Introduction
Colorectal cancer is one of the most prevalent types
of cancer, with a high incidence of disease-related mortality and
morbidity (1). The development of
colorectal cancer is a multi-step process that is regulated by
complex molecular networks. These networks are altered via
sequential alterations in oncogenes, tumor-suppressor genes and
non-coding RNAs (ncRNAs). microRNAs (miRs) are a type of ncRNA
molecules, which negatively regulate protein expression at the
post-transcriptional level by interacting with the 3′-untranslated
region (3′-UTR) of target mRNAs and by inhibiting protein
translation. Therefore, understanding the role of miRs is critical
for defining cancer pathogenesis and developing new methods for
diagnosis and treatment.
The family of miR-34 comprises some of the most
studied miRs, which have been described as tumor suppressor genes
in multiple cancer types including melanoma (2), pancreatic (3), prostate (4), colorectal (5) and non-small cell lung cancer
(6) and neuroblastoma (7). miR-34a maps to the 1p36 genomic
region in humans, and is expressed at higher levels compared to
other family members. Several studies have indicated that
upregulation of miR-34a can induce apoptosis, senescence,
differentiation, cell-cycle arrest, and growth suppression
(8–10). The abnormal expression of miR-34a
results in cell-cycle arrest, growth inhibition and attenuated
chemoresistance to antitumor drugs. It was previously suggested
that miR-34a has a potential role in the treatment of p53-defective
prostate cancer (4). miR-34a is
also a promising therapeutic target for patients with
hormone-refractory prostate cancer or patients showing drug
resistance, where conventional chemical drug treatment exerts
limited effects, or patients with distant tumor metastasis and
recurrence (11). Sirtuin 1
(SIRT1) is a nicotinamide adenine dinucleotide (NAD)-dependent
histone deacetylase, which has been associated with inflammation,
circadian rhythm, hypoxic responses, cell survival, longevity and
metabolic processes (12–15). SIRT1 is also involved in the
mitochondrial antioxidant capacity, attenuating oxidative stress in
coronary arterial endothelial cells (16). The tumor protein p53 is a sensor of
chronic or acute alterations in cellular physiology, and more
importantly, engages with DNA to maintain chromosomal integrity
(17). The p53 levels are
associated with those of miR-34a in keratinocytes (18), human mammary epithelial cells
(19) and lymphoblast cell lines
(20). miR-34a enhances p53
activity through a decrease in deacetylation, which in turn results
in a decrease in SIRT1 expression. This decrease is achieved at the
post-transcriptional level through binding to the 3′-UTR (21,22).
In addition, inhibition of SIRT1 activates p53-dependent apoptosis
via deacetylation and stabilization of p53. miR-34a-mediated
inhibition of SIRT1 led to apoptosis in wild-type human colon
cancer cells, while no apoptosis was observed in p53-deficient
cancer cells (23). The positive
feedback loop involving p53, SIRT1 and miR-34a may thus provide new
therapeutic tools for the treatment of cancer.
However, the effect of the combination of miR-34a
and chemotherapeutic drugs on colorectal cancer has rarely been
syetematically explored. In addition, there are no reports
investigating the synergistic effect of miR-34a with 5-fluorouracil
(5-FU) on SW480 cells. In this study, we explored the effects of
miR-34a in cell invasion, migration and apoptosis in SW480 cells.
We further investigated the antitumor effect of both miR-34a and
5-FU in SW480 cells. Our experimental data provides evidence that
miR-34a may be a suitable molecular target for colorectal cancer
therapy. Finally, we examined the physiological pathway involving
miR34a, p53 and SIRT1, which may be involved in the observed
effects.
Materials and methods
Cell culture, transfection and
treatments
SW480 cells were obtained from Nanfang Hospital,
Southern Medical University (Guangzhou, China). The cells were
cultured in Dulbecco’s modified Eagle’s medium (DMEM, HyClone
Logan, UT, USA) with 10% fetal bovine serum (FBS; Sijiqing,
Hangzhou, China) and 100 U/ml of penicillin and streptomycin,
following standard procedures. Transfections were performed using
Invitrogen™ Lipofectamine® 2000 (Thermo Fisher
Scientific, Waltham, MA, USA). Cells were treated as follows:
negative control mimic (control group), 100 nM miR-34a mimic
(miR-34a group), 200 μg/ml 5-FU (5-FU group), or 200 μg/ml 5-FU
plus 100 nM miR-34a mimic (5-FU + miR-34a group) for 48 h. The
negative control and the miR-34a mimics were obtained from GeneChem
(Shanghai, China), and 5-FU was purchased from Jinyao Amino Acid
Co., Ltd. (Tianjin, China).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted using TRIzol reagent (Thermo
Fisher Scientific, Bremen, Germany) according to the manufacturer’s
instructions. The first strand cDNA was synthesized by stem-loop
primer reverse transcription reaction (Thermo Fisher Scientific).
The following primer sequences were used: Hsa-miR-34a RT primer,
5′-CTCAACTGGTGTCGTGGAG TCGGCAATTCAGTTGAGACAACCAG-3′; sense: 5′-ACA
CTCCAGCTGGGTGGCAGTGTCTTAG-3′; and antisense:
5′-CTCAACTGGTGTCGTGGAGTCG-3′ for Hsa-miR-34a; and sense:
5′-CTCGCTTCGGCAGCACA-3′ and antisense: 5′-AACGCTTCACGAATTTGCGT-3′
for U6. The real-time quantitative PCR kit (Fermentas, Helsingborg,
Sweden) was used to facilitate the reactions. Reactions were
conducted on an Applied BioSystems 7900HT thermocycler (Applied
Biosystems, Inc.) and performed under the following thermal cycling
conditions: 95°C for 10min, followed by 40 cycles of 95°C for 15
sec, 60°C for 30 sec and 72°C for 15 sec; followed by a 60°C for 1
min, 95°C for 15 sec. Raw data of all samples were normalized to
that of the control and fold changes were calculated using a
relative quantification equation (RQ=2−ΔΔCt).
Western blot analysis
Western blot analysis was performed as previously
described (24). Briefly, SW480
cells were homogenized in phosphate-buffered saline (PBS)
containing a protease inhibitor cocktail (Beyotime Institute of
Biotechnology, Shanghai, China). The samples were incubated
overnight at 4°C with rabbit anti-p53 antibody, -acetyl p53,
-SIRT1, or -acetyl p21 antibody (all from Cell Signaling Technology
Inc., Danvers, MA, USA). The antibody signal was detected using a
Chemiluminescent Detection kit according to the manufacturer’s
protocol (Beyotime Institute of Biotechnology, Jiangsu, China). The
relative band intensities in the blots were determined using the
Adobe Photoshop software (Adobe Systems Inc., San Jose, CA,
USA).
Apoptosis analysis
Following treatment for 48 h as described above,
SW480 cells were harvested, washed in ice-cold PBS, resuspended in
500 μl of binding buffer (C1062-2,Beyotime Institute of
Biotechnology, Jiangsu, China) and incubated for 15 min in the dark
with 5 μl of propidium iodide (PI; Beyotime Institute of
Biotechnology, Jiangsu, China) and 5 μl of Annexin V-fluorescein
isothiocyanate (FITC; Beyotime Institute of Biotechnology, Jiangsu,
China). The samples were washed and resuspended in 500 μl PBS, and
immediately analyzed by fluorescence-activated cell sorting (FACS)
on a EPICS XL-MCL flow cytometer (Beckman Coulter, Brea, CA,
USA).
Cell cycle analysis
Following a 48 h treatment, SW480 cells were
harvested, washed with PBS, and fixed in ice-cold 70% ethanol.
Fixed cells were treated with DNase-free RNaseA (TransGen Biotech,
Beijing, China) in PBS at 37°C for 30 min, followed by staining
with PI at room temperature for 10 min. The proportion of cells at
the different stages of the cell cycle was estimated by flow
cytometry.
Transwell cell migration assay
SW480 cells (48 h post-treatment) were trypsinized
with 0.25% trypsin (Beyotime Institute of Biotechnology, Jiangsu,
China) and suspended in serum-free DMEM at 5×105
cells/ml. A total of 200 μl of the cell suspension were placed in
the top chamber of a two-chamber Transwell assay system (Corning
Inc., Corning, NY, USA) and 800 μl of medium containing 10% FBS
were added in the lower chamber. Cells were cultured at 37°C for 12
h. The cells on the surface of the upper chamber were swapped and
the cells under the surface of the lower chamber were stained with
crystal violet (0.1%). Cell migration was evaluated by counting the
cells that had migrated into the filters.
Transwell cell invasion assay
Similar to the migration assay, 50 μl BD Matrigel™
(BD Biosciences, Franklin Lakes, NJ, USA) was added into each
Transwell upper chamber and placed in a 37°C incubator for 2 h to
solidify. The tumor cell invasive capacity was then assessed
similarly to the cell migration assay.
Statistical analysis
The results are expressed as mean ± standard
deviation. Statistical significance was determined with Student’s
t-tests (two-tailed, unpaired). P<0.05 was considered to
indicate a statistically significant difference.
Results
miR-34a enhances the 5-FU effect on the
SIRT1/p53 pathway in SW480 cells
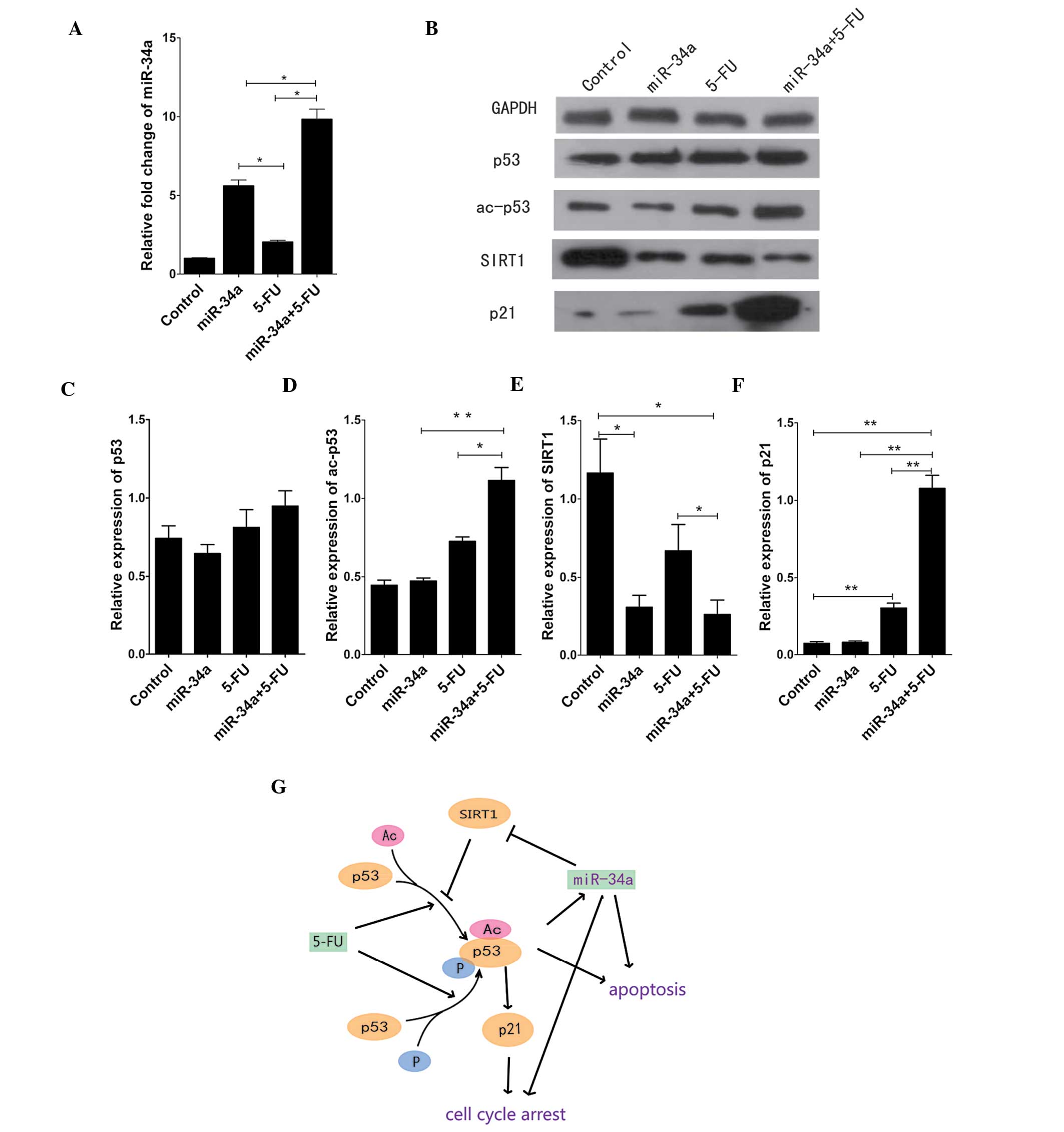
To first understand the effects of miR-34a, the
expression level of this miR was measured by RT-qPCR in SW480 cells
before and after transfection with the miR-34a mimic. The level of
miR-34a in SW480 cells after transfection was markedly higher
compared to the control group. In addition, the combination of the
miR-34a mimic and 5-FU showed a synergistic effect on miR-34a
expression (Fig. 1A). We also
examined the protein expression of p53 and acetylated (ac)-p53 by
western blot analysis. There was no significant change in the p53
level after treatment with the miR-34a mimic or 5-FU. The level of
p53 was slightly but not significantly increased following combined
treatment with the miR-34a mimic and 5-FU. By contrast, the
combined treatment increased the level of ac-p53 compared to the
control group (p<0.05), while no change was observed when
miR-34a was used alone (Fig. 1B and
D). To further understand the miR-34a pathway, we examined the
protein expression of SIRT1 and p21, and found that SIRT1
expression is significantly decreased following treatment with
miR-34a compared to the control group (p<0.05). 5-FU had a
similar effect on SIRT1 expression in SW480 cells, with the changes
being statistically significant (p<0.05) (Fig. 1B and E). In addition, the level of
p21 was significantly and markedly changed by the combined miR-34a
+ 5-FU treatment compared to the other groups (p<0.01) (Fig. 1B and F). The results from western
blot analysis were used in combination with published data to
create a model illustrating the relationships among miR-34a, 5-FU
and SIRT1/p53 (Fig. 1G).
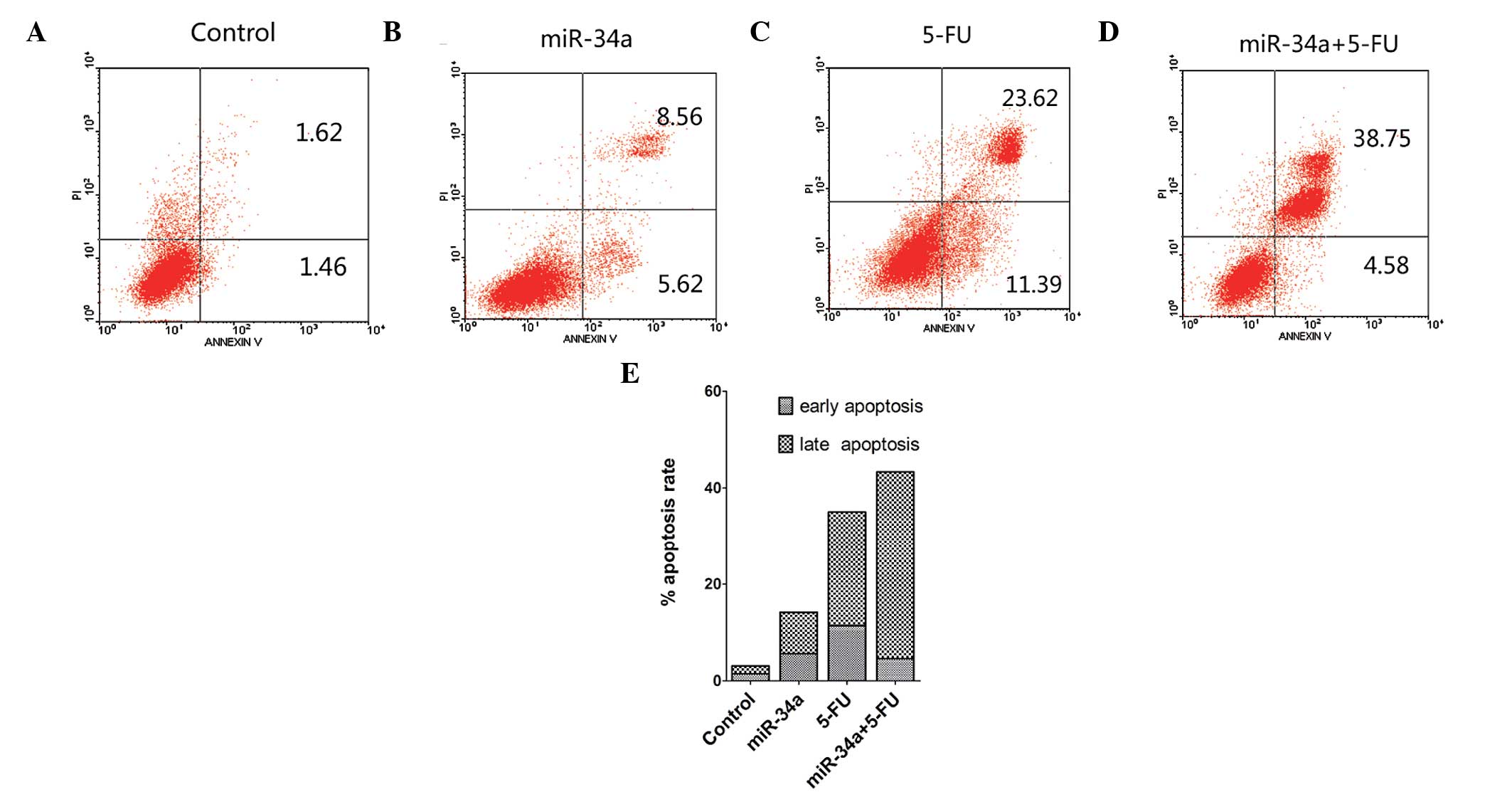
miR-34a induces apoptosis in SW480 cells
and acts synergistically with 5-FU
To determine the effects of miR-34a and 5-FU
treatment on cell death, we double-stained SW480 cells with Annexin
V-FITC and PI and analyzed apoptosis by FACS at 48 h
post-treatment. The shifts in cell population with the different
treatments clearly indicated that the apoptotic rate of the miR-34a
+ 5-FU-treated group is higher than that of the miR-34a or the 5-FU
group, while the control group showed the lowest rate of apoptosis
(Fig. 2).
miR-34a blocks the cell cycle in synergy
with 5-FU
We further examined the effect of miR-34a on the
SW480 cell cycle by flow cutometry. This assay showed that both
miR-34a and 5-FU block cell cycle progression and have a
synergistic effect when combined. Individually, both miR-34a and
5-FU increased the percentage of cells detected at the G1 phase,
and the combined treatment further increased this percentage, while
the control group had the lowest proportion of G1-phase cells
(Fig. 3A–D). The cell-cycle
machinery involves the cyclin-dependent kinases (CDK)/cyclin
complex, and p21 is known to suppress CDK1 activity via a
p53-independent pathway. This event blocks cell progression into
the G2/M phase. Moreover, p21 can block the progression of cells
into the S phase, through inhibition of the CDK2 activity. miR-34a
is involved in this cascade by blocking the progression of cells
into the S phase through inhibition of the CDK2/4/6 activity
(Fig. 3F).
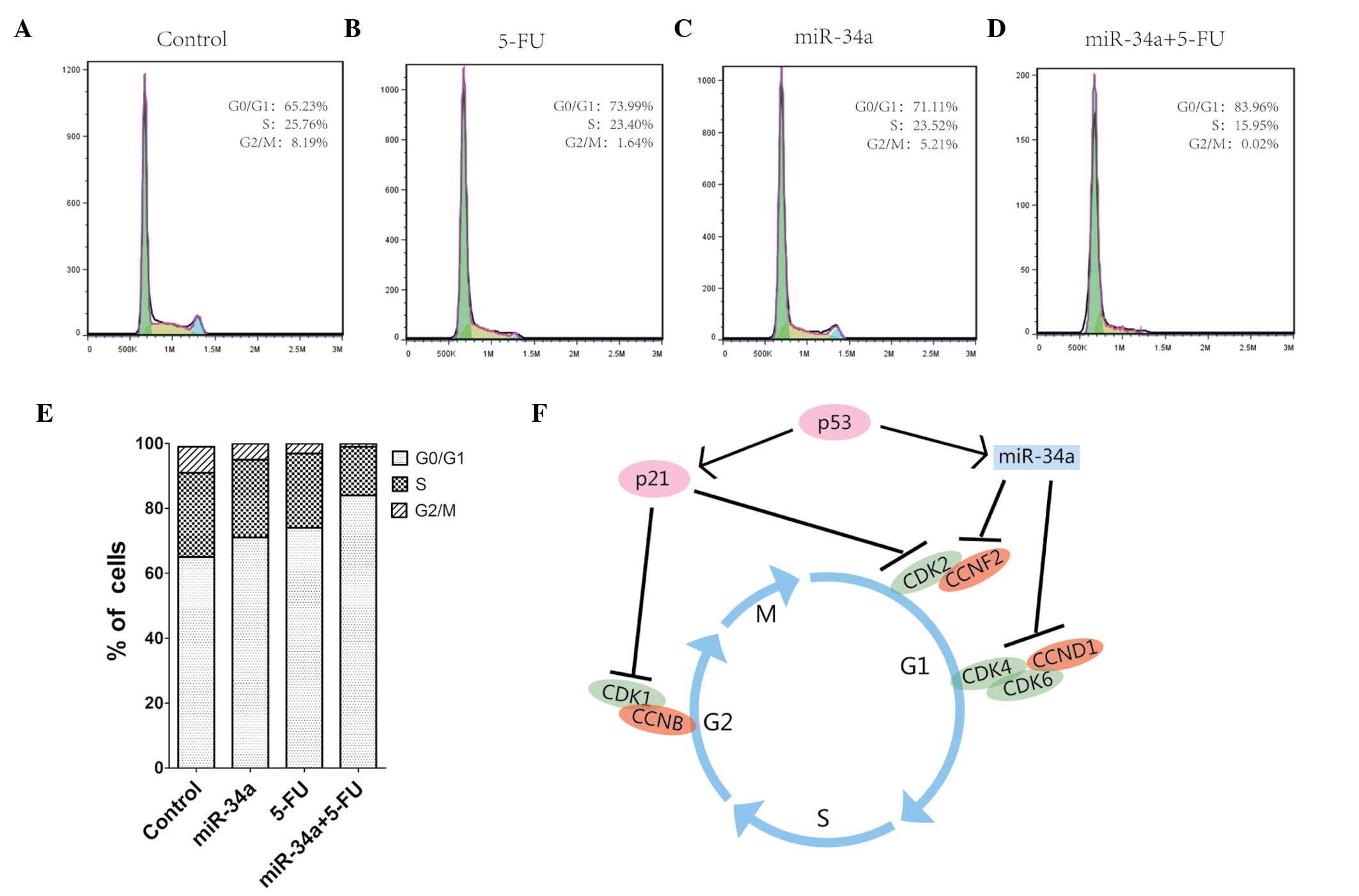 | Figure 3miR-34a inhibits the cycle phase
transition from G0/G1 to S. (A–D) SW480 cells were treated with the
negative control mimic (control), miR-34a mimic, 5-FU, or both, and
were analyzed by flow cytometry to determine the percentage of
cells in each of the different cell cycle phases, G1, S, and G2/M.
(E) Bar diagram illutrating the distribution of cells from each
group in the different cell cycle phases. (F) A model summarizing
the findings from this study combined with previously published
data, and describing the molecular interactions involved in cell
cycle regulation. miR, microRNA; 5-FU, 5-fluorouracil; CDK,
cyclin-dependent kinase; CCND1, cyclin D1. |
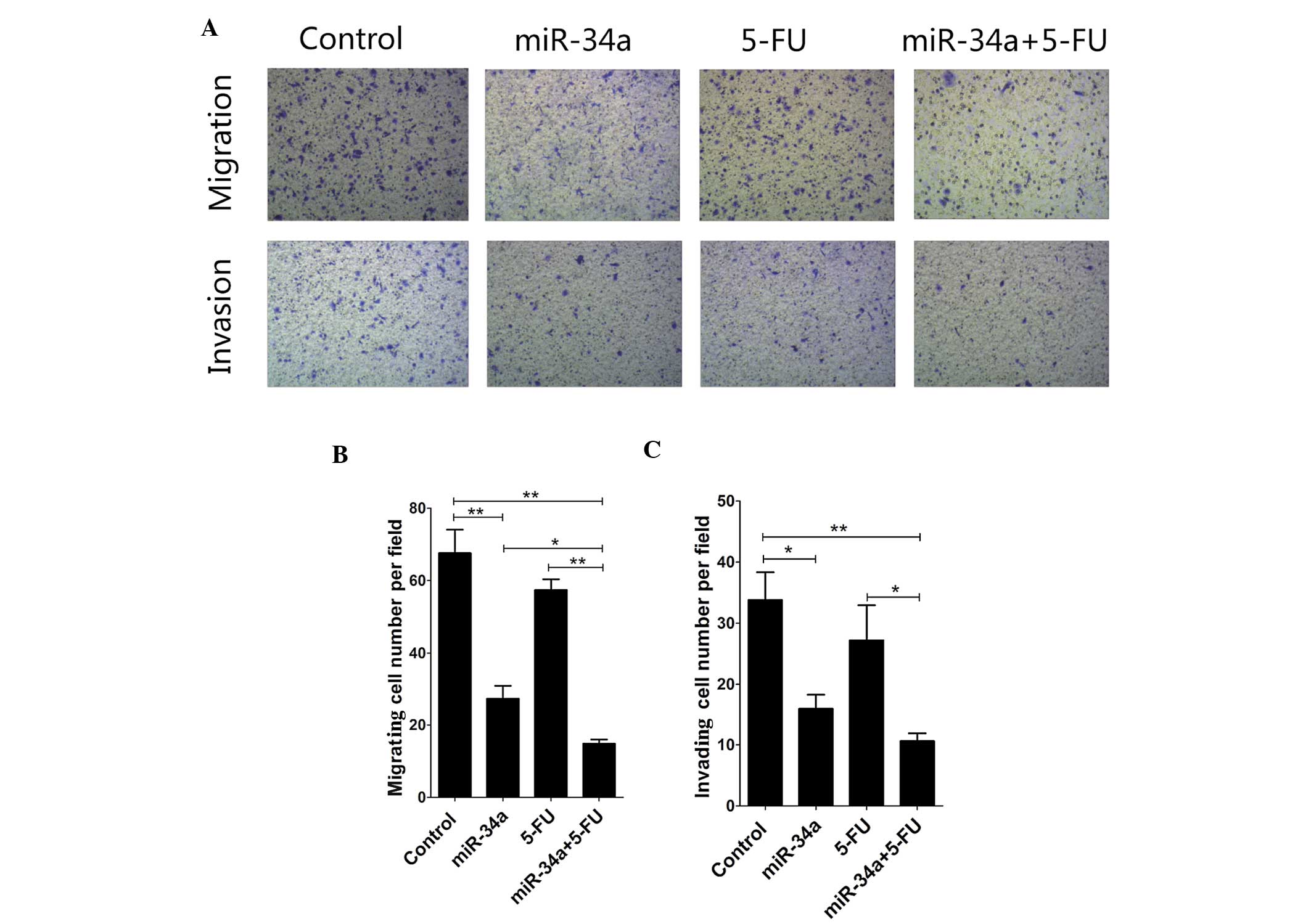
miR-34a inhibits migration and invasion
of SW480 cells in vitro
To investigate whether miR-34a also plays a role in
cell migration and invasion, we tested if SW480 cells have the
potential to digest Matrigel and migrate through the 8-mm membrane
pores of a Transwell chamber. The Transwell tumor cell migration
assay demonstrated that the miR-34a mimic-treated cells have a
significantly reduced migrating capacity compared to the control
group (p<0.01), and this effect was enhanced with the combined
treatment miR-34a mimic + 5-FU (p<0.05) (Fig. 4A and B). The Transwell tumor cell
invasion assay also showed that transfection with the miR-34a mimic
significantly inhibits the invasive capacity of SW480 cells
(p<0.05). This effect was enhanced with the miR-34a mimic + 5-FU
combination, compared to 5-FU treatment alone (p<0.05) (Fig. 4A and C).
Discussion
SW480 cells were used as a model to investigate the
biological function of miR-34a in the context of colorectal cancer.
The transient overexpression of miR-34a combined with 5-FU
treatment reduced cell migration and invasion and increased
apoptosis in these cells. These changes were most probably a result
of the increased expression of ac-p53 and p21 and the decreased
expression of SIRT1. These results demonstrate that miR-34a
regulates the expression of critical proteins involved in cell
apoptosis, proliferation and the response to chemotherapy.
Moreover, miR-34a allowed sensitization of colon cancer cells to
5-FU, likely acting through the p53/SRIT1 pathway.
Typically, miRs are transcribed and processed in the
nucleus to form pre-miRs. These pre-miRs are then exported to the
cytoplasm and processed into miR duplexes. One strand from the
duplex is incorporated into the miR-induced silencing complex
(25). The study of miRs is a
rapidly expanding research field, which includes investigation of
their roles in tumor- or non-tumor-related diseases. miRs play an
important role in cell function and fate in both the disease and
the homeostatic states. These molecules are continuously reported
as oncogenes or tumor suppressor genes. Not only are they detected
in virtually every type of tumor, but they also display specific
profiles in pathologies, which allow assessingmalignancy and
evaluating the potential for metastasis (26). Increasing evidence has ascertained
that a large number of miRs exhibit dysregulated expression in
primary cancer specimens compared to tissues from healthy patient
populations, including miR-21, miR-125b, miR-143, miR-145, miR-10b,
miR-26a, miR-155 and miR-301 (27,28).
Although the tumor inhibition effect of miR-34a has
been previously documented, its therapeutic potential on colorectal
cancer remained unclear to date. Most research studies so far have
focused on the role of miR-34a as a p53 transcriptional target and
on its involvement in p53-mediated tumor suppression processes
(29). Reduced or no miR-34a
expression has been detected in a variety of tumors and cancer cell
lines. To understand the role of miR-34a in SW480 cells, we tested
the effects of miR-34a transfection on cell migration and invasion,
and demonstrated that miR-34a inhibits SW480 cell migration and
invasion; notably, this inhibition effect is enhanced when miR-34a
transfection is combined with 5-FU treatment (Fig. 4). FACS analysis further showed
indicated that miR-34a induces SW480 cell apoptosis,an effect again
enhanced by 5-FU treatment (Fig.
2). These data overall suggest that miR-34a exerts an important
antitumor effect on SW480 cells, which is similar to results
reported in other studies on chronic lymphocytic leukemia (30), lung cancer (31), mesothelioma (10), neuroblastoma (32), prostate (33) and pancreatic cancer (34,35).
miR-34a induces cell cycle arrest by downregulating
cell-cycle-related proteins such as cyclin D1 (CCND1), cyclin E2
(CCNE2), CDK4 and CDK6. Our data suggests that p53, p21 and miR-34a
form a strong interaction network in the cell cycle (Fig. 3F). miR-34a has documented roles in
the increase of acetylated p53 and in modification of p21
expression through the inhibition of SIRT1 expression. Our study
confirmed that miR-34a significantly decreases the SIRT1 protein
level, and the levels of ac-p53 and p21 were found particularly
increased with the combined miR-34a + 5-FU treatment (Fig. 4). 5-FU is a pyrimidine
antimetabolite cytotoxin, which induces DNA and RNA damage,
resulting in cell death. 5-FU functions in a p53-dependent manner,
likely causing changes in DNA metabolism and initiating events that
culminate in the alteration of p53 expression (35). When the damaged cells cannot be
repaired, p53 triggers cell elimination by inducing the expression
of pro-apoptotic genes such as Fas and Bax (36). Interestingly, miR-34a can
negatively regulate 5-FU resistance in human colorectal cancer
DLD-1 cells by targeting the SIRT1 and E2F3 genes
(37).
SIRT1 inactivates p53 by deacetylating a specific
lysine residue to target it degradation. An increase in the miR-34a
and a decrease in the SIRT1 levels were observed in leukemic cells
that had been simultaneously exposed to nicotinamide and etoposide
(38). miR-34a restoration alone
confers drug resistance via the SIRT1-NFκB pathway in tumors with
p53 deficiency, which renders the combination of an NF-κB inhibitor
and miR-34a a promising therapeutic strategy (39). A previous study indicated that
SIRT1 regulates the expression of several antioxidant genes in
bovine aortic endothelial cells, including MnSOD,
Prx3, Prx5, Trx2, TR2, and UCP-2
(40), which may be involved in
the p53-independent pathway.
In summary, our data has demonstrated, for the first
time to the best of our knowledge, that miR-34a reduces the
migratory and invasive ability of SW480 cells, and induces
apoptosis and cell cycle arrest, in a synergetic manner with 5-FU.
As previously reported, miR-34a plays an important role as an
apoptotic mediator, by alleviating drug resistance of colorectal
cancer cells through the SIRT1/p53 pathway. Our data clearly
illustrates the therapeutic potential of miR-34a, especially in
combination with 5-FU, in the treatment of colorectal cancer.
Acknowledgements
This study was supported by grants from the Natural
Science Foundation of Guangdong Province (10151802001000002), the
Science and Technology Planning Project of Shenzhen (201201013),
and the Technical Research and Development Project of Shenzhen
(JCYJ20130402092657774).
References
|
1
|
Stillwell AP, Buettner PG, Siu SK, Stitz
RW, Stevenson AR and Ho YH: Predictors of postoperative mortality,
morbidity, and long-term survival after palliative resection in
patients with colorectal cancer. Dis Colon Rectum. 54:535–544.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Cozzolino AM, Pedace L, Castori M, et al:
Analysis of the miR-34a locus in 62 patients with familial
cutaneous melanoma negative for CDKN2A/CDK4 screening. Fam Cancer.
11:201–208. 2012. View Article : Google Scholar
|
|
3
|
Nalls D, Tang SN, Rodova M, Srivastava RK
and Shankar S: Targeting epigenetic regulation of miR-34a for
treatment of pancreatic cancer by inhibition of pancreatic cancer
stem cells. PLoS One. 6:e240992011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Fujita Y, Kojima K, Hamada N, et al:
Effects of miR-34a on cell growth and chemoresistance in prostate
cancer PC3 cells. Biochem Biophys Res Commun. 377:114–119. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Winton DJ: miR-34a sets the ‘sweet spot’
for notch in colorectal cancer stem cells. Cell Stem Cell.
12:499–501. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Gallardo E, Navarro A, Vinolas N, et al:
miR-34a as a prognostic marker of relapse in surgically resected
non-small-cell lung cancer. Carcinogenesis. 30:1903–1909. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Cole KA, Attiyeh EF, Mosse YP, et al: A
functional screen identifies miR-34a as a candidate neuroblastoma
tumor suppressor gene. Mol Cancer Res. 6:735–742. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Dorn GN II: miR-34a and the cardiomyopathy
of senescence: SALT PNUTS, SALT PNUTS! Cell Metab. 17:629–630.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ito T, Yagi S and Yamakuchi M:
MicroRNA-34a regulation of endothelial senescence. Biochem Biophys
Res Commun. 398:735–740. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ghawanmeh T, Thunberg U, Castro J, Murray
F and Laytragoon-Lewin N: miR-34a expression, cell cycle arrest and
cell death of malignant mesothelioma cells upon treatment with
radiation, docetaxel or combination treatment. Oncology.
81:330–335. 2011. View Article : Google Scholar
|
|
11
|
Kojima K, Fujita Y, Nozawa Y, Deguchi T
and Ito M: MiR-34a attenuates paclitaxel-resistance of
hormone-refractory prostate cancer PC3 cells through direct and
indirect mechanisms. Prostate. 70:1501–1512. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Guarente L: Franklin H. Epstein Lecture:
sirtuins, aging, and medicine. N Engl J Med. 364:2235–2244. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Li X and Kazgan N: Mammalian sirtuins and
energy metabolism. Int J Biol Sci. 7:575–587. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li YG, Zhu W, Tao JP, et al: Resveratrol
protects cardiomyocytes from oxidative stress through SIRT1 and
mitochondrial biogenesis signaling pathways. Biochem Biophys Res
Commun. 438:270–276. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Cui Y, Wang H, Chen H, et al: Genetic
analysis of the SIRT1 gene promoter in myocardial infarction.
Biochem Biophys Res Commun. 426:232–236. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Csiszar A, Labinskyy N, Jimenez R, et al:
Anti-oxidative and anti-inflammatory vasoprotective effects of
caloric restriction in aging: role of circulating factors and
SIRT1. Mech Ageing Dev. 130:518–527. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Olivier M, Petitjean A, Marcel V, et al:
Recent advances in p53 research: an interdisciplinary perspective.
Cancer Gene Ther. 16:1–12. 2009. View Article : Google Scholar
|
|
18
|
Lefort K, Brooks Y, Ostano P, et al: A
miR-34a-SIRT6 axis in the squamous cell differentiation network.
EMBO J. 32:2248–2263. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang B, Li D and Kovalchuk O: p53 Ser15
phosphorylation and histone modifications contribute to IR-induced
miR-34a transcription in mammary epithelial cells. Cell Cycle.
12:2073–2083. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chen X, Yan J and Chen T: Expression level
of miR-34a rather than P53 gene status correlates with mutability
in related human lymphoblast cell lines. Mol Carcinog. 51:674–677.
2012. View
Article : Google Scholar
|
|
21
|
Yamakuchi M and Lowenstein CJ: MiR-34,
SIRT1 and p53: the feedback loop. Cell Cycle. 8:712–715. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Castro RE, Ferreira DM, Afonso MB, et al:
miR-34a/SIRT1/p53 is suppressed by ursodeoxycholic acid in the rat
liver and activated by disease severity in human non-alcoholic
fatty liver disease. J Hepatol. 58:119–125. 2013. View Article : Google Scholar
|
|
23
|
Yamakuchi M, Ferlito M and Lowenstein CJ:
miR-34a repression of SIRT1 regulates apoptosis. Proc Natl Acad Sci
USA. 105:13421–13426. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kouchi Z, Fujiwara Y, Yamaguchi H,
Nakamura Y and Fukami K: Phosphatidylinositol 5-phosphate 4-kinase
type II beta is required for vitamin D receptor-dependent
E-cadherin expression in SW480 cells. Biochem Biophys Res Commun.
408:523–529. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Gregory RI, Chendrimada TP and Shiekhattar
R: MicroRNA biogenesis: isolation and characterization of the
microprocessor complex. Methods Mol Biol. 342:33–47.
2006.PubMed/NCBI
|
|
26
|
Krutovskikh VA and Herceg Z: Oncogenic
microRNAs (OncomiRs) as a new class of cancer biomarkers.
Bioessays. 32:894–904. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wei Y, Nazari-Jahantigh M, Neth P, Weber C
and Schober A: MicroRNA-126, -145, and -155: a therapeutic triad in
atherosclerosis? Arterioscler Thromb Vasc Biol. 33:449–454. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Krichevsky AM and Gabriely G: miR-21: a
small multi-faceted RNA. J Cell Mol Med. 13:39–53. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
He L, He X, Lim LP, et al: A microRNA
component of the p53 tumour suppressor network. Nature.
447:1130–1134. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Dufour A, Palermo G, Zellmeier E, et al:
Inactivation of TP53 correlates with disease progression and low
miR-34a expression in previously treated chronic lymphocytic
leukemia patients. Blood. 121:3650–3657. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Duan W, Xu Y, Dong Y, Cao L, Tong J and
Zhou X: Ectopic expression of miR-34a enhances radiosensitivity of
non-small cell lung cancer cells, partly by suppressing the LyGDI
signaling pathway. J Radiat Res. 54:611–619. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Feinberg-Gorenshtein G, Avigad S, Jeison
M, et al: Reduced levels of miR-34a in neuroblastoma are not caused
by mutations in the TP53 binding site. Genes Chromosomes Cancer.
48:539–543. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Liu C, Kelnar K, Liu B, et al: The
microRNA miR-34a inhibits prostate cancer stem cells and metastasis
by directly repressing CD44. Nat Med. 17:211–215. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Pritchard DM, Watson AJ, Potten CS,
Jackman AL and Hickman JA: Inhibition by uridine but not thymidine
of p53-dependent intestinal apoptosis initiated by 5-fluorouracil:
evidence for the involvement of RNA perturbation. Proc Natl Acad
Sci USA. 94:1795–1799. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Roos WP and Kaina B: DNA damage-induced
cell death: from specific DNA lesions to the DNA damage response
and apoptosis. Cancer Lett. 332:237–248. 2013. View Article : Google Scholar
|
|
36
|
Wiman KG: Strategies for therapeutic
targeting of the p53 pathway in cancer. Cell Death Differ.
13:921–926. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Akao Y, Noguchi S, Iio A, Kojima K, Takagi
T and Naoe T: Dysregulation of microRNA-34a expression causes
drug-resistance to 5-FU in human colon cancer DLD-1 cells. Cancer
Lett. 300:197–204. 2011. View Article : Google Scholar
|
|
38
|
Audrito V, Vaisitti T, Rossi D, et al:
Nicotinamide blocks proliferation and induces apoptosis of chronic
lymphocytic leukemia cells through activation of the
p53/miR-34a/SIRT1 tumor suppressor network. Cancer Res.
71:4473–4483. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Tan J, Fan L, Mao JJ, et al: Restoration
of miR-34a in p53 deficient cells unexpectedly promotes the cell
survival by increasing NFκB activity. J Cell Biochem.
113:2903–2908. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Olmos Y, Sanchez-Gomez FJ, Wild B, et al:
SirT1 regulation of antioxidant genes is dependent on the formation
of a FoxO3a/PGC-1α complex. Antioxid Redox Signal. 19:1507–1521.
2013. View Article : Google Scholar : PubMed/NCBI
|