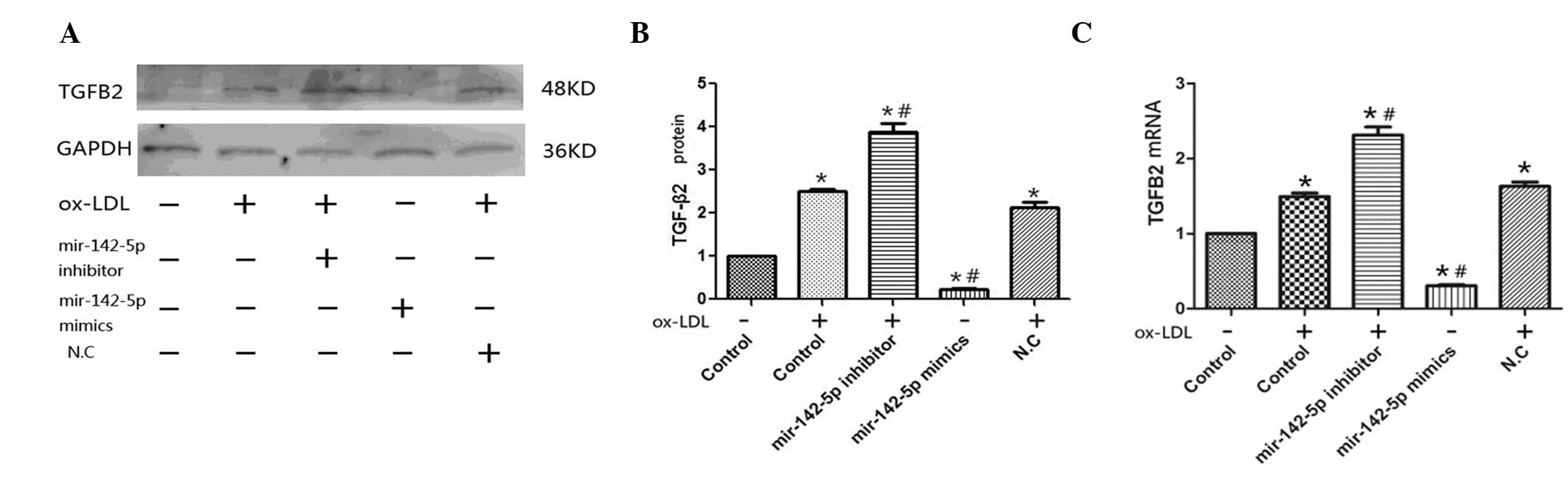
TGF-β2 is predicted to be a target gene
of miR-142-5p
miRanda (www.microrna.org/microrna/home.do) target-gene
prediction software was used to predict the target gene of
miR-142-5p, and TGF-β2 was predicted to be the most probable target
gene. To verify whether TGF-β2 was a target gene, a miR-142-5p
inhibitor and mimic were transfected into macrophages. The
expression levels of TGF-β2 were higher in the cells transfected
with the miR-142-5p inhibitor and treated with ox-LDL, as compared
with the cells undergoing ox-LDL treatment alone (P<0.05;
Fig. 4), and were the lowest when
the cells were transfected with the miR-142-5p mimic (P<0.05).
These results suggest that TGF-β2 may be a target gene of
miR-142-5p.
Discussion
Tissue-specific expression is an important
characteristic of miRNA expression (16). The present study demonstrated that
miR-142-5p expression was upregulated in atherosclerotic plaques
obtained from apoE−/− mice. In addition, miR-142-5p was shown to be
associated with the apoptosis of macrophages, through the
regulation of its predicted target gene, TGF-β2.
MiRNAs are small, non-coding, highly conserved RNAs
that regulate gene expression at the posttranscriptional level
(17–19). MiRNAs may negatively regulate gene
expression either by promoting the decomposition of mRNAs, or
inhibiting the translation of protein (20,21).
MiRNAs have been shown to have an important role in cardiovascular
diseases, including atherosclerosis (22–24),
and can regulate the functions of endothelial cells, macrophages
and vascular smooth muscle cells (25–28).
They have previously been demonstrated to modulate every stage of
atherosclerosis, by different stimuli (29–31).
MiR-142-5p is a member of the miR-142 family, which
is involved in the pathogenesis of various diseases (13,32,33).
Previous research into miR-142-5p has mainly focused on its
associations with tumors, immune diseases and stem cells (32); however, its role in atherosclerosis
remains unknown. In the present study, an atherosclerotic plaque
apoE−/− mouse model was generated and the expression levels of
miR-142-5p were upregulated in the atherosclerotic plaques of the
apoE−/− mice.
Atherosclerosis is a chronic non-resolving
inflammatory disease. Monocytes/macrophages are major immune cells,
which are thought to be responsible for the development of
atherosclerosis (34–36). In the present study, significant
miR-142-5p expression was detected in macrophages, but not
endothelial or smooth muscle cells. Apoptosis of macrophages has
been shown to contribute to both early and advanced atherosclerosis
(37,38). The accumulation of apoptotic
macrophages leads to secondary necrosis, necrotic core enlargement
and plaque instability (39,40).
Furthermore, macrophages have been shown to be involved in cell
apoptosis in atherosclerotic plaques, through targeting specific
control genes (40,42). The present study determined that
apoptosis of macrophages could be affected by miR-142-5p.
MiRNAs negatively regulate the expression of target
genes. A database-based target gene prediction software predicted
that TGF-β2 was the most probable target gene of miR-142-5p. To
verify whether TGF-β2 was the target of miR-142-5p, an inhibitor
and a mimic of miR-142-5p were transfected into macrophages, and
the effects were observed on TGF-β2 protein and mRNA expression
levels. The results verified that TGF-β2 was the likely target gene
of miR-142-5p. TGF-β2 is a cytokine associated with a variety of
functions, it has previously been shown to participate in cell
proliferation, apoptosis and differentiation (42–45).
It has an important role in the pathophysiological processes of
tissue repair, inflammation, arterial atherosclerosis and cancer
(45–48). In the present study, miR-142-5p was
shown to be associated with the apoptosis of macrophages by
negatively regulating TGF-β2. In conclusion, miR-142-5p was shown
to be involved in atherosclerosis in mice, and TGF-β2 was
identified as its target. MiR-142-5p was shown to regulate
macrophage apoptosis by targeting TGF-β2. The present study
provides a novel target for further study of atherosclerosis.
Acknowledgements
The present study was supported by the National
Basic Research Program of China (973 Program; nos. 2010CB732605 and
2011CB503906), the HI-TECH Technique and Development Program of
China (863 Program, no. 2007AA02Z448), the National Natural Science
Foundation of China (nos. 81270404 and 30970709), Science Program
of Shandong Province (no. 2006GG2202039).
References
|
1
|
Rodrigues AN, Abreu GR, Resende RS,
Goncalves WL and Gouvea SA: Cardiovascular risk factor
investigation: a pediatric issue. Int J Gen Med. 6:57–66. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Cesarino EJ, Vituzzo AL, Sampaio JM,
Ferreira DA, Pires HA and de Souza L: Assessment of cardiovascular
risk of patients with arterial hypertension of a public health
unit. Einstein (Sao Paulo). 10:33–38. 2012. View Article : Google Scholar
|
|
3
|
Pollex RL, Spence JD, House AA, et al: A
comparison of ultrasound measurements to assess carotid
atherosclerosis development in subjects with and without type 2
diabetes. Cardiovasc Ultrasound. 3:152005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Small EM and Olson EN: Pervasive roles of
microRNAs in cardiovascular biology. Nature. 469:336–342. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chang TC and Mendell JT: microRNAs in
vertebrate physiology and human disease. Annu Rev Genomics Hum
Genet. 8:215–239. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Mendell JT and Olson EN: MicroRNAs in
stress signaling and human disease. Cell. 148:1172–1187. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
He L and Hannon GJ: MicroRNAs: small RNAs
with a big role in gene regulation. Nat Rev Genet. 5:522–531. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Amelio I, Lena AM, Viticchie G, et al:
miR-24 triggers epidermal differentiation by controlling actin
adhesion and cell migration. J Cell Biol. 199:347–363. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Donners MM, Wolfs IM, Stöger LJ, et al:
Hematopoietic miR155 deficiency enhances atherosclerosis and
decreases plaque stability in hyperlipidemic mice. PLoS One.
7:e358772012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Nossent AY, Hansen JL, Doggen C, Quax PH,
Sheikh SP and Rosendaal FR: SNPs in microRNA binding sites in
3′-UTRs of RAAS genes influence arterial blood pressure and risk of
myocardial infarction. Am J Hypertens. 24:999–1006. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hergenreider E, Heydt S, Tréguer K, et al:
Atheroprotective communication between endothelial cells and smooth
muscle cells through miRNAs. Nat Cell Biol. 14:249–256. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kwanhian W, Lenze D, Alles J, et al:
MicroRNA-142 is mutated in about 20% of diffuse large B-cell
lymphoma. Cancer Med. 1:141–155. 2012. View
Article : Google Scholar
|
|
13
|
Saito Y, Suzuki H, Tsugawa H, Imaeda H,
Matsuzaki J, Hirata K, et al: Overexpression of miR-142-5p and
miR-155 in gastric mucosa-associated lymphoid tissue (MALT)
lymphoma resistant to Helicobacter pylori eradication. PLoS One.
7:e473962012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
von der Thüsen JH, van Berkel TJ and
Biessen EA: Induction of rapid atherogenesis by perivascular
carotid collar placement in apolipoprotein E-deficient and
low-density lipoprotein receptor-deficient mice. Circulation.
103:1164–1170. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang XL, Zhang L, Youker K, et al: Free
fatty acids inhibit insulin signaling-stimulated endothelial nitric
oxide synthase activation through upregulating PTEN or inhibiting
Akt kinase. Diabetes. 55:2301–2310. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lagos-Quintana M, Rauhut R, Yalcin A,
Meyer J, Lendeckel W and Tuschl T: Identification of
tissue-specific microRNAs from mouse. Curr Biol. 12:735–739. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Guo H, Ingolia NT, Weissman JS and Bartel
DP: Mammalian microRNAs predominantly act to decrease target mRNA
levels. Nature. 466:835–840. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Baek D, Villén J, Shin C, Camargo FD, Gygi
SP and Bartel DP: The impact of microRNAs on protein output.
Nature. 455:64–71. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Pasquinelli AE, Hunter S and Bracht J:
MicroRNAs: a developing story. Curr Opin Genet Dev. 15:200–205.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Bartel DP: MicroRNAs: genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Farh KK, Grimson A, Jan C, et al: The
widespread impact of mammalian MicroRNAs on mRNA repression and
evolution. Science. 310:1817–1821. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Nazari-Jahantigh M, Wei Y, Noels H, et al:
MicroRNA-155 promotes atherosclerosis by repressing Bcl6 in
macrophages. J Clin Invest. 122:4190–4202. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sun X, Zhang M, Sanagawa A, et al:
Circulating microRNA-126 in patients with coronary artery disease:
correlation with LDL cholesterol. Thromb J. 10:162012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang E and Wu Y: MicroRNAs: important
modulators of oxLDL-mediated signaling in atherosclerosis. J
Atheroscler Thromb. 20:215–227. 2013. View Article : Google Scholar
|
|
25
|
Xie C, Huang H, Sun X, et al: MicroRNA-1
regulates smooth muscle cell differentiation by repressing
Kruppel-like factor 4. Stem Cells Dev. 20:205–210. 2011. View Article : Google Scholar :
|
|
26
|
Tréguer K, Heinrich EM, Ohtani K, Bonauer
A and Dimmeler S: Role of the microRNA-17-92 cluster in the
endothelial differentiation of stem cells. J Vasc Res. 49:447–460.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chan YC, Roy S, Khanna S and Sen CK:
Downregulation of endothelial microRNA-200b supports cutaneous
wound angiogenesis by desilencing GATA binding protein 2 and
vascular endothelial growth factor receptor 2. Arterioscler Thromb
Vasc Biol. 32:1372–1382. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Tserel L, Runnel T, Kisand K, et al:
MicroRNA expression profiles of human blood monocyte-derived
dendritic cells and macrophages reveal miR-511 as putative positive
regulator of Toll-like receptor 4. J Biol Chem. 286:26487–26495.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Urbich C, Kuehbacher A and Dimmeler S:
Role of microRNAs in vascular diseases, inflammation, and
angiogenesis. Cardiovasc Res. 79:581–588. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Virtue A, Mai J, Yin Y, et al: Structural
evidence of anti-atherogenic microRNAs. Front Biosci (Landmark Ed).
16:3133–3145. 2011. View
Article : Google Scholar
|
|
31
|
Shan Z, Yao C, Li ZL, et al:
Differentially expressed microRNAs at different stages of
atherosclerosis in ApoE-deficient mice. Chin Med J (Engl).
126:515–520. 2013.
|
|
32
|
Ding S, Liang Y, Zhao M, et al: Decreased
microRNA-142-3p/5p expression causes CD4+ T cell activation and B
cell hyperstimulation in systemic lupus erythematosus. Arthritis
Rheum. 64:2953–2963. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Park S, Kang S, Min KH, et al:
Age-associated changes in microRNA expression in bone marrow
derived dendritic cells. Immunol Invest. 42:179–190. 2013.
View Article : Google Scholar
|
|
34
|
Gray EE and Cyster JG: Lymph node
macrophages. J Innate Immun. 4:424–436. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Hutchinson JA, Riquelme P, Geissler EK and
Fändrich F: Isolation of murine macrophages. Methods Mol Biol.
6:181–192. 2011.
|
|
36
|
Chadban SJ, Wu H and Hughes J: Macrophages
and kidney transplantation. Semin Nephrol. 30:278–289. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Seimon TA, Nadolski MJ, Liao X, et al:
Atherogenic lipids and lipoproteins trigger CD36-TLR2-dependent
apoptosis in macrophages undergoing endoplasmic reticulum stress.
Cell Metab. 12:467–482. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Liao X, Sluimer JC, Wang Y, et al:
Macrophage autophagy plays a protective role in advanced
atherosclerosis. Cell Metab. 15:545–553. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Tsukano H, Gotoh T, Endo M, et al: The
endoplasmic reticulum stress-C/EBP homologous protein
pathway-mediated apoptosis in macrophages contributes to the
instability of atherosclerotic plaques. Arterioscler Thromb Vasc
Biol. 30:1925–1932. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Inagaki Y, Yamagishi S, Amano S, et al:
Interferon-gamma-induced apoptosis and activation of THP-1
macrophages. Life Sci. 71:2499–2508. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Jin ZG, Lungu AO, Xie L, Wang M, Wong C
and Berk BC: Cyclophilin A is a proinflammatory cytokine that
activates endothelial cells. Arterioscler Thromb Vasc Biol.
24:1186–1191. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
de Winther MP, Kanters E, Kraal G and
Hofker MH: Nuclear factor kappaB signaling in atherogenesis.
Arterioscler Thromb Vasc Biol. 25:904–914. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Beswick EJ, Pinchuk IV, Earley RB, Schmitt
DA and Reyes VE: Role of gastric epithelial cell-derived
transforming growth factor beta in reduced CD4+ T cell
proliferation and development of regulatory T cells during
Helicobacter pylori infection. Infect Immun. 79:2737–2745. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Bartram U, Molin DG, Wisse LJ, Mohamad A,
Sanford LP, Doetschman T, et al: Double-outlet right ventricle and
overriding tricuspid valve reflect disturbances of looping,
myocardialization, endocardial cushion differentiation, and
apoptosis in TGF-beta(2)-knockout mice. Circulation. 103:2745–2752.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Singla DK, Singla RD, Lamm S and Glass C:
TGF-β2 treat ment enhances cytoprotective factors released from
embryonic stem cells and inhibits apoptosis in infarcted
myocardium. Am J Physiol Heart Circ Physiol. 300:H1442–H1450. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Mallat Z, Gojova A, Marchiol-Fournigault
C, et al: Inhibition of transforming growth factor-beta signaling
accelerates atherosclerosis and induces an unstable plaque
phenotype in mice. Circ Res. 89:930–934. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Lyons RE, Anthony JP, Ferguson DJ, et al:
Immunological studies of chronic ocular toxoplasmosis:
up-regulation of major histocompatibility complex class I and
transforming growth factor beta and a protective role for
interleukin-6. Infect Immun. 69:2589–2595. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Sun CK, Chua MS, He J and So SK:
Suppression of glypican 3 inhibits growth of hepatocellular
carcinoma cells through up-regulation of TGF-beta2. Neoplasia.
13:735–747. 2011.PubMed/NCBI
|