Introduction
Human tumor necrosis factor (hTNF)-α is a
multifunctional cytokine produced by activated macrophages. TNF-α
is known to have potent antitumor properties in vivo and
in vitro (1), and thus has
potential clinical applications in cancer treatment. However, it is
unstable with a short half-life and significant side effects in
vivo, which greatly restricts its use in a clinical
setting.
Urokinase-type plasminogen activator (uPA) is a
serine protease. It is able to activate the conversion of
plasminogen to plasmin, which is then involved in triggering the
fibrinolysis of a variety of matrix proteins. A number of studies
have suggested (2–4) that the over-expression of uPA is
important in the evolution of malignant tumors, and that it is
closely correlated with tumor cell proliferation, angiogenesis,
tumor invasion, metastasis and the inhibition of apoptosis. uPA is
highly expressed in tumor tissues, but low or no expression is
observed in normal tissues. Furthermore, it displays a high
affinity for the specific substrate, plasminogen. Therefore, hTNF-α
mediated by uPA may exhibit a decrease in the toxicity compared
with wild-type hTNF-α and also solve the problem of tumor
targeting, as it is only active when cleaved by tumor-expressing
urokinase.
The current study was conducted in order to obtain
preliminary data on the effect of a novel hTNF-α fusion protein on
tumor suppression, its mechanism of action and its influence on
liver and renal function.
Materials and methods
Expression vectors, cell lines and
animals
Escherichia coli with Rosetta2 was purchased
from Novagen (Madison, WI, USA). The S180 mouse cells were provided
by the laboratory of Guangdong Medical College (Donguang, China).
Kunming mice, (body weight, 28–30 g); 18:18, male:female, were
obtained from The Animal Center of Guangdong Medical College.
Animals were housed in a temperature-controlled room (24°C) with a
12 h/12 h cycle of light and dark and were allowed ad
libitum access to food and water throughout the experiment. The
study was approved by the ethics committee of the Experimental
Animal Center of Guangdong Medical College (Zhanjiang, China).
Primary instruments
Vertical electrophoresis, electrophoresis apparatus
and a 450 enzyme-labeled meter were purchased from Bio-Rad
(Hercules, CA, USA). An HWI thermostatic water tank was purchased
from Beijing Medical Equipment Factory (Beijing, China). An MAIS
pathological color image analyzer, a microscope (CX31; Olympus
Corp., Tokyo, Japan) and any relevant software (MAIS image-Pro
System, version 2000 P2; Jiangsu Software Technology Co., Ltd.,
Zhejiang, China) were provided by the Department of Pathology,
Guangdong Medical College. All other frequently-used apparatus was
provided by the Biochemical Laboratory of Guangdong Medical
College.
Drugs and reagents
The fusion protein was extracted in concentrations
of 0.1, 0.2 and 0.3 μg/μl. BugBuster Protein
Extraction kits were obtained from Novagen. uPA assay kits,
terminal deoxynucleotidyl transferase-mediated dUTP nick end
labeling (TUNEL) kits, bcl-2 immunohistochemical assay kits, bax
immunohistochemical assay kits and vascular endothelial growth
factor (VEGF) immunohistochemical kits were all obtained from
Shanghai Bogu Biological Technology Co., Ltd. (Shanghai, China).
Other reagents were obtained from the Biochemical Laboratory of
Guangdong Medical College.
Plasmid construction
The plasmid pET-42a(+) foldon-uPA-hTNF-α was made by
cloning the foldon sequence, the uPA distinguishing sequence and
the wild-type hTNF-α sequence [with eight amino acids (aa) deleted
in the N terminal] into a pET-42a(+) vector (Zhejiang Tianhang
Biological Technology Co., Ltd., Hangzhou, China). Using two 5′-end
modified PCR (that is adding a synthetic nucleotide sequence to the
5′ end), the foldon sequence, uPA distinguishing sequence and
wild-type hTNF-α sequence (with eight aa deleted in the N terminal)
of a total 572bp was cloned into pET-42a between the PshA and Sac
restriction enzyme sites and thus the pET-42a (+)-foldon-uPA-hTNF
recombinant plasmid was obtained. It carried a GS-foldon sequence
(27 aa), a GS-uPA distinguishing sequence (7 aa) and an hTNF-α
sequence with 8 aa deleted from the N terminal. The GS-foldon
sequence encodes 29 aa, GYIPEAPRDGQAYVRKDGEWVLLSTFL and each end of
the GS has a connecting function to foldon sequence and a uPA
distinguishing sequence. The uPA distinguishing sequence was Pro
Gly Arg↓Val Ala Lys. ↓ Represents the cleavage site of uPA.
Expression, extraction, purification and
deblocking of the fusion protein
Rosetta2 (DE3; Zhejiang Tianhang Biological
Technology Co., Ltd.) with the correct sequence was grown in luria
broth (LB) agar containing kanamycin (Kana) and chloramphenicol
(Cam) at an appropriate concentration to obtain the monoclone. The
monoclone was grown overnight at 37°C in 3 ml LB liquid medium
containing 30 mg/ml Kana and 34 mg/ml Cam. The culture was then
diluted into 3 litres of medium containing Kana, and grown at 37°C
until the culture reached an A600 of 0.5 (MCF
Nephelometer wi93008; Dongxiyi Technology Co., Ltd., Beijing,
China). The culture was diluted to a final concentration of 0.8
mmol/l by adding IPTG (Dongguan Mag Biotechnology Science Co.,
Ltd., Dongguan, China) at 20°C and agitating at 160 rpm for 24 h.
The cells were collected by centrifugation at 3,000 × g for 15 min
at 4°C, after the culture had been on ice for 10 min. The collected
cells were suspended in phosphate-buffered saline (PBS), then
removed by centrifugation at 3,000 × g for 15 min at 4°C. The cells
were weighed and treated according to the manufacturer’s
instructions of the BugBuster GST-Bind Purification kits) in order
to extract the soluble protein. The fusion protein was removed from
glutathione S-transferase (GST) by Factor Xa (Novagen), and the
target protein was obtained from the collected elution. The fusion
protein levels were analyzed by separating the proteins with 6%
SDS-PAGE, followed by staining with coomassie brilliant blue. The
concentration of fusion protein without the GST tag was detected by
bicinchonininc acid and the pick-up rate was calculated: Pick-up
rate = fusion protein rate/bacteria rate. The deblocked fusion
protein was the product of uPA acting on the uPA dsitinguishing
sequence in the rhTNF within a Tris-HCl buffer of pH 7.5.
Cytotoxic activity of fusion protein on
L929 cells by an
3-(4,5-dimethylthiaxol-2-yl)-2-5-diphenyltetrazolium (MTT)
assay
Cytotoxic activity was estimated by an MTT assay, as
described previously (5). The
suspension was adjusted with L929 cells in logarithmic growth to a
concentration of 2×105 cells/ml, and added into a
96-well plate (0.1 ml per well) for 24 h, after which the
supernatent was removed. The rhTNF-α fusion protein without the GST
tag, and the deblocked rhTNF-α fusion protein were each diluted
into 1, 10, 100 and 1,000 nmol/l with RPMI-1640 (Gibco-BRL,
Invitrogen Life Technologies, Carlsbad, CA, USA). The above
dilution, the negative control and the standard TNF-α positive
control were added into the wells for another 24 h. MTT (20
μl, 5 mg/ml) was then added to the wells for 4 h at 37°C,
after which the supernatent was again removed. The crystals were
fully dissolved by adding 200 μl dimethyl sulfoxide to each
well for 0.5 h at 37°C. Finally, the absorbance was determined at
570 nm with a 450 enzyme-labeled meter (GF-M3000; Aloka Medical
Equipment Co., Ltd., Tokyo, Japan) and the percentage of dead cells
at different concentrations of fusion protein was calculated. The
specific activity (activity of TNF-α per mg protein) of the fusion
protein and the deblocked protein, based on the definition of an
active unit (U) as the concentration which is capable of killing
50% of cells.
Establishment of the animal model, and
grouping and injection
Frozen S180 cells were recovered and inoculated into
the abdomens of four healthy mice (0.1 ml per mouse). The mice were
sacrificed by cervical dislocation after seven days and the ascites
were extracted under sterile conditions. Ascitic cells were diluted
down to 1.8×107/ml with sterile saline. The cell
suspension was injected into 32 Kunming mice (0.2/ml per mouse;
subcutaneously into the axilla) in order to generate a solid tumor
model. The following day, the S180 tumor-bearing mice were randomly
divided into four groups after weighing. The groups were: Saline
control group (group S); low dose group (group L); middle dose
group (group M) and high dose group (group H). Saline and saline
plus 0.1, 0.2 or 0.3 μg/μl of fusion protein (0.5 ml)
were administered intraperitoneally every other day for 11 days
(i.e. six times in total).
Antitumor effect of the fusion protein
and its effect on liver and kidney function
Blood was drawn by removing eyeballs the day
following drug withdrawl. The serum of each mouse was preserved at
−20°C for measurement of liver function, assessed by levels of
alanine transaminase (ALT) and albumin (ALB), and kidney function,
assessed by levels of creatinine (Cr) and blood urea nitrogen
(BUN). The tumor, liver and kidneys were surgically removed and
weighed after measurement of mouse weight, and the liver and kidney
indices were calcu lated: Liver index (mg/g) = (liver weight/body
weight) × 1,000; kidney index(mg/g) = (kidney weight/body weight) ×
1,000. The percentage reduction in mean tumor weights between the
groups was assessed. The organs were fixed in formalin for
analysis.
Determination of uPA in serum
Serum levels of uPA were determined by an
enzyme-linked immunosorbent assay (ELISA). ELISA was conducted
according to the manufacturer’s instructions for uPA. A standard
curve was drawn to calculate the concentration of uPA in each group
and the reduction rate in each group. The reduction rate of uPA (%)
= [1−(mean uPA content of treatment group/mean uPA content of
saline control group)] × 100%.
Expression of bcl-2, bax and VEGF by a
streptavidin-biotin complex (sABC) assay
An immunohistochemical method was used to observe
levels of expression of bcl-2, bax and VEGF in tumor tissues in
mice from each group (6). An
immunohistochemical sABC staining method was adopted, and the
procedures were followed according to the manufacturer’s
instructions. Briefly, tissues were fixed by adding neutral
formaldehyde-buffered solution into the tissue, prior to routine
paraffin imbedding. The slides were dried following 1 h soak in
poly-L-lysine slide anti-creep solution, and tissues were sliced
into 4-μm slices and routinely de-waxed prior to being
incubated in 10% formaldehyde-H2O2 for 10 min
at room temperature. The tumor tissue slices were blocked with goat
serum confining liquid for 20 min. Rabbit monoclonal anti-bcl-2
(Sigma-Aldrich), anti-bax (Sigma-Aldrich) and anti-VEGF (Santa Cruz
Biotechnology, Inc., Dallas, TX, USA) antibodies were incubated
overnight at 4°C (working concentration, 1:2,000). The secondary
antibody (1:2000; biotinylated goat anti rabbit IgG; Santa Cruz
Biotechnology, Inc.) was incubated at 3°C for 20 min. sABC was
added (Shanghai Bogu Biological Technology Co., Ltd., Shanghai,
China) and incubated at 37°C for 20 min prior to
DAB-H2O2 coloration. Hematoxylin staining was
performed. A negative control was established, with PBS replacing
the first antibody, in order to rule out non-specific staining.
Positive staining of bcl-2, bax and VEGF was yellowish-brown or
brown. The positive area was determined from approximately five
visual fields in each section at ×100 magnification by an MAIS
pathological color image analyzer.
Apoptotic cell determination by a TUNEL
assay
The TUNEL method was used to observe the percentage
of apoptotic cells in tumor tissues of mice from each group
(7). Briefly, tissues were sliced
and routinely de-waxed. They were then washed in 3%
H2O2 three times, digested by the protease,
dissolved in Tris/HCl, pH 7.4–8.0 at 37°C, and then washed again.
Next, 1 μl terminal deoxynucleotidyl transferase, 1
μl dig-dUTP (Shanghai Bogu Biological Technology Co., Ltd.)
and 18 μl mark buffer were added and then mixed. The
redundant liquid was removed and 20 μl mark liquid was added
and incubated in a moisture box (Shanghai Yuejin Technology Co.,
Ltd., Shanghai, China) at 37°C for 2 h. The mixture was then washed
in PBS three times. The sample was incubated with enzyme labeled
anti-fluorescein antibody at 37°C for 30 min, and then washed three
times again with PBS. DAB coloration was performed for 10–30 min
and the sample was then washed with water. Finally the samples were
stained with hematoxylin, dehydrated, dehydrated, soaked in
dimethylbenzene and mounted. The apoptotic cells were stained
yellowish-brown or brown. TUNEL-positive cells were counted on five
randomly selected visual fields at ×400 magnification, and the
apoptotic index was calculated.
Statistical analysis
The results are presented as the mean ± the standard
error. An analysis of variance test was conducted using SPSS 19.0
(IBM, Armonk, NY, USA) for Windows. P<0.05 and P<0.01 were
considered to indicate a statistically significant difference.
Results
Plasmids contain the foldon, uPA
substrate and hTNF-α mutant gene sequences
The two sequencing assays confirmed that a fusion
gene sequence with a length of 572 bp was inserted between the two
restriction sites of PshA↓ and Sac↓ in the pET-42a carrier,
including the foldon sequence, the uPA substrate sequence and the
hTNF-α mutant gene, which was consistent with the intended
design.
Urokinase specifically and effectively
deblocks the fusion protein
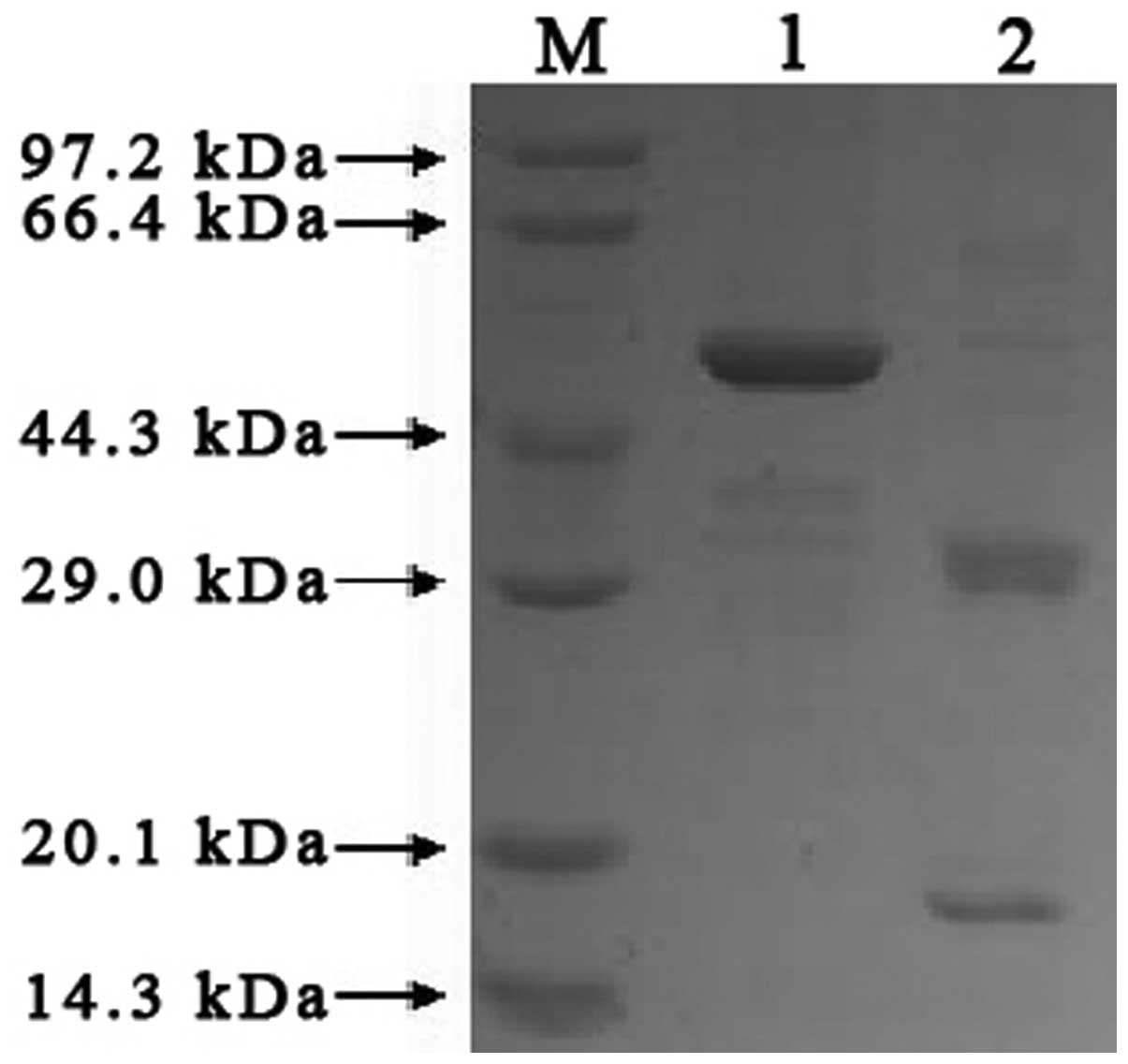
Based on the results of the SDS-PAGE gray scale scan
and protein concentration measurement, 2.33 mg target protein
without a GST tag was obtained from 1 g (wet weight) of
E.coli. Protein expression was induced by IPTG, and then the
protein was purified by GST•Bind™ before the GST tag was removed by
factor Xa. Theoretically, after cleavage by urokinase, the rhTNF
protein should change from the original band (of 49 kDa) to two
bands (of 17.5 kDa and 31.5 kDa). The results of the SDS-PAGE are
shown in Fig. 1. By comparing with
standard protein controls and the undigested rhTNF protein, two
protein bands in the ranges 14.3–20.1 kDa and 29.0–44.3 kDa were
observed. Given the predicted molecular weight of the cleaved
protein, the results suggest that urokinase had an effective and
specific deblocking effect on the fusion protein.
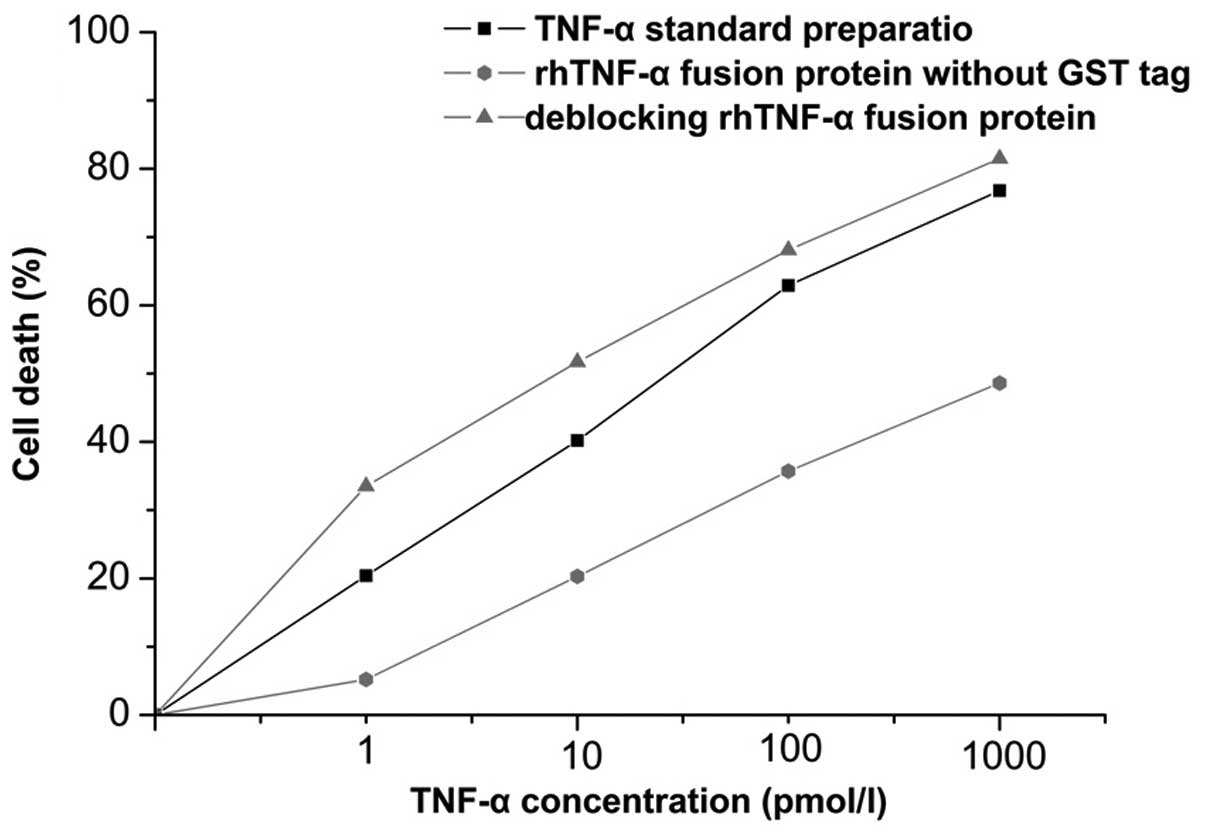
Cytotoxic activity of the deblocked
fusion protein is significantly higher than that of the fusion
protein
In order to examine the effects of the deblocked
fusion protein, the cytotoxic activity of tumor cells of the fusion
protein and active fusion protein were detected by L929 cells. As
shown in Fig. 2, after 24 h
incubation there was an effect on cell apoptosis, which increased
in a dose-dependent manner with the level of the deblocked fusion
protein. By contrast, the fusion protein showed no significant
cytotoxic activity on L929 cells, as the receptor binding site had
been blocked. Based on the regression equation with the
concentration values (expressed logarithmically) as the independent
variable and the percentage of cell death as the dependent
variable, and taking a cytotoxic activity of 50% as an active unit
(U), the specific activity of the fusion protein was calculated as
1.82×105 U/mg, and that of the deblocked fusion protein
as 6.30×105 IU/mg.
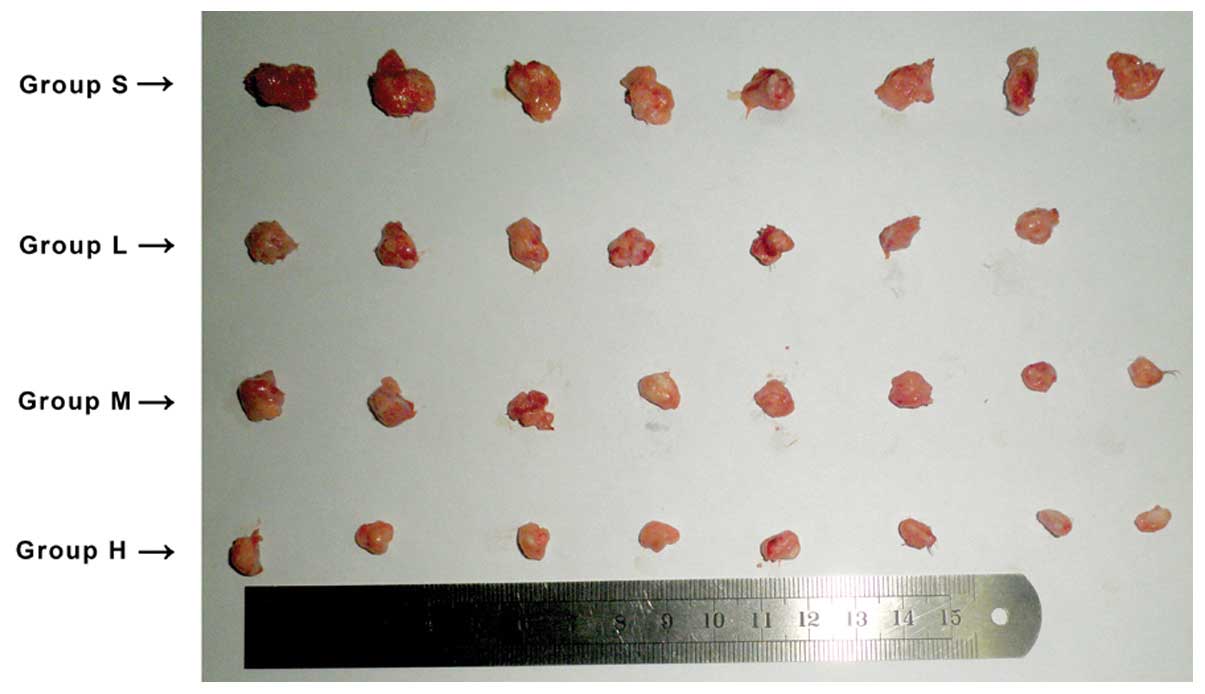
Injection of the fusion protein into the
abdominal cavity reduces tumor size in a dose-dependent manner
S180 mouse cells were selected to generate solid
tumor models, as they are an established tumor model (8,9). In
the saline control group, tumors growth was first observed four
days after being vaccinated with the S180 cells. By contrast,
growth in the high dose group was delayed until the seventh day and
thereafter grew more slowly. One mouse in the low dose group died,
and the autopsy showed this was most likely to have been due to
infection. Tumors from mice in the control group were notably
larger than those from mice in the groups treated with the fusion
protein. The control group tumors were also harder to extract and
showed evidence of infiltrative growth. By contrast, tumors in the
fusion protein groups were easier to remove and solitary. The size
and weight of solid tumors from each group are shown in Fig. 3 and Table I. The study showed that the fusion
protein inhibited growth of S180 tumors in a mouse model, and that
the percentage difference in mean tumor weight increased with
increasing doses of fusion protein compared with the control group,
rising to 75.61% in the high dose group.
 | Table ITumor inhibition rate of S180 bearing
mice. |
Table I
Tumor inhibition rate of S180 bearing
mice.
| Group | Number of mice | Fatalities | Tumor weight (g) | t-value | P-value | Reduction in mean
tumor growth (%) |
|---|
| S | 8 | 0 | 0.529±0.225 | – | – | – |
| L | 8 | 1 | 0.289±0.056 | 2.737 | 0.016 | 45.37 |
| M | 8 | 0 | 0.198±0.078 | 4.372 | 0.002 | 62.57 |
| H | 8 | 0 | 0.129±0.029 | 4.967 | <0.001 | 75.61 |
Levels of serum uPA decrease following
fusion protein treatment in a dose-dependent manner
With standard concentration on the vertical axis and
optical density on the horizontal axis on logarithmic graph paper,
a standard curve was plotted and the serum uPA of each group was
calculated. In the high-dose group, the serum uPA level was
1.035±0.062 ng/ml, which was reduced by 29.40% compared with the
saline control group. uPA levels decreased with increasing doses of
the fusion protein. All results were statistically significant
(P<0.05), as listed in Table
II.
| Table IIuPA content in serum. |
Table II
uPA content in serum.
| Group | uPA content
(ng/ml) | t-value | P-value | Reduction rate
(%) |
|---|
| S | 1.466±0.134 | – | – | – |
| L | 1.324±0.175 | 1.701 | 0.043 | 9.69 |
| M | 1.225±0.137 | 3.516 | 0.003 | 16.44 |
| H | 1.035±0.062 | 8.173 | <0.001 | 29.40 |
Fusion protein exhibits a protective
effect on the liver at all doses, and a mildly toxic effect on the
kidney at higher doses
Liver and kidney indices were calculated based on
liver, kidney and body weights. As shown in Table III and Fig. 4, compared with group S, the liver
and kidneys indices of tumor-bearing mice in each fusion protein
group were not significantly different (P>0.05). However, levels
of ALT and ALB in the fusion protein groups were significantly
different compared with the control group (P<0.05). ALT
decreased, whilst ALB increased as the dose of fusion protein
increased. Levels of BUN and Cr levels in groups L and M were not
significantly different from that in group S. However, BUN and Cr
were significantly higher in group H (9.583±1.457 and 18.29±3.352
mmol/l, respectively) compared with that in the control group
(7.099±1.710 and 13.000±2.944 mmol/l, respectively).
| Table IIIEffects of fusion protein on the liver
and kidney. |
Table III
Effects of fusion protein on the liver
and kidney.
| Group | Liver index (%) | Kidney index (%) | ALT (U/l) | ALB (g/l) | BUN (mmol/l) | Cr (umol/l) |
|---|
| S | 6.516±0.867 | 1.580±0.286 | 54.290±9.268 | 11.586±1.753 | 7.099±1.710 | 13.000±2.944 |
| L | 6.383±0.915 | 1.501±0.155 | 48.710±9.759a | 13.886±1.617a | 7.096±0.531 | 15.57±1.988 |
| M | 6.430±1.208 | 1.484±0.253 | 46.83±8.208a | 14.533±1.136b | 7.300±0.933 | 15.17±3.312 |
| H | 6.369±0.700 | 1.506±0.126 | 34.71±6.370b |
14.657±1.693b | 9.583±1.457a | 18.29±3.352b |
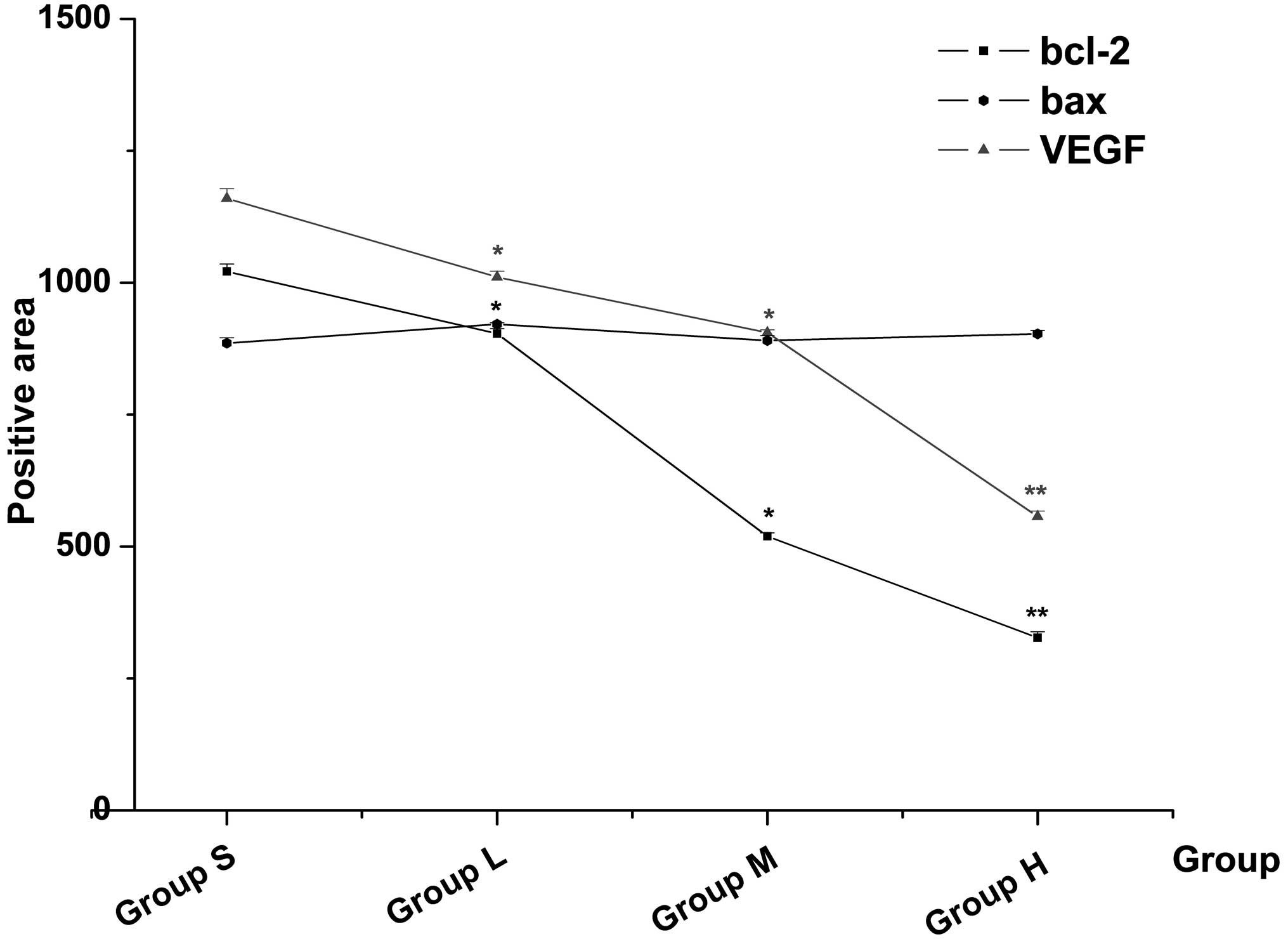
Immunohistochemical results of bcl-2, bax
and VEGF expression
The results of the immunohistochemistry are shown in
Table IV and Figs. 5 and 6. The expression of bcl-2 and VEGF in the
fusion protein groups decreased as the dose increased (all
P<0.05). The positive areas of bcl-2 and VEGF expression in
group H were 327.01±11.36 and 556.74±10.58, respectively. These
were significantly lower than that in group S (P<0.01). Notably,
the expression of bax was not significantly different compared with
that in group S (P>0.05).
| Table IVExpression of bcl-2, bax and VEGF in
tumor tissue. |
Table IV
Expression of bcl-2, bax and VEGF in
tumor tissue.
| Group | Bcl-2 | bax | VEGF |
|---|
| S | 1021.21±14.32 | 885.52±10.23 | 1160.01±18.26 |
| L | 903.33±9.59a | 921.26±2.65 |
1010.55±11.25a |
| M | 519.59±6.47a | 890.49±9.26 | 905.38±5.36a |
| H |
327.01±11.36b | 903.24±6.24 |
556.74±10.58b |
Apoptosis is increased by fusion protein
treatment compared with control
The apoptotic nuclei in tumor cells from each group
were detected by the TUNEL assay (Fig.
7), as described previously (10). The apoptotic index of group S was
4.50±1.05%. Those of groups L and H, were 34.01±5.20 and
71.16±3.44% respectively, and were significantly higher than in
group S (P<0.01). The apoptotic index of group M was
24.46±4.34%, which was also significantly different compared with
group S (P<0.05). Although the apoptotic index was lower in
group M than in group L, there was a general dose-dependent
increase in apoptosis following fusion protein treatment (Fig. 8).
Discussion
With increasing understanding of the molecular
biology of cancer in recent years, targeted therapy has become an
increasingly important part of cancer therapy, where significant
progress has been made in the treatment of various types of tumors
(11–12). To date, research on hTNF-α
structural transformation, has suggested that using molecular
methods to transform TNF-α molecular structure may be of great
significance in the development of cancer therapies (13–14).
A novel tumor-targeting transformation strategy involving hTNF-α
was adopted in this study. Within each hTNF-α monomer, the eight
amino-acid residues at the N-terminal are replaced by a urokinase
substrate sequence, which is coupled with foldon domain sequences
from bacteriophage T4 fibritin at the N-terminal. Theoretically,
the foldon sequence is used to block the N-terminal at the bottom
of the molecular structure of hTNF-α, and the blocked N-terminal is
able to autonomously form trimers, following those formed by whole
hTNF-α. As a result, the enzyme substrate sequence is involved in
deblocking the receptor binding site for hTNF-α. That is, the
blocked fusion protein does not exhibit the normal activity of
hTNF-α. However, when recombinant hTNF-α reaches the tumor site,
where uPA is highly expressed, this enzyme will cleave the enzyme
substrate sequence, thus cutting off the triple helix foldon
structure along with parts of the uPA substrate sequence. The
receptor binding site is then exposed, resulting in activated
hTNF-α, which is then involved in mediating tumor cell
apoptosis.
A recombinant plasmid containing hTNF-α, uPA and
foldon sequences, which was transformed into Rosetta2 (DE3) under
specified conditions was produced. The plasmid was induced to
express soluble fusion protein by Kana and Cam, which was then
purified by (GST) resin purification kit. The pick-up rate of
fusion protein was 0.23% and the purification rate was 93%.
Additionally, the analysis of fusion protein digestion by uPA
showed that urokinase had an efficient and specific deblocking
effect on this fusion protein. Through the analysis on the
cytotoxic activity of the fusion protein, it was observed that the
deblocked rhTNF-α fusion protein significantly increased cell death
compared with the inactive fusion protein. Moreover, the percentage
of L929 cell death was positively correlated with the level of the
deblocked protein, while the fusion protein had no significant
biological activity. This was consistent with the theory of
transformation. The specific activities of rhTNF-α and deblocked
fusion protein were measured. They were 1.82×105 and
6.30×105 IU/mg respectively. These values are markedly
higher than those reported by Jia et al (15).
The results also showed that the inhibitory effect
of the fusion protein on tumor growth was dose-dependent
(r2=0.993) in S180 tumor-bearing mice, and that the
percentage reduction in mean tumor volume in the high dose group
was up to 75.61% compared with the control group. uPA is the main
plasminogen activator in vivo that cleaves plasminogen to
plasmin. Plasmin hydrolyzes various components of the extracellular
matrix to promote tumor cell invasion into adjacent tissue as well
as metastasis. Furthermore, the level of uPA in serum is positively
correlated with the extent of metastatic spread and activity of
tumors (16). The current study
showed that the level of uPA in tumor-bearing mouse serum decreased
as the dose of fusion protein increased. Furthermore, the levels of
uPA in the high dose group decreased by 29.40%, and tumors were
better circumscribed and easier to remove than those in the control
group. The changes in levels of uPA further reflect the anti-tumor
effects of the fusion protein. The results demonstrated that the
fusion protein induced a greater degree of tumor cell apoptosis and
that the nuclei in these cells typically showed pyknosis and lysis.
These observations were most evident in the high-dose group, in
which the percentage of apoptotic cells was increased to 71.16%.
Furthermore, studies on apoptosis show that bcl-2 and bax are the
most important inhibitor and promotor of apoptosis in the Bcl-2
family, respectively (17).
Upregulation of bcl-2 or downregulation of bax can inhibit
apoptosis in a variety of types of tumor. Conversely,
downregulation of bcl-2 or upregulation of bax can promote
apoptosis in a number of types of tumor. The expression of bcl-2 in
tumor tissues detected by immunohistochemistry decreased as the
dose of the fusion protein increased, whilst the dose of the fusion
protein had little effect on the expression of bax.
VEGF is a type of highly specific cytokine, which
promotes angiogenesis and increases vascular permeability It is
also closely associated with tumor growth and metastasis (18). The expression of VEGF in different
types of tumor tissues, detected by immunohistochemistry, showed
that VEGF-positive areas and the dose of rhTNF were negatively
correlated. Gu et al (19)
demonstrated that TNF-α selectively inhibited tumor vascular
endothelial cells and downregulated the expression of the VEGF
receptor to inhibit angiogenesis. Whether this fusion protein may
also reduce the expression of the VEGF receptor requires further
investigation.
TNF-α is frequently investigated as a potential
therapy in cancer research, but its antitumor mechanism remains
unclear. The fusion protein produced in this study had significant
antitumor effects in S180 tumor-bearing mice. A number of
mechanisms may be involved. Firstly, the fusion protein inhibited
the expression of uPA in tumor tissue (or accelerates its
hydrolysis) reducing the capacity of the tumor to invade and
transform. This led to a reduction in tumor growth. Secondly, the
fusion protein reduced the expression of bcl-2, which promotes
tumor cell apoptosis and inhibits tumor growth. Thirdly, the fusion
protein acts on vascular endothelial cells of the tumor tissue to
inhibit the expression of VEGF, thus exhibiting anti-angiogenic
activity.
In order to explore the effects on liver and kidney
function of the tumor-bearing mice, a series of indicators were
examined, which showed that the fusion protein had no significant
effect on liver and kidney indices. The decreasing level of ALT and
increasing level of ALB suggests that the fusion protein was able
to reduce tumor-induced compensatory liver injury, and enhance the
synthetic function of the liver. In terms of kidney function, the
levels of BUN and Cr in tumor-bearing mouse serum varied little in
the low dose and middle dose groups compared with the control
group, which indicated that the fusion protein had no significant
effects on mouse kidney function at these doses. However, BUN and
Cr did increase marginally in the high dose group, indicating a
degree of reversible or irreversible kidney injury at this point.
Since the fusion protein may have a protective effect on the liver
and appear not to affect kidney function within a certain dose
range, the toxic effects of TNF may be ameliorated to a certain
extent.
In conclusion, the present study obtained a high
pick-up rate of rhTNF-α fusion protein mediated by urokinase
through plasmid construction. This protein showed a clear effect on
tumor cell growth and apoptosis in S180 tumor-bearing mice, as well
as a low toxicity. There is thus significant potential for its use
in a range of clinical applications for the treatment of
cancer.
Acknowledgments
This study was supported by the First Science and
Technology Program of Guangdong province (grant no.
2008B030301023), the Science and Technology Program of Higher
Learning Institutions in Dongguan (grant nos. 200910815264 and
2012108102016) and the Innovative Experiment Program of College
Students of Guangdong (grant no. KY1040). Furthermore, the authors
would like to thank all the teachers at the Clinical Biochemistry
Staff Room Guangdong Medical College Dongguan for their technical
assistance.
References
|
1
|
Van Ostade X, Vandenabeele P, Everaerdt B,
et al: Human TNF mutants with selective activity on the p55
receptor. Nature. 361:266–269. 1993. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Gerstein ES, Shcherbakov AI, Kaz’min AI,
et al: Urokinase and tissue plasminogen activators and their type-1
inhibitor (PAI-1) in gastric cancer. Vopr Onkol. 49:165–169.
2003.In Russian.
|
|
3
|
Morodomi Y, Yano T, Kinoh H, et al:
BioKnife, a uPA activity-dependent oncolytic Sendai virus,
eliminates pleural spread of malignant mesothelioma via
simultaneous stimulation of uPA expression. Mol Ther. 20:769–777.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Thummarati P, Wijitburaphat S, Prasopthum
A, et al: High level of urokinase plasminogen activator contributes
to cholangiocarcinoma invasion and metastasis. World J
Gastroenterol. 18:244–250. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wang S, Chen T, Chen R, et al: Emodin
loaded solid lipid nanoparticles: preparation, characterization and
anti-tumor activity studies. Int J Pharm. 430:238–246. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhang XJ, Yue J and Zhao TB: Expression
change of TNF-α in myocardium and hepatic tissue of rats with
compound stress of hyperthermia and lipopolysaccharide. Asian Pac J
Trop Med. 6:300–304. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Phalitakul S, Okada M, Hara Y, et al:
Vaspin prevents methylglyoxal-induced apoptosis in human vascular
endothelial cells by inhibiting reactive oxygen species generation.
Acta Physiol (Oxf). 209:212–219. 2013.
|
|
8
|
Han W, Wang S, Liang R, et al: Non-ionic
surfactant vesicles simultaneously enhance antitumor activity and
reduce the toxicity of cantharidin. Int J Nanomedicine.
8:2187–2196. 2013.PubMed/NCBI
|
|
9
|
Jiang Y, Jiang X, Law K, et al: Enhanced
anti-tumor effect of 9-nitro-camptothecin complexed by
hydroxypropyl β cyclodextrin and safety evaluation. Int J Pharm.
415:252–258. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhou S, Yang Y, Yang Y, et al: Combination
therapy of VEGF-trap and gemcitabine results in improved anti-tumor
efficacy in a mouse lung cancer model. PLoS One. 8:e685892013.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Weber WA, Czernin J, Phelps ME and
Herschman HR: Technology Insight: novel imaging of molecular
targets is an emerging area crucial to the development of targeted
drugs. Nat Clin Pract Oncol. 5:44–54. 2008. View Article : Google Scholar
|
|
12
|
Klener P Jr and Klener P:
Molecularly-targeted and biological anti-cancer therapy. Folia Biol
(Praha). 58:1–6. 2012.
|
|
13
|
McGuire AT and Mangroo D: Cex1p is a novel
cytoplasmic component of the Saccharomyces cerevisiae nuclear tRNA
export machinery. EMBO J. 26:288–300. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhao Q, Hou G, Huang D and Chen S:
Construction and expression of hTNF-alpha fusion protein mediated
by MMP1. Sheng WuYi Xue Gong Cheng Xue Za Zhi. 28:534–537. 2011.In
Chinese.
|
|
15
|
Jia FL and Khan A: Studies on acute
toxicity and serum level changes of tumor necrosis factor. Zhongguo
Yi Xue Ke Xue Yuan Xue Buo. 11:302–305. 1989.In Chinese.
|
|
16
|
Horn LC, Pippig S, Raptis G, et al:
Clinical relevance of urokinase type plasminogen activator and its
inhibitor type 1 (PAI-1) in squamous cell carainoma of the uterine
cervix. Aust N Z J Obstet Gynaecol. 42:383–386. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Strasser A, Cory S and Adams JM:
Deciphering the rules of programmed cell death to improve therapy
of cancer and other diseases. EMBO J. 30:3667–3683. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Sivaprasad S, Govardhan B, Harithakrishna
R, et al: Association of vascular endothelial growth factor (VEGF)
gene polymorphism and increased serum VEGF concentration with
pancreatic adenocarcinoma. Pancreatology. 13:267–272. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Gu CP and Zhang YP: Progress on anti-tumor
mechanism of tumor necrosis factor-alpha. Bulletin of Chinese
Cancer. 16:102–105. 2007.In Chinese.
|