Introduction
Vitiligo is a prevalent disorder, affecting 0.5–1%
of the population worldwide, which results in depigmented areas of
the skin (1). The absence of
melanocytes in the skin lesions was previously reported to be the
key event in the pathogenesis of vitiligo (2); however, the aetiology of vitiligo
remains to be elucidated. Previous studies have suggested that
oxidative stress may result in the loss of melanocytes (3,4);
increased intracellular reactive oxygen species (ROS) production
was observed in the epidermis of vitiligo patients (5), which therefore indicates the
presences of systemic oxidative stress in vitiligo (6,7).
Accumulated oxidative stress leads to DNA damage, lipid and protein
peroxidation and cell death (8,9).
Hydrogen peroxide (H2O2)-mediated oxidation
was reported to result in inhibition of tyrosinase (10) and the significant decrease in
acetylcholine esterase (AChE) activity (11). A previous study showed that
antioxidants may be resistant to cell death mediated by oxidative
stress. Green tea extract and quercetin were demonstrated to have
potent cytoprotective effects on H2O2-induced
cell death (12); in addition,
another study showed that quercetin inhibited
H2O2-induced melanocyte apoptosis (13). Furthermore, a double-blind placebo
controlled trial revealed that oral supplementation with an
antioxidant pool (AP)containing α-lipoic acid prior to and during
narrowband ultraviolet B (NB-UVB) exposure significantly improves
the clinical effectiveness of NB-UVB, reducing vitiligo-associated
oxidative stress (14).
Extensive dilation of the rough endoplasmic
reticulum (RER) was observed in numerous vitiligo patients;
however, the cause and the proteins involved remain to be
elucidated (15). A previous study
demonstrated that swollen ER were present in melanocytes
transfected with FBXO11 siRNA; tyrosinase, the rate-limiting
enzyme for melanin synthesis, was also reported to be regulated by
the FBXO11 gene (16).
Furthermore, H2O2 was found to induce
partially damaged plasma membranes, swollen RER and swollen or
deformed mitochondria with ruptured cristae (17). Therefore, it was suggested that
H2O2 may induce dilated ER and melanocyte
dysfunction.
The present study aimed to evaluate the effects of
H2O2 on the morphology of melanocyte ER and
the export of tyrosinase from the ER, as well as to determine the
mechanisms underlying the protective role of quercetin against the
effects of H2O2.
Materials and methods
Reagents
Culture medium and supplements were obtained from
Gibco-BRL (Carlsbad, CA, USA), with the exception of recombinant
human basic fibroblast growth factor (bFGF), isobutylmethylxanthine
(IBMX) and cholera toxin (CT), which were purchased from PeproTech,
Inc. (Rocky Hill, NJ, USA). H2O2 and
quercetin were purchased from Sigma-Aldrich (St. Louis, MO, USA).
Mouse monoclonal anti-tyrosinase and rabbit polyclonal
anti-calreticulin antibodies were purchased from Abcam (Cambridge,
MA, USA), anti-β-actin mouse monoclonal antibody was from Santa
Cruz Biotechnology, Inc. (Dallas, TX, USA), Alexa Fluor®
488 donkey anti-mouse Immunoglobulin G (IgG; heavy and light chain,
H+L) and Alexa Fluor® 594 donkey anti-rabbit IgG (H+L)
from Life Technologies (Grand Island, NY, USA) and IRDye 680RD goat
anti-mouse IgG highly cross adsorbed was from LI-COR Biosciences
(Lincoln, NE, USA).
Cell culture
Human epidermal melanocytes were obtained from
normal foreskins as previously described (18). Written informed consent was
obtained from the patients and the study was approved by the ethics
committee of the Third People’s Hospital of Hangzhou (Hangzhou,
China). In brief, the samples were incubated in a solution of 0.25%
trypsin and 0.2% ethylenediamine tetraacetic acid for 20 h;
trypsinization was terminated by the addition of Dulbecco’s
modified Eagle’s medium (Gibco-BRL) containing 10% fetal bovine
serum (FBS; Gibco-BRL). Melanocytes were then scraped from the
epidermis, washed with phosphate-buffered saline (PBS) and
centrifuged at 100 × g for 5 min. The pellet was resuspended in
Hu16 medium (F12 medium with 20 ng/ml bFGF, 20 μg/ml IBMX,
10 ng/ml CT, 50 μg/ml gentamicin and 10% FBS). Cells were
incubated in a humidified 95% air/5% CO2 atmosphere at
37°C. Geneticin was added to the medium (100 μg/ml) three
days later in order to eliminate contaminating cells. Primary
cultures were stored until they reached 80% confluence and then the
melanocytes were detached using 0.125% trypsin/0.01 M EDTA
solution, centrifuged at 100 × g for 5 min, resuspended and then
seeded into culture flasks for subculture. Melanocytes used in the
present study were of passage 2–3.
Exposure to
H2O2
In order to evaluate effect of
H2O2 on melanocytes, cultured cells were
seeded at 1×104 per well in 96-well plates at different
H2O2 concentrations (0–400 μM) for 24
h. Cell viability was then assessed using an MTT assay kit
(Promega, Sunnyvale, CA, USA).
Treatment of quercetin
In order to investigate the cytoprotective activity
of quercetin, cells (2×105/well) were seeded onto
six-well plates. Following serum starvation for 24 h, cells were
pretreated with quercetin (0–200 μM dissolved in NaOH) for
24 h, H2O2 was then added to each well and
incubated for 24 h. Cell viabilities were determined using an MTT
assay.
Cell viability assay
MTT assays were used to assess cell viability in
melanocytes following H2O2 and quercetin
treatment. Following treatment, 10 μl MTT (10 mg/ml) was
added to cells seeded in 96-well plates and incubated for 4 h, then
100 μl dimethyl sulfoxide was added for 15 min to dissolve.
The absorbance value was measured at 490 nm using a microplate
spectrophotometer (SoftMax Pro5; Molecular Devices, LLC, Sunnyvale,
CA, USA).
Intracellular ROS measurement
ROS levels were determined by measuring the
oxidative conversion of 2′,7′-dichloro-fluorescin diacetate
(DCFH-DA) into the fluorescent compound dichlorofluorescin (DCF)
using a ROS assay kit (Beyotime Insitute of Biotechnology, Jiangsu,
China). In brief, 1×104 cells/well were seeded into a
96-well plate and then the medium was replaced with serum-free
medium for 24 h starvation. Cells from each well were then
incubated with 10 μM DCFH-DA for 20 min at 37°C. Cells were
treated with quercetin or NaOH for 30 min and then
H2O2 was added to each well, except those
containing the untreated group, and the cells were incubated for 30
min. For the estimation of intracellular ROS, DCF fluorescence was
determined at 485 nm excitation and 520 nm emission using a flow
cytometer (BD FACSCalibur™; BD Biosciences, Franklin Lakes, NJ,
USA). Three independent experiments were performed.
Electron microscopy observation
Cells were seeded into a six-well plate and the
medium was replaced with serum-free medium for 24 h starvation.
Cells were pretreated with 0.01 μM NaOH or 25 μM
quercetin for 24 h at 37°C; 200 μM
H2O2 was then added to each well, except the
untreated group, and incubated for 24 h. ER configuration was
observed using electron microscopy as previously described
(16). In brief, cells were
collected and fixed with 2.5% glutaraldehyde in 0.1 M PBS at 4°C
overnight; cells were then post-fixed with 1% OsO4 in
0.1 M PBS at 4°C for 1 h. Cells were then embedded in 5% agarose
(Sigma-Aldrich) and cut into 2–3-mm2 blocks, dehydrated
in a graded series of ethanol and embedded in epoxy resin (West
System, Bay City, MI, USA). Ultrathin sections were stained with
uranyl acetate (Sigma-Aldrich) and lead citrate (Sigma-Aldrich) and
were examined using a transmission electron microscope (JEM-1230;
JEOL, Ltd., Tokyo, Japan).
Immunofluorescence assay
In order to examine the co-localization of
tyrosinase and calreticulin, cells were grown in six-well plates
containing coverslips, with various treatments. Cells were then
fixed with 4% formaldehyde in PBS for 30 min. Anti-tyrosinase and
anti-calreticulin antibodies were used at 10 μg/ml in buffer
(0.5% BSA in PBS) and incubated with the coverslips for 1 h. Cells
were then incubated with the secondary antibodies, Alexa
Fluor® 488 donkey anti-mouse IgG (H+L) for tyrosinase
and Alexa Fluor® 594 donkey anti-rabbit IgG (H+L) for
calreticulin, at a dilution of 1:500 in PBS for 30 min at room
temperature. Cover slips were washed three times in PBS for 5 mins
each, mounted using a mounting medium and observed with confocal
laser scanning microscope (TCS SP2; Leica Microsystems, Wetzlar,
Germany).
Western blot analysis of tyrosinase
Total proteins were isolated from cells and
separated on a 10% polyacrylamide gel. Proteins were measured using
western blot analysis with mouse antibodies against tyrosinase,
followed by incubation with IRDye 680RD goat anti-mouse IgG highly
cross adsorbed (1:10,000) for 1 h. β-actin was used as the internal
control. An infrared imaging system, Licor Odyssey (LI-COR
Biosciences), was used to visualize the protein bands and the
relative intensities of bands were quantified using Image Studio
for Licor Odyssey CLx and Classic (LI-COR Biosciences).
Statistical analysis
Values are presented as the mean ± standard
deviation. Comparisons among groups were analyzed by a one-way
analysis of variance using SPSS Version 13.0 (SPSS Inc., Chicago,
IL, USA). P<0.05 and P<0.01 were considered to indicate a
statistically significant difference.
Results
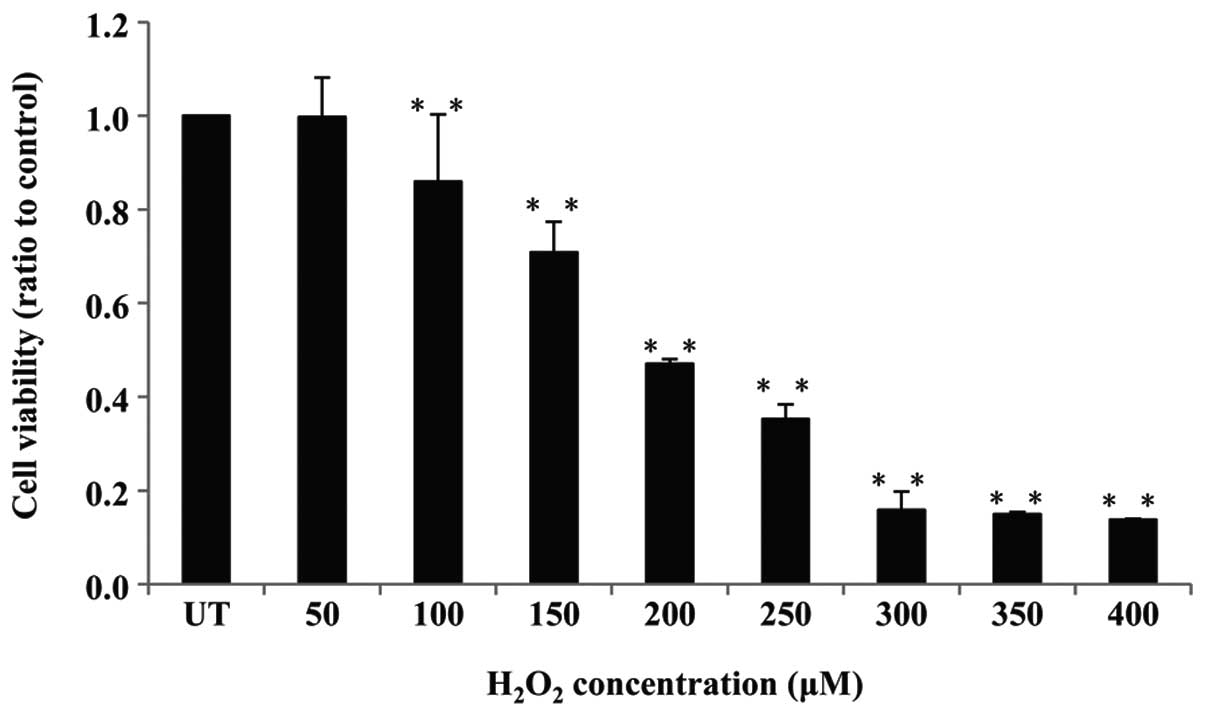
Cell viability in the presence and
absence of H2O2
Human melanocytes and H2O2
were used as a model for oxidative stress in order to investigate
cell viability using an MTT assay. Cells were treated with
different concentrations of H2O2 (0–400
μM) for 24 h, and cell viability was measured. As shown in
Fig. 1, the viability of
H2O2-treated cells significantly decreased in
a dose-dependent manner. Following treatment with 200 μM
H2O2, cell viability was reduced by ~50%,
this was considered to be the optimum concentration of
H2O2 and was used for subsequent
experiments.
Quercetin protects against
H2O2-induced cell death
In order to investigate the effect of quercetin on
H2O2-induced oxidative stress,
H2O2-treated cells were pretreated with
different concentrations of quercetin. In contrast to the
H2O2-treated cells, quercetin was observed to
have a minor, but not significant, protective effect at
concentrations of 6.25 and 12.5 μM; however, quercetin
concentrations of 25–200 μM exhibited significant protective
effects against H2O2-induced cell death
(Fig. 2). Therefore, 25 μM
quercetin was selected to be used for following experiments.
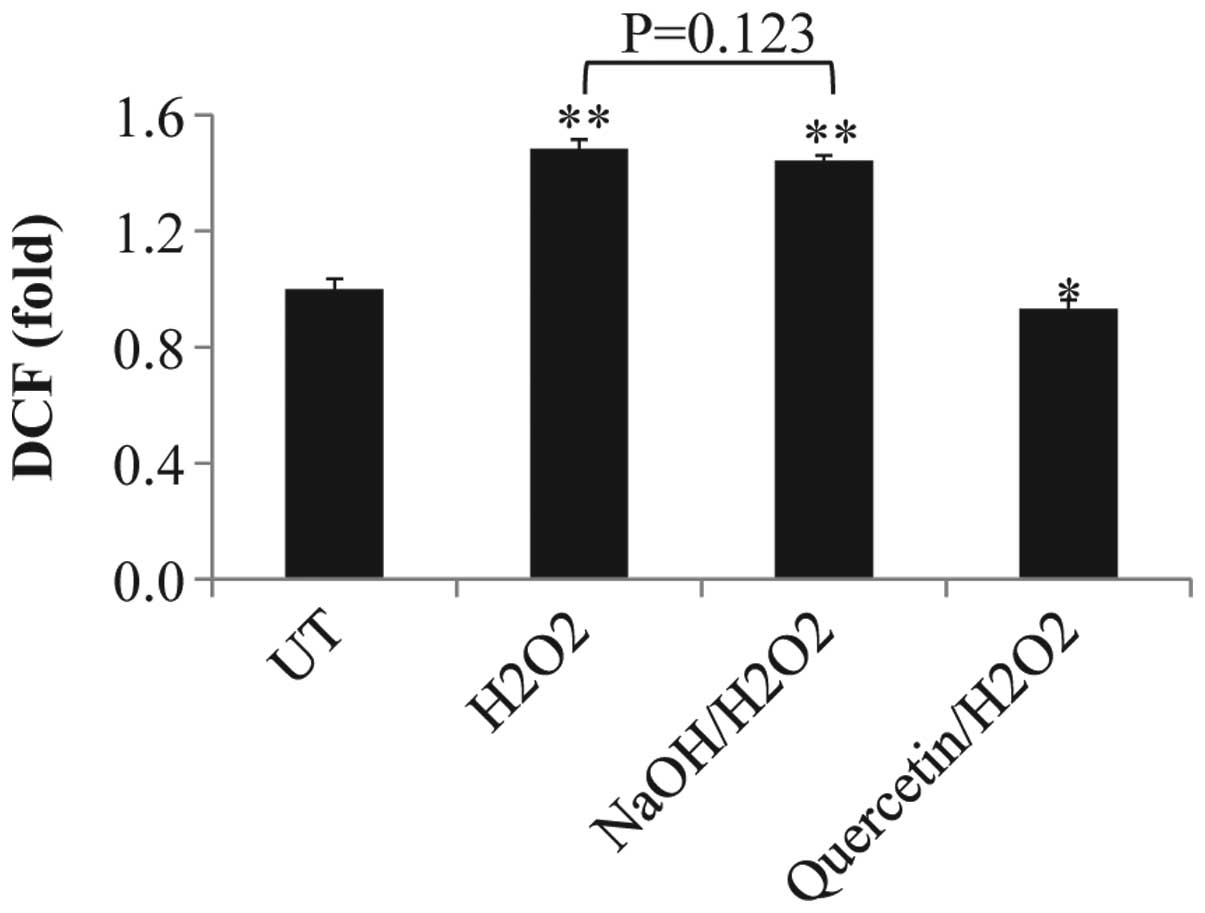
ROS production in melanocytes under
different conditions
ROS production was evaluated in melanocytes
following exposure to quercetin or NaOH in the presence or absence
of H2O2. As shown in Fig. 3, H2O2 and
NaOH/H2O2 treatments resulted in an 0.5 fold
increase in ROS production compared with that of the untreated
control levels (P<0.01); however, no significant difference was
identified between these two groups (P=0.123). In addition, ROS
levels in the quercetin/H2O2 group
demonstrated a significant decrease compared with those of the
untreated control group (P=0.022).
Effect of quercetin and
H2O2 on ER configuration
ER modality was analyzed using electron microscopy
in order to observe the effects of different conditions on
melanoctyes. As shown in Fig. 4,
dilated ER was observed in H2O2- and
NaOH/H2O2-treated cells. In untreated cells,
ER configuration was shown to be normal. Notably, normal ER
configuration was also observed in
quercetin/H2O2-treated cells. This therefore
suggested that quercetin was able to prevent
H2O2-induced ER dilation.
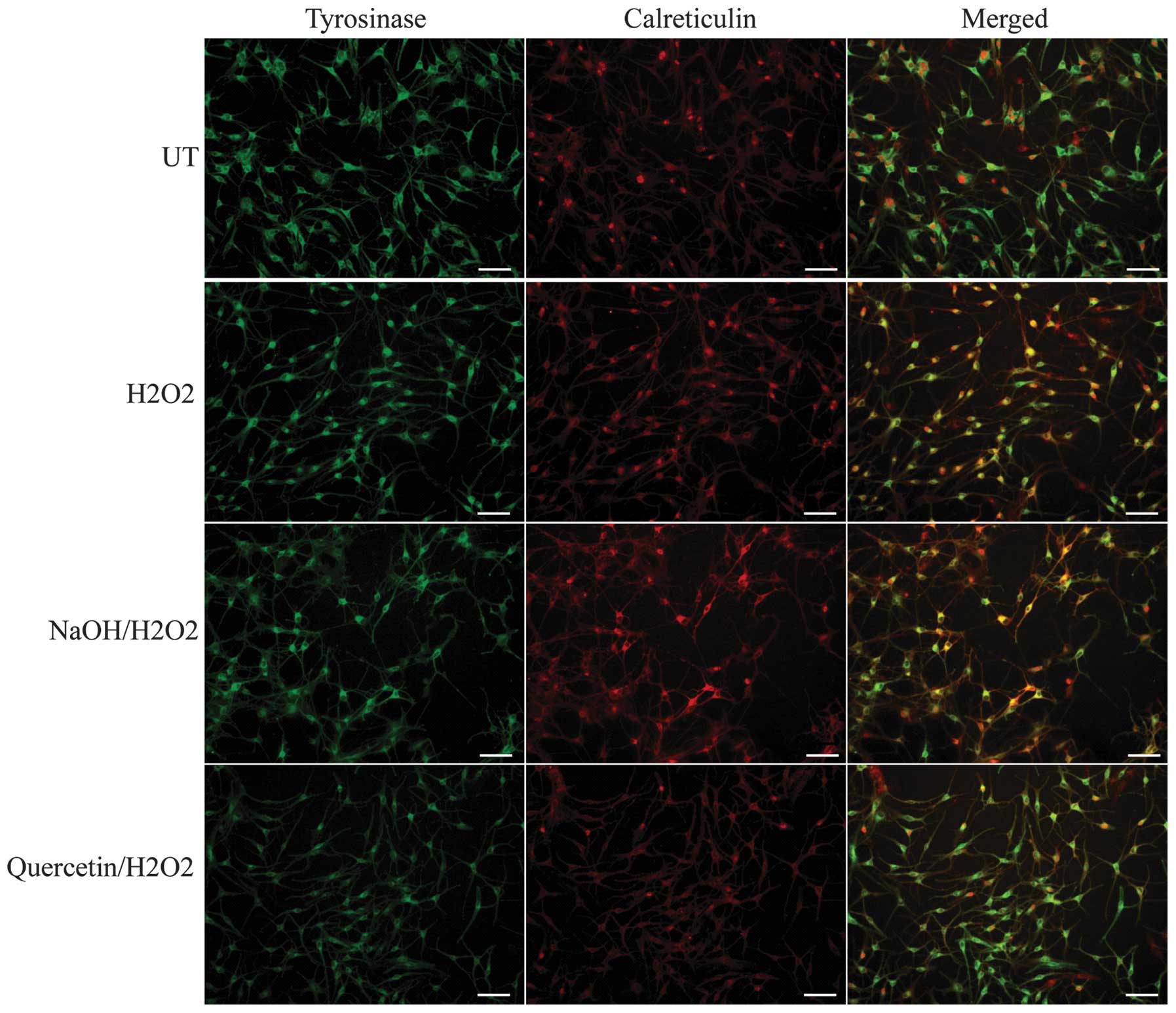
Co-localization of tyrosinase and
calreticulum
Confocal laser scanning microscopy was performed in
order to assess the co-localization of tyrosinase, the
rate-limiting enzyme for melanin synthesis, and calreticulin, an ER
marker protein. In untreated cells and
quercetin/H2O2-treated cells, tyrosinase
fluorescence was beyond the fluorescence marked by calreticulin,
which indicated that tyrosinase was effectively exported from the
ER. However, in H2O2- and
NaOH/H2O2-treated cells, the distribution of
tyrosinase fluorescence was pronounced in ER marked by
calreticulinin, which suggested that the export of tyrosinase from
the ER was disordered (Fig. 5);
this therefore indicated that H2O2 interfered
with the functional export of tyrosinase from the ER.
Quercetin attenuates
H2O2-induced inhibition of tyrosinase
expression
Western blot analysis was used to determined the
expression levels of tyrosinase in melanocytes. As shown in
Fig. 6, tyrosinase expression was
significantly increased in the quercetin/H2O2
group compared with that of the untreated group (P=0.046). By
contrast, tyrosinase expression was significantly decreased in the
H2O2 (P<0.01) and
NaOH/H2O2 groups (P<0.01) compared with
that of the control; however, no significant differences were
observed between these two groups (P=0.185). This therefore
indicated that quercetin attenuated the
H2O2-induced inhibition of tyrosinase
expression.
Discussion
H2O2 was previously
demonstrated to inhibit tyrosinase expression and melanocyte
viability; in addition, quercetin was found to protect melanocytes
from H2O2-mediated oxidative stress (13). This previous study primarily
focused on melanocyte viability; however, in the present study, the
effects of H2O2 on the tyrosinase export from
the ER and morphology of the ER, as well as the attenuation of
H2O2-induced oxidative stress by quercetin,
were further investigated.
The present study determined that 200 μM
H2O2 and 25 μM quercetin, with
incubation for 24 h were the optimum parameters for the experiments
performed. ROS levels in melanocytes of different treatment groups
were determined using DCFH-DA. ROS production was found to increase
1.5 fold in the H2O2-treated group compared
with that of the untreated group, whereas cells that underwent
quercetin/H2O2 treatment demonstrated
comparative ROS levels with those of the untreated group. This
therefore suggested that quercetin attenuated the
H2O2-induced increase in ROS levels. The
effects of H2O2 on ER modality in melanocytes
was observed using electron microscopy; markedly dilated ER were
observed in the H2O2-treated group, whereas
normal ER configuration was found in the
quercetin/H2O2-treated and untreated groups.
This therefore indicated H2O2 induced ER
dilation in melanocytes, which was prevented by quercetin
treatment. The ER is a vital and highly dynamic organelle present
in all eukaryotic cells; a multitude of parameters inside the cell
and in its microenvironment significantly influence the complex
functions of ER. Factors including the availability of glucose
(hypoglycemia), hyperthermia, calcium levels and the redox milieu,
impact and disturb the proper functioning of the ER, resulting in
ER stress, which results in improper protein folding in the lumen
of the ER (19,20). Pronounced dilation of the ER lumen
is a well established ultrastructural response to ER stress; under
which mammalian cells have been reported to expand their ER volume
several fold (21,22). The results of the present study
demonstrated that ER volume of melanocytes increased 2.02±0.07 fold
following H2O2 treatment, therefore
indicating that H2O2 disturbed the proper
function of the ER, while quercetin attenuated the effects of
H2O2.
Tyrosinase is the core enzyme that catalyzes
melanogenesis in melanocytes. Abnormalities in the
post-translational processing of tyrosinase have been implicated in
several depigmentation diseases (23). Stagnation of tyrosinase in ER was
found to be relevant to the phenotype of pigment loss in melanoma
(24). The dysfunctional
transportation of tyrosinase from the golgi to melanosomes leads to
different diseases, including generalized albinism types 2 and 4 as
well as Hermansky-Pudlak syndrome (25–27).
Therefore, in the present study, in order to evaluate protein
processing and transport in the ER of melanocytes in different
treatment groups, confocal laser scanning microscopy was performed
to assess the co-localization of tyrosinase and calreticulin. The
result demonstrated that tyrosinase and calreticulin expression
were both localized in the ER of H2O2- and
NaOH/H2O2-treated cells. A large amount of
tyrosinase was not observed at the endoplasmic reticulum marked by
calreticulin in untreated and
quercetin/H2O2-treated cells. These results
suggested that H2O2 hindered tyrosinase
export from the ER; however, quercetin pretreatment enabled cells
to maintain the effective export of tyrosinase from the ER.
Furthermore, comparative expression levels of tyrosinase were
observed in the quercetin/H2O2-treated and
untreated groups; however, tyrosinase expression was significantly
decreased in the H2O2 and
NaOH/H2O2 groups.
Numerous studies have reported that antioxidants may
protect melanocytes against oxidative stress; green tea extract was
found to protect cellular membranes against
t-butylhydroperoxide-induced oxidative damage (28), and a combination of vitamins C and
E demonstrated a protective effect against ultraviolet radiation
(29,30). Quercetin is found in a variety of
plant-based foods, including red onions, red grapes and a certain
berries (31). The potential
chemopreventive effects of quercetin have been attributed to
various mechanisms, including its antioxidative activity as well as
its capacity to inhibit enzymes that activate carcinogens
(resulting in the modification of signal transduction pathways) and
interact with and regulate cell receptors and other proteins
(32). The results of the present
study demonstrated that quercetin protected melanocytes from the
effects of H2O2 on the morphology of ER,
tyrosinase export from the ER and tyrosinase expression.
In conclusion, the results of the present study
showed that H2O2 induced the dilation of ER
lumina and hindered the functional export of tyrosinase from the
ER, while quercetin attenuated these effects induced by
H2O2. To the best of our knowledge, the
present study provided the first evidence that
H2O2 has an important role on the ER
morphology of melanocytes and functional export of tyrosinase from
ER. These results may aid in elucidating the potential effect of
antioxidants on the ultrastructure of melanocytes.
Acknowledgments
The present study was supported partly by grants
from the National Natural Science Foundation of China (grant nos.
30800563 and 81071294), the Zhejiang Provincial Natural Science
Foundation of China (grant nos. Y2101132, Z2100973, LY12H11008 and
LY12H11009), the Science Technology Department of Zhejiang Province
(grant no. 2013C33093) and the Hangzhou Science and Technology
project (grant no. 20092133W04). Additionally, the authors would
like to acknowledge the financial support from the State Clinical
Key Specialty Construction Project and Zhejiang Provincial Program
for the Cultivation of High-level Health talents.
References
|
1
|
Taïeb A and Picardo M; VETF Members: The
definition and assessment of vitiligo: a consensus report of the
Vitiligo European Task Force. Pigment Cell Res. 20:27–35. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Le Poole IC, Das PK, van den Wijngaard RM,
Bos JD and Westerhof W: Review of the etiopathomechanism of
vitiligo: a convergence theory. Exp Dermatol. 2:145–153. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Schallreuter KU: Successful treatment of
oxidative stress in vitiligo. Skin Pharmacol Appl Skin Physiol.
12:132–138. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Maresca V, Roccella M, Roccella F, et al:
Increased sensitivity to peroxidative agents as a possible
pathogenic factor of melanocyte damage in vitiligo. J Invest
Dermatol. 109:310–313. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Dell’Anna ML, Maresca V, Briganti S, et
al: Mitochondrial impairment in peripheral blood mononuclear cells
during the active phase of vitiligo. J Invest Dermatol.
117:908–913. 2001. View Article : Google Scholar
|
|
6
|
Schallreuter KU, Wood JM and Berger J: Low
catalase levels in the epidermis of patients with vitiligo. J
Invest Dermatol. 97:1081–1085. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sravani PV, Babu NK, Gopal KV, et al:
Determination of oxidative stress in vitiligo by measuring
superoxide dismutase and catalase levels in vitiliginous and
non-vitiliginous skin. Indian J Dermatol Venereol Leprol.
75:268–271. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Salem MM, Shalbaf M, Gibbons NC, et al:
Enhanced DNA binding capacity on up-regulated epidermal wild-type
p53 in vitiligo by H2O2-mediated oxidation: a
possible repair mechanism for DNA damage. FASEB J. 23:3790–3807.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Giovannelli L, Bellandi S, Pitozzi V, et
al: Increased oxidative DNA damage in mononuclear leukocytes in
vitiligo. Mutat Res. 556:101–106. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Westerhof W and d’Ischia M: Vitiligo
puzzle: the pieces fall in place. Pigment Cell Res. 20:345–359.
2007.PubMed/NCBI
|
|
11
|
Schallreuter KU, Elwary SM, Gibbons NC,
Rokos H and Wood JM: Activation/deactivation of
acetylcholinesterase by H2O2: more evidence
for oxidative stress in vitiligo. Biochem Biophys Res Commun.
315:502–508. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Jeong YM, Choi YG, Kim DS, et al:
Cytoprotective effect of green tea extract and quercetin against
hydrogen peroxide-induced oxidative stress. Arch Pharm Res.
28:1251–1256. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Nagata H, Takekoshi S, Takeyama R, Homma T
and Yoshiyuki Osamura R: Quercetin enhances melanogenesis by
increasing the activity and synthesis of tyrosinase in human
melanoma cells and in normal human melanocytes. Pigment Cell Res.
17:66–73. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Dell’Anna ML, Mastrofrancesco A, Sala R,
et al: Antioxidants and narrow band-UVB in the treatment of
vitiligo: a double-blind placebo controlled trial. Clin Exp
Dermatol. 32:631–636. 2007. View Article : Google Scholar
|
|
15
|
Boissy RE, Liu YY, Medrano EE and Nordlund
JJ: Structural aberration of the rough endoplasmic reticulum and
melanosome compartmentalization in long-term cultures of
melanocytes from vitiligo patients. J Invest Dermatol. 97:395–404.
1991. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Guan C, Lin F, Zhou M, et al: The role of
VIT1/FBXO11 in the regulation of apoptosis and tyrosinase export
from endoplasmic reticulum in cultured melanocytes. Int J Mol Med.
26:57–65. 2010.PubMed/NCBI
|
|
17
|
Jinbo L, Zhiyuan L, Zhijian Z and WenGe D:
Olfactory ensheathing cell-conditioned medium protects astrocytes
exposed to hydrogen peroxide stress. Cell Mol Neurobiol.
33:699–705. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hong WS, Hu DN, Qian GP, McCormick SA and
Xu AE: Ratio of size of recipient and donor areas in treatment of
vitiligo by autologous cultured melanocyte transplantation. Br J
Dermatol. 165:520–525. 2011.PubMed/NCBI
|
|
19
|
Malhotra JD and Kaufman RJ: The
endoplasmic reticulum and the unfolded protein response. Semin Cell
Dev Biol. 18:716–731. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ron D and Walter P: Signal integration in
the endoplasmic reticulum unfolded protein response. Nat Rev Mol
Cell Biol. 8:519–529. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hartley T, Siva M, Lai E, et al:
Endoplasmic reticulum stress response in an INS-1 pancreatic
beta-cell line with inducible expression of a folding-deficient
proinsulin. BMC Cell Biol. 11:592010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zuber C, Fan JY, Guhl B and Roth J:
Misfolded proinsulin accumulates in expanded pre-Golgi
intermediates and endoplasmic reticulum subdomains in pancreatic
beta cells of Akita mice. FASEB J. 18:917–919. 2004.PubMed/NCBI
|
|
23
|
Toyofuku K, Wada I, Valencia JC, Kushimoto
T, Ferrans VJ and Hearing VJ: Oculocutaneous albinism types 1 and 3
are ER retention diseases: Mutation of tyrosinase or Tyrp1 can
affect the processing of both mutant and wild-type proteins. Faseb
J. 15:2149–2161. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Halaban R, Cheng E, Zhang Y, et al:
Aberrant retention of tyrosinase in the endoplasmic reticulum
mediates accelerated degradation of the enzyme and contributes to
the dedifferentiated phenotype of amelanotic melanoma cells. Proc
Natl Acad Sci USA. 94:6210–6215. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Berson JF, Frank DW, Calvo PA, et al: A
common temperature-sensitive allelic form of human tyrosinase is
retained in the endoplasmic reticulum at the nonpermissive
temperature. J Biol Chem. 275:12281–12289. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Chiang PW, Oiso N, Gautam R, et al: The
Hermansky-Pudlak syndrome 1 (HPS1) and HPS4 proteins are components
of two complexes, BLOC-3 and BLOC-4, involved in the biogenesis of
lysosome-related organelles. J Biol Chem. 278:20332–20337. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kushimoto T, Valencia JC, Costin GE, et
al: The Seiji memorial lecture: the melanosome: an ideal model to
study cellular differentiation. Pigment Cell Res. 16:237–244. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Saffari Y and Sadrzadeh SM: Green tea
metabolite EGCG protects membranes against oxidative damage in
vitro. Life Sci. 74:1513–1518. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Quevedo WC Jr, Holstein TJ, Dyckman J and
McDonald CJ: The responses of the human epidermal melanocyte system
to chronic erythemal doses of UVR in skin protected by topical
applications of a combination of vitamins C and E. Pigment Cell
Res. 13:190–192. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Smit N, Vicanova J, Cramers P, Vrolijk H
and Pavel S: The combined effects of extracts containing
carotenoids and vitamins E and C on growth and pigmentation of
cultured human melanocytes. Skin Pharmacol Physiol. 17:238–245.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Bischoff SC: Quercetin: potentials in the
prevention and therapy of disease. Curr Opin Clin Nutr Metab Care.
11:733–740. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Murakami A, Ashida H and Terao J:
Multitargeted cancer prevention by quercetin. Cancer Lett.
269:315–325. 2008. View Article : Google Scholar : PubMed/NCBI
|