Introduction
At present, the majority of cases of sensorineural
hearing loss remain incurable. Gene therapy is an effective
approach for the treatment of genetic disorders of the inner ear,
where defective genes may be replaced or novel genes may be
introduced into specific cell types to elicit a therapeutic
response. Such genetic therapies have received significant
attention as they provide potential for the arrest, reversal or
cure of deafness (1,2).
Viral vectors, including adenovirus (AV),
adeno-associated virus (AAV) and lentivirus (LV), present the
advantage of facilitating sustained gene expression within
transfected cochlear cells, with the potential for the vector to be
dependent upon virus-specific properties. These viral vectors have
evolved to be highly efficient gene delivery systems and have been
used in various fields of inner ear research. However, while the
use of viral vectors offers numerous experimental advantages, they
are also associated with certain limitations. Significant
challenges which remain for these viral vectors include the
induction of the immune response, toxicity of the gene transfer
vehicle and potential transformation-associated risks of cells
modified by insertional mutagenesis (3–5).
Non-viral gene delivery systems have lower
immunogenicity, are comparatively easier to scale up and exhibit
enhanced abilities in terms of vector modification and DNA
incorporation capacity (6). It has
been hypothesized that gene therapy in humans will likely utilize
non-viral vectors over viral vectors in the future (6).
Non-viral, amine-containing polymers, including
polyethylenimine (PEI), polylysine and polyamidoamine dendrimers
have previously been well investigated. These cationic polymers
spontaneously form stable complexes with DNA that are able to be
employed for gene delivery (7). To
date, PEI is one of the most successful non-viral gene carriers
reported (8), newly designed and
synthesized materials are frequently compared with it and a
plethora of PEIs with various molecular weights and structures,
have garnered attention. Among them, the 25 kDa linear PEI (L-PEI)
is a benchmark, due to its relatively high gene delivery efficiency
and availability (9). Apart from
uses in cancer therapy, PEIs have been utilized in diverse fields,
including neurological research (10,11);
however, to date, few studies regarding the use of non-viral
vectors have been conducted by otolaryngologists. To the best of
our knowledge, Tan et al (14) were the first to introduce L-PEI as
a gene vector to the field of inner ear research.
However, a factor which has not yet gained
sufficient attention in the nanomedical field is that nanoparticles
are not inherently benign and therefore influence physiological
functions at the cellular, subcellular, protein and gene levels
(15,16). Nanotoxicology describes the study
of the interactions between nanostructures and biological systems,
particularly focusing on elucidating the associations between the
physiochemical properties (including the size, shape, surface
chemistry, composition and aggregation) of nanostructures with the
induction of pathophysiological or toxic biological responses
(17). The risks associated with
individual nanomaterials may differ from their bulk material
counterparts due to their altered physiochemical properties
(17). To date, suitable
standardized in vitro tests and experimental protocols for
the evaluation of nanotoxicity have remained unavailable. Whether
PEI-based nanoparticles possess nano-ototoxicity, which may result
in disadvantages associated with cochlear disruption, has remained
to be elucidated. PEI is unable to be effectively eliminated and
therefore accumulates over time, highlighting an urgent need for
the confirmation of the safety profile of PEI prior to the
initiation of PEI-associated clinical trials (16). The present study therefore aimed to
identify any potential nano-ototoxicity that PEI nanoparticles may
exert on the cultured cochlear epithelium of neonatal mice, and
provide a rationale for the use of in vivo animal studies to
assess nanotoxicity of various nanoparticles in multiple cell
types.
Materials and methods
Nanoparticle formation
Plasmids encoding the enhanced green fluorescent
protein reporter gene (pEGFP-C1) were purchased from Clontech
Laboratories, Inc. (Mountain View, CA, USA) and 25 kDa L-PEI was
obtained from Sigma-Aldrich (St. Louis, MO, USA). The plasmids were
amplified and purified according to the manufacturer’s
instructions. Briefly, the pEGFP-C1 plasmid was propagated in
competent Escherichia coli strain DH5α (Gibco, Rockville,
MD, USA) and purified using the Qiagen Endotoxin-Free Plasmid
Purification kit (Qiagen, Valencia, CA, USA). The UV absorbance was
measured at 260 nm using a biophotometer (6131; Eppendorf, Hamburg,
Germany), in order to determine DNA concentration. The purity of
the DNA was confirmed by measuring UV absorbance at 260/280 nm.
Stock solutions of L-PEI (1 μg/μl) and plasmid DNA
(pDNA; 1 μg/μl) were prepared, and prior to each
experiment were combined at various weight ratios of up to 2
μg:14 μg (pDNA/L-PEI). The pH of the stock L-PEI
solution was neutralized using concentrated HCl (Sunshine
Biotechnology Co., Ltd., Nanjing, China). Polyplex formation
utilized solutions of equal volumes (50 μl) with L-PEI being
gently added to the plasmid solution. Samples were continuously
vortexed during addition and equilibrated at room temperature for
20 min prior to use. L-PEI was stored in aliquots at −20°C and the
polyplex was freshly prepared prior to each individual measurement.
Complexes were formed in phosphate-buffered saline (PBS; pH 7.4)
(Sangon Biotechnology Co., Ltd., Shanghai, China) unless otherwise
stated.
Characterization
The morphologies of the L-PEI/pDNA polyplexes were
examined with a JEM-200CX transmission electron microscope (JEOL,
Ltd, Tokyo, Japan). Polyplexes were prepared as described above.
The samples were prepared as follows: A drop of dilute particle
dispersion was spread over a carbon-coated grid (Agar Scientific
Ltd., Stansted, UK). The dried specimens were subsequently
negatively stained with a drop of 2% phosphotungstic acid
(Sigma-Aldrich) for 30 min and dried at room temperature.
Agarose gel retardation assay
An agarose gel retardation assay was used to
evaluate the binding ability of L-PEI/pDNA. Complexes were prepared
in PBS using 2 μg pDNA and various weights of L-PEI, ranging
from 0 to 14 μg. Following the addition of appropriate
amounts of 6X gel-loading buffer (Thermo Fisher Scientific,
Waltham, MA, USA), the samples were electrophoresed on a 1% (w/v)
agarose gel containing ethidium bromide (0.5 μg/ml of the
gel) in 0.5X Tris-boric acid-EDTA (Sunshine Biotechnology Co.,
Ltd.) at 100 V. DNA in the gel was visualized with an ultraviolet
illuminator and photographed with a UVIpro gel image system
(UVItec, Ltd, Cambridge, UK).
Dissection, culture and transfection
procedure of the cochlear explants
Four-day-old C57BL/6J mice (male:female, 1:1) were
purchased from the Model Animal Research Center of Nanjing
University (Nanjing, China). Animals were housed under
pathogen-free conditions, under a natural day/night cycle. All
animal experimental procedures were conducted in accordance with
the guidelines set by the Animal Care and Use Committee of Nanjing
Drum Tower Hospital and Nanjing University (Nanjing, China). A
total of 48 four-day-old mouse pups (weighing 1.25±0.70 g) were
anesthetized with CO2 and were sacrificed. The
experiments were performed in quadruplicate. Dissection and cell
culture were conducted as previously described (18). Briefly, following being cleaned
with 75% ethanol (Sangon Biotechnology Co., Ltd.), pups were
decapitated on ice and the cochleae were dissected. Soft tissues
and cartilage were removed, and the cochlear epithelium was
isolated. The specimens were separated into apical, middle and
basal turns. The organotypic cultures were maintained in Dulbecco’s
modified Eagle’s medium (HyClone, Thermo Fisher Scientific)
supplemented with B27 (Gibco, Invitrogen Life Technologies,
Carlsbad, CA, USA) and 10% fetal bovine serum (HyClone) at 37°C in
a humidified atmosphere containing 5% CO2. After 24 h,
the culture medium was removed and replaced with polyplex solution
and fresh media. Cochlear explants were randomly divided into two
groups: Control (without transfection; 0, 24 and 48 h) and
treatment (0, 24 and 48 h post-transfection).
Immunocytochemical analysis
Following the indicated length of incubation,
cochlear explants were fixed with 4% paraformaldehyde for 30 min.
Following rinsing with 0.1 mM PBS, the cochlear explants were
permeabilized with 0.5% Triton X-100 in PBS for 20 min and washed
three times with PBS. Incubation with PBS containing 5% horse serum
(Sigma-Aldrich) and 0.1% Triton X-100 (Thermo Fisher Scientific)
for 1 h at room temperature blocked non-specific binding. The
explants were subsequently incubated with the following primary
immunoglobulin G antibodies: Unconjugated mouse monoclonal
anti-β-tubulin III (1:200; Sigma-Aldrich) or goat polyclonal
anti-otoancorin (1:200; Santa Cruz Biotechnology, Inc., Dallas, TX,
USA) overnight at 4°C. The explants were then washed three times
with 0.1% Tween (Sigma-Aldrich) in PBS, and incubated with the
following secondary antibodies: Donkey anti-goat Cy3 (1:500;
Invitrogen Life Technologies, Carlsbad, CA, USA) or donkey
anti-mouse Alexa Flour 647 (1:500; (Invitrogen Life Technologies)
overnight at room temperature. Fluorescein
isothiocyanate-phalloidin (Sigma-Aldrich) and Hoechst 33342
(Invitrogen Life Technologies) were added to the secondary antibody
solution to label hair cell stereocilia and the nuclei. Following
staining, samples were mounted in Fluoromount G (Thermo Fisher
Scientific). All images were captured using a Nikon confocal laser
scanning microscope (TE2000-U; Nikon Corp., Tokyo, Japan), and
images were analyzed using Nis-element Basic research software
(version 4.0; Nikon Corp.). Each experiment was performed in
triplicate. Negative control experiments were performed with the
omission of the primary antibody, in order to verify the lack of
non-specific binding of the secondary antibody.
Transmission electron microscopy
(TEM)
For TEM, the cochlear explants were cultured on
polyethylene terephthalate membranes (EMD Millipore, Billerica, MA,
USA). Samples were collected at various time-points (without
transfection, and 0, 24 and 48 h post-transfection) and fixed in
2.5% glutaraldehyde (Sigma-Aldrich) in PBS at 4°C for 6 h, followed
by 1% osmium tetroxide (Sigma-Aldrich) at 4°C for 1 hour,
dehydration, infiltration and polymerization in araldite. Ultrathin
sections (80–100 nm) were post-stained with uranyl acetate
(Sigma-Aldrich) for 30 min in the dark and lead citrate
(Sigma-Aldrich) for 6 min. The sections were then examined using a
Hitachi-7650 TEM (Hitachi, Ltd, Tokyo, Japan). In all TEM
experiments, 2 μg pDNA and 14 μg L-PEI was used.
Statistical analysis
Values are presented as the mean ± standard
deviation. Statistical variations between the groups were analyzed
using the Student-Newman-Keuls-Q test with SAS version 9 software
(SAS, Cary, NC, USA). P<0.05 was considered to indicate a
statistically significant difference between values.
Results
Characterization of the PEI-pDNA
nanoparticles
The PEI-pDNA nanoparticles were almost spherical
with a diameter of ~20–100 nm (Fig.
1A). In PBS solution they frequently aggregated, forming larger
clots.
 | Figure 1Characterization of L-PEI/pDNA
polyplexes. (A) TEM analysis of polyplex morphologies (pDNA/L-PEI,
2 μg:14 μg). Polyplex particles were formed by
electrostatic interactions between L-PEI and pDNA. (B) Agarose gel
electrophoresis of 0.5 μg enhanced green fluorescent pDNA
complexed with various amounts of L-PEI. Lane 1, 0 μg; lane
2, 0.5 μg; lane 3, 1 μg; lane 4, 1.5 μg; lane
5, 2 μg; lane 6, 2.5 μg; lane 7, 3 μg; lane 8,
4 μg. L-PEI, linear polyethylenimine; pDNA, plasmid DNA. |
Agarose gel retardation is achieved with
0.5 μg L-PEI
A prerequisite for potential polymeric gene carriers
is the ability to condense DNA (19). A gel retardation assay was
therefore conducted to determine the quantity of L-PEI that was
able to condense DNA. The binding of L-PEI to pDNA results in
neutralization of the negative charges in the phosphate backbone of
DNA; these large electroneutralized complexes are therefore
incapable of migrating towards the anode through the agarose gel
(20). A series of polyplexes were
prepared using the protocol described (2 μg pEGFP-C1 plasmid
DNA complexed with 0–14 μg L-PEI) in a reduced total volume
of 50 μl. In PBS, retardation of migration was achieved with
0.5 μg L-PEI (Fig. 1B).
In vitro transfection studies of neonatal
mouse cochlear cultures
The sensory epithelium of four-day-old mouse pups
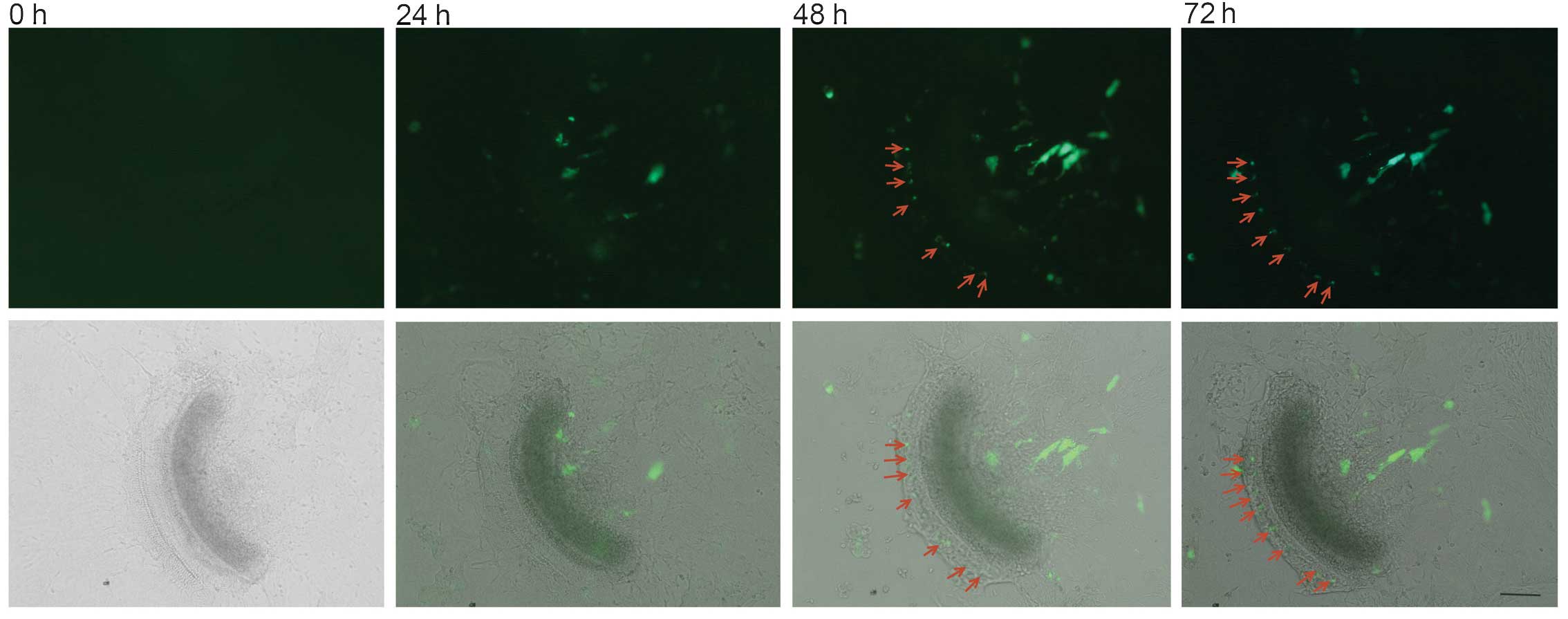
was transfected at various L-PEI to eGFP plasmid weight ratios in
order to identify the optimum transfection ratio. A complex
comprised of 3 μg L-PEI and 1 μg pDNA was
demonstrated to be able to transfect the outgrowing cells, and it
was revealed that the use of >2 μg pDNA with ≥5 μg
L-PEI transfected the fibrocytes in the spiral limbus (data not
shown). Of note, when the L-PEI/pDNA ratio was altered to 7:1
μg, the cells in the organ of the Corti region were
transfected (Fig. 2).
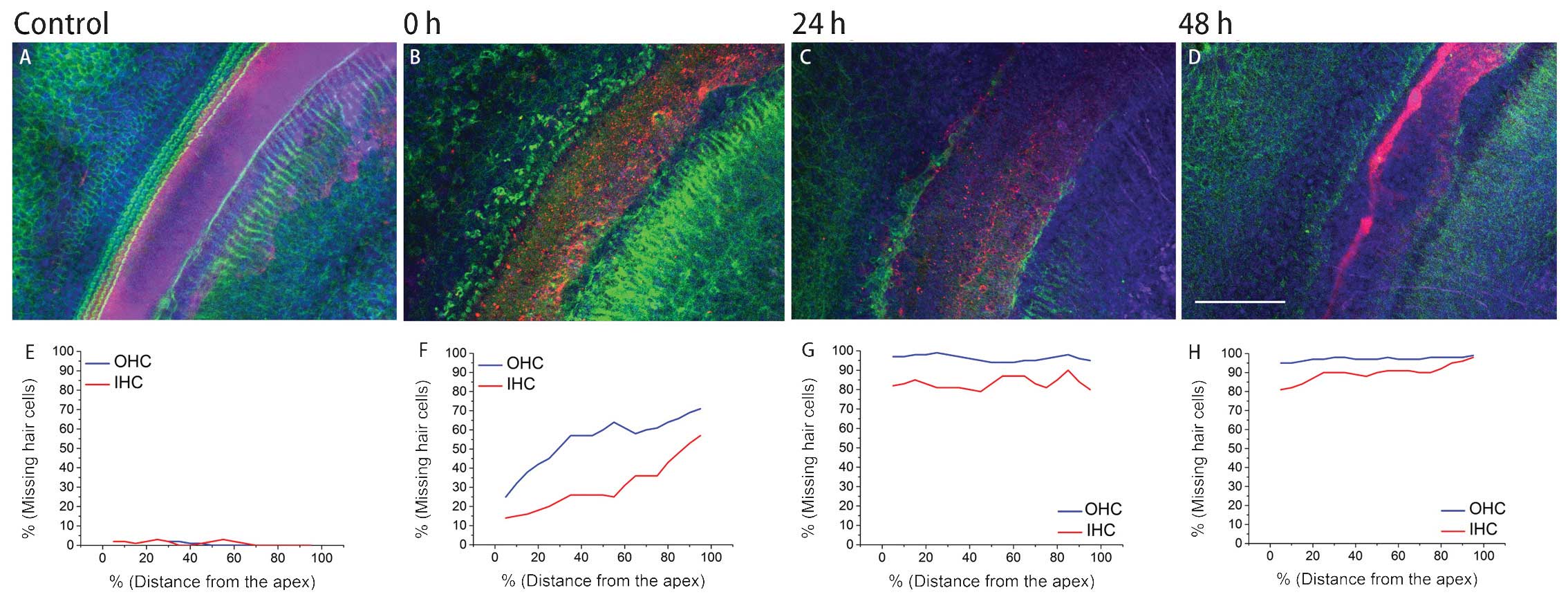
Hair cell damage occurs immediately
following transfection
No marked hair cell loss was identified in the
control samples. The explants survived well in the nutritional
culturing medium, even following 72 h of culture. As the above
level of transfection was only achieved when the L-PEI/pDNA weight
ratio was increased to 7:1, the effects of nanoparticles generated
at this transfection level on the hair and inter-dental cells in
the spiral limbus was further examined. The otoancorin antibody was
applied not only for staining of the interdental cells, but also
for better discrimination between the spiral limbus region and the
hair cell region under harsh nano-ototoxic conditions. Following
addition of the polyplex, oto-nanotoxicity to hair cells
immediately commenced (Fig. 3A).
Compared with the inner hair cells (IHCs), the outer hair cells
(OHCs) were more vulnerable at the acute phase. Following 24 h of
culture with the polyplex, only a sparse population of hair cells
remained (Fig. 3B). Following 48 h
of culture, few hair cells survived and the damage had spread to
the Hensen and Claudius cells’ region (Fig. 3C). Statistically significant
differences in the levels of cellular loss between each time-group
were detected (all P<0.05) for IHCs and OHCs (Fig. 3E-H). Counting random regions of
interdental cells revealed that statistical divergence also existed
(all P<0.05).
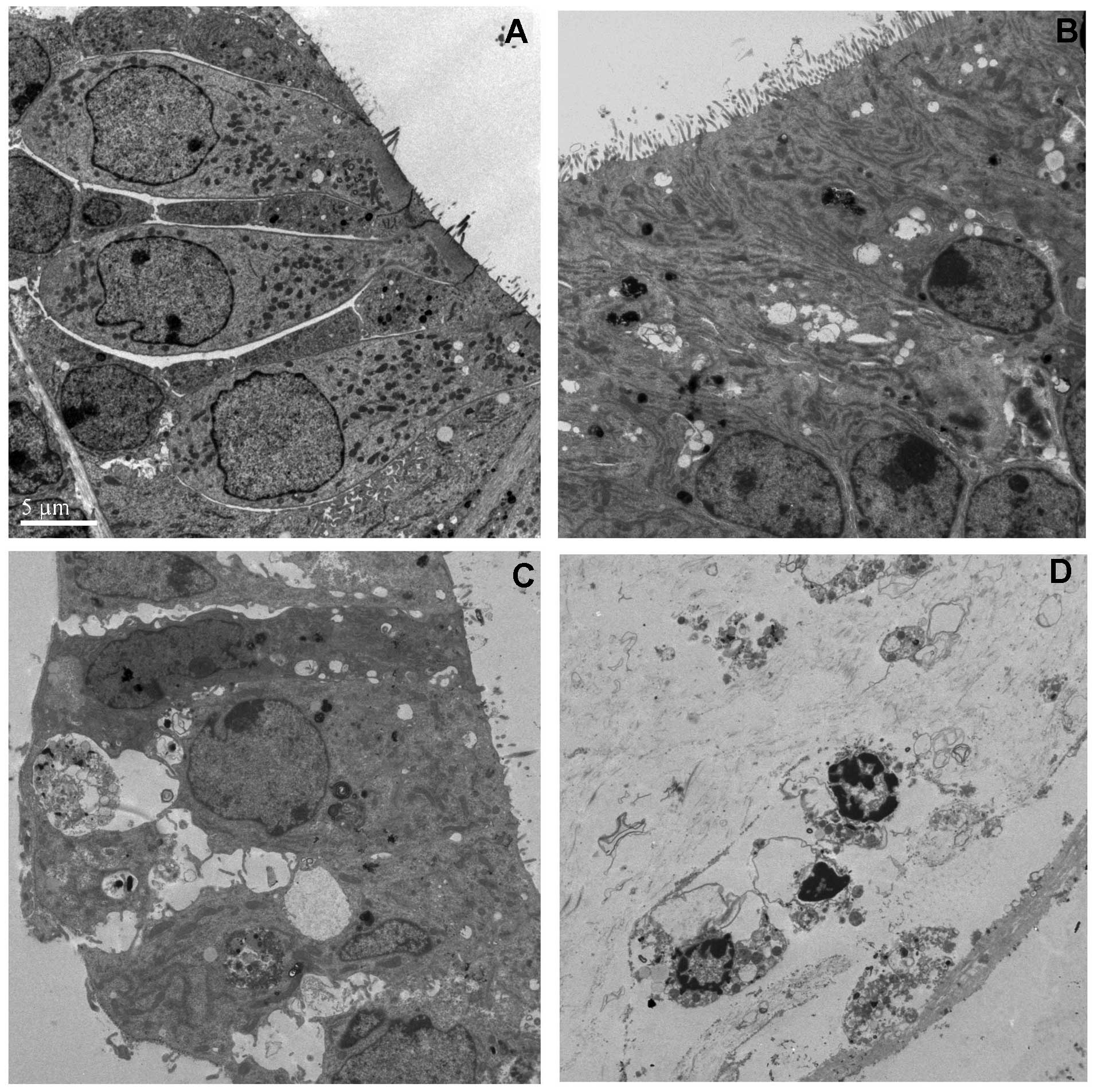
Cellular toxicity occurs immediately
following transfection
The toxic effects of the L-PEI-pDNA polyplexes were
further evaluated by TEM. The hair cells and supporting cells were
functioning effectively, even on the fourth day of explantation.
The cell membranes were intact, stereocillia were orderly arranged
and the organelles appeared normal (Fig. 4A). However, following incubation
with the polyplex, it was demonstrated that the nanoparticles were
able to penetrate into the cell bodies of the hair cells and their
supporting cells (Fig. 4B–D)
(21–23). Immediately following transfection,
large quantities of bubbles were detected within the sensory
epithelial cells, stereocilla were partially damaged and
mitochondrial numbers were significantly reduced. Approximately 24
h later, the sensory epithelium detached from the basilar membrane,
the cell membrane was ruptured and large quantities of cytoplasm
were lost from the cell bodies. Within certain sections, condensed
chromatin in the nucleus and apoptotic bodies were identified.
Following a further 24 h of incubation, the cells died. This
phenomenon was in accordance with the results of the
immunofluorescent staining analysis.
Discussion
Non-viral gene delivery has emerged as a potential
alternative to conventional viral transfection systems. The
structure and physical properties of synthetic gene carriers,
including their size, shape, side groups and charge, influence
their association with nucleic acids, intracellular pathways and
overall delivery efficiency (24,25).
Since the first published examination of PEI as a gene delivery
vehicle (26), numerous studies
aimed at elucidating its (or modified PEI’s) potential role as a
nanocarrier for DNA, RNA and oligonucleotides (19). The development of PEI represented
significant progress in the field of non-viral gene transfer
(27), and to date, PEI remains
one of the most effective and frequently utilized non-viral vectors
(28).
PEI is a highly positively charged, synthetic
polymer, which is comprised of primary, secondary and tertiary
amino groups at a 1:2:1 ratio (20). Every third atom of PEI is a
nitrogen that is able to be protonated, and ~20% of these nitrogens
are protonated under normal physiological conditions (29). PEI is therefore able to alter its
ionization state over a broad pH range, conferring significant
buffering capacity (20). Via
electrostatic interaction with negatively charged DNA, these agents
typically form toroidal or spherical particles. Fourier transform
infrared analysis revealed that DNA is maintained in B form
following complexation with various amounts, molecular weights or
forms of PEI (30).
The positively charged PEI polyplexes are able to
interact with highly anionic polysaccharides, for example
glycosaminoglycans, including hyaluronic acid, chondroitin sulfate
and heparan sulfate, that are located in the extracellular space
and on the surface of the majority of cells (31–33).
Subsequently, the polyplexes are internalized via endocytosis or
pinocytosis (34–35). PEI therefore efficiently mediates
gene delivery without the requirement for exogenous endosomolytic
agents. According to the proton sponge hypothesis, during the
transition from the plasma membrane (pH 7) to the early endosome
(pH 6.5), late endosome (pH 5.5) and finally to the lysosome (pH
5), ionizable PEI exhibits broad pKa values and functions as a
buffer by pumping additional protons along with the concurrent
influx of chloride ions, which increases the ionic strength inside
the endolysosome (29,36). This results in an influx of water,
osmotic swelling and endolysosomal bursting, resulting in the
escape of the polyplex in order to protect the DNA in the cell
(29,36). However, Godbey et al
(27) demonstrated that the
transfection efficiency of PEI is not a result of its lysosomal
buffering; instead, PEI moves towards the nucleus while keeping the
complexed plasmids tightly condensed and protected.
Protein, microtubules and other organelles are all
able to perturb polyplex movement. Naked DNA moves slowly through
the cytoplasm (5). It has been
suggested that positively charged polyplexes are actively
transported by microtubules and microfilaments (37–40).
Polyplexes may move along microtubules via non-specific
interactions with anionic microtubules or motor proteins, or they
may rely upon the natural transport of endolysosomes along the
microtubules. Polyplexes may be delivered to the perinuclear region
via the microtuble and actin networks (41–43).
Nuclear entry is used to transport objects <40 nm in diameter
(44), and certain polyplexes or
free plasmids may enter the nucleus via this mechanism, while
numerous polyplexes or plasmids are localized to the perinuclear
region where they await rupture of the nuclear membrane envelope
during cell division (45). PEI
also possesses an additional mechanism for mediating the nuclear
entry of pDNA which does not require cell division, as post-mitotic
cells, including neurons, are able to be transfected in
vitro and in vivo. However, the intracellular barriers
to plasmid trafficking vary quantitatively with cell type (41,46,47).
Originally, 0.5% low-molecular weight PEI (molecular
weight, 1,800) in cacodylate buffer was used as a cationic tracer
for the identification of anionic sites created by chondroitin- and
heparin-rich glycosaminoglycan on the capillary and subepithlial
basement membrane of the stria vascularis, spiral ligament, spiral
limbus, Reissner’s membrane and the vestibular sensory epithlium
(ampulla and macula) (48,49). These anionic sites were considered
to be associated with a negative charge
blood-labyrinth/perilymphatic-endolymphatic barrier for
electrically charged macromolecules and ions, and with the
maintenance of steep chemical gradients (50). This barrier already exists in
four-day-old rats and remains immature in the early postnatal
period. Once damaged, the biological membrane barrier becomes more
permeable, resulting in hearing impairment (51). Though low-molecular weight PEIs are
less toxic (52), they lack
effective transfection properties as, due to their low positive
charge, they are incapable of effectively condensing DNA (53). Additionally, their low associated
surface charge does not induce effective cellular uptake via
charge-mediated interactions (53).
The forms of PEI most widely used for cellular
transfection are 25 kDa L-PEI, branched PEI (B-PEI) and 22 kDa
L-PEI; however, their transfection abilities differ. The greater
transfection efficiency of L-PEI may be a result of its inherent
kinetic instability under salt conditions (54). The 22-kDa L-PEI has been
demonstrated to have higher transfection activity in vivo
when complexed with plasmids in salt-free buffer and administered
intravenously (55–57). Under equivalent salt conditions,
the transfection efficiency of 22-kDa L-PEI is higher than that of
25-kDa B-PEI. Tan et al (14) used 22-kDa L-PEI (ExGen500) diluted
in 5% (w/v) glucose solution for the transfection of cochlear cells
in vitro and in vivo. Using the relatively low
nitrogen:phosphorus ratios (26)
(the ratio of moles in the amine group of cationic polymers to
those of the phosphate groups), neither syringe transfection nor
weeks of infusion via the osmotic pump yielded satisfactory levels
of transfection in vivo (14). In the present study, PBS was
selected as the dilution medium as the number of PEI-based
nanoparticles generated in salt-free conditions were reported to
inefficiently transfect the cultured cell lines. Furthermore, PBS
more closely mimicked the high ionic strength of the cochlear
lymphatic system. In accordance with the results of previous
experiments, which demonstrated that supporting cells were able to
be transfected by L-PEI-pDNA nanoparticles, the results of the
present study also indicated that the cells of the sensory
epithelium may be transfected. Furthermore, as the hair cells
barely survived following transfection, eGFP protein was likely
expressed by the supporting cells.
Certain concerns have emerged regarding the toxicity
of nanoparticle delivery systems (58). In vitro studies have
demonstrated that the cytotoxicity of PEI may potentially be a
result of the presence of excess free polymer following
complexation with DNA, and its limited biodegradability (59–61).
This lack of biodegradability may be attributed to the highly
branched non-degradable methylene backbone, and the high cationic
charge density of PEI, which is inherently toxic to normal tissues
if the carrier is not degraded following internalization (62,63).
The branched form of PEI exhibits a higher condensation ability and
transfection efficiency, but is associated with higher toxicity
(54). Furthermore, the
endocytosed nanoparticles may induce oxidative stress by
stimulating the production of intracellular reactive oxygen
species, a process which represents the first event in a cellular
toxicity cascade, where multiple gene expression alterations are
elicited in cells in vitro and in vivo (64). Moghimi et al (65) revealed that 25 kDa B-PEI induced
membrane damage and initiated apoptosis in three clinically
relevant human cell lines. As early as 30 min following
transfection, compromised membrane integrity resulted in
necrotic-like changes. Significant lactate dehydrogenase was
released and phosphatidylserine was translocated from the inner
plasma membrane to the cell surface. At 24 h post-transfection,
PEI-induced channel formation in the outer mitochondrial membrane
triggers a ‘mitochondrially mediated apoptotic program’,
pro-apoptotic cytochrome c is released and the apopotic
pathway is activated (65–67). Lin et al (68) additionally demonstrated the
involvement of autophagy. The results of the present study
supported this hypothesis, and further indicated that the
nanoparticles based on 25 kDa L-PEI resulted in similar
oto-nanotoxicity effects in the cultured cochlear epithelium. The
auditory sensory epithelial cells (in particular, the outer and
inner hair cells) were degenerated in all experimental groups, more
markedly so in the 48 h group, in addition to other cell types, for
example interdental cells in the spiral limbus. However, the hair
cells appeared to be more sensitive to oto-nanotoxicity than
interdental cells. These differences in the extent of
oto-nanotoxicity were suggested to be associated with the inherent
variations in the uptake and removal ability of various cell types,
which facilitate their ability to deal with nanomaterials,
including antioxidant levels or repair mechanisms (69). Using zebrafish as a model animal
for nanocytotoxic research (70),
Rizzo et al (71) verified
that the 25-kDa L-PEI prevented proper embryo development at
concentrations as low as 0.01 μg/μl. Such potentially
hazardous effects, which are intrinsic to multiple synthetic
polymer vector carriers, including PEI and its derivatives,
represent a significant limitation to their use. The present study
demonstrated that the use of PEI-derived gene delivery vehicles in
the inner ear remains limited, due to their relatively high
cytotoxicity.
Recent developments in nanomedicine have revealed
novel insights into inner ear gene therapy (72). However, current knowledge regarding
nanotoxicity is dwarfed by what remains to be elucidated (69). Fears over the potential dangers of
nanomedicine may be exaggerated, but are not necessarily unfounded.
The results of the present study revealed that the cultured hearing
sensory epithelium was able to be transfected by the classical
non-viral cationic polymer PEI-based nanoparticles. However, the
excess polyplexes and cationic applications of PEI hampered its
effectiveness by inducing nano-ototoxicity to the delicate cochlear
structures. The molecular mechanisms underlying the interactions
between nanoparticles and cells, and a more detailed nanotoxicty
profile require further elucidation. For these reasons, significant
obstacles remain regarding the use of nanoparticle therapeutics,
prior to their evaluation in clinical trials and subsequent use for
the delivery of therapeutics in humans. Novel high-throughput
approaches to design and screening may aid in the assessment of
chemically and physically diverse nanomaterials, enabling the
identification of a successful, highly efficient, biocompatible,
biodegradable, non-toxic next-generation polymer vectors for use in
inner ear gene therapy (28,73).
Acknowledgments
This study was supported by the Jiangsu Provincial
Natural Science Foundation of China (no. BK2010104).
References
|
1
|
Lustig LR and Akil O: Cochlear gene
therapy. Curr Opin Neurol. 25:57–60. 2012. View Article : Google Scholar
|
|
2
|
Ryan AF, Mullen LM and Doherty JK:
Cellular targeting for cochlear gene therapy. Adv Otorhinolaryngol.
66:99–115. 2009.PubMed/NCBI
|
|
3
|
Marshall E: Gene therapy what to do when
clear success comes with an unclear risk? Science. 298:510–511.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sadelain M: Insertional oncogenesis in
gene therapy: how much of a risk? Gene Ther. 11:569–573. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Pack DW, Hoffman AS, Pun S and Stayton PS:
Design and development of polymers for gene delivery. Nat Rev Drug
Discov. 4:581–593. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wang W, Li W, Ma N and Steinhoff G:
Non-viral gene delivery methods. Curr Pharm Biotechnol. 14:46–60.
2013.PubMed/NCBI
|
|
7
|
Zhou T, Llizo A, Wang C, Xu G and Yang Y:
Nanostructure-induced DNA condensation. Nanoscale. 5:8288–8306.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Godbey WT, Wu KK and Mikos AG:
Poly(ethylenimine) and its role in gene delivery. J Control
Release. 60:149–160. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wanga P, Zhang P, Huang J, Li M and Chen
X: Trichostatin a protects against cisplatin-induced ototoxicity by
regulating expression of genes related to apoptosis and synaptic
function. Neurotoxicology. 37:51–62. 2013. View Article : Google Scholar
|
|
10
|
Bergen JM, Park IK, Horner PJ and Pun SH:
Nonviral approaches for neuronal delivery of nucleic acids. Pharm
Res. 25:983–998. 2008. View Article : Google Scholar :
|
|
11
|
Pérez-Martínez FC, Carrión B and Ceña V:
The use of nanoparticles for gene therapy in the nervous system. J
Alzheimers Dis. 31:697–710. 2012.PubMed/NCBI
|
|
12
|
Kim DK, Park SN, Park KH, et al:
Development of a drug delivery system for the inner ear using
poly(amino acid)-based nanoparticles. Drug Deliv. Jan 22–2014.Epub
ahead of print. View Article : Google Scholar
|
|
13
|
Pritz CO, Dudás J, Rask-Andersen H,
Schrott-Fischer A and Glueckert R: Nanomedicine strategies for drug
delivery to the ear. Nanomedicine (Lond). 8:1155–1172. 2013.
View Article : Google Scholar
|
|
14
|
Tan BT, Foong KH, Lee MM and Ruan R:
Polyethylenimine-mediated cochlear gene transfer in guinea pigs.
Arch Otolaryngol Head Neck Surg. 134:884–891. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Nel A, Xia T, Mädler L and Li N: Toxic
potential of materials at the nano level. Science. 311:622–627.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Calarco A, Bosetti M, Margarucci S, et al:
The genotoxicity of PEI-based nanoparticles is reduced by
acetylation of polyethylenimine amines in human primary cells.
Toxicol Lett. 218:10–17. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
El-Ansary A and Al-Daihan S: On the
toxicity of therapeutically used nanoparticles: an overview. J
Toxicol. 2009:7548102009. View Article : Google Scholar
|
|
18
|
Sobkowicz HM, Loftus JM and Slapnick SM:
Tissue culture of the organ of Corti. Acta Otolaryngol Suppl.
502:3–36. 1993.PubMed/NCBI
|
|
19
|
Kichler A: Gene transfer with modified
polyethylenimines. J Gene Med. 1:S3–S10. 2004. View Article : Google Scholar
|
|
20
|
Akinc A, Thomas M, Klibanov AM and Langer
R: Exploring polyethylenimine-mediated DNA transfection and the
proton sponge hypothesis. J Gene Med. 7:657–663. 2005. View Article : Google Scholar
|
|
21
|
Zhang W, Zhang Y, Löbler M, et al: Nuclear
entry of hyperbranched polylysine nanoparticles into cochlear
cells. Int J Nanomedicine. 6:535–546. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zou J, Saulnier P, Perrier T, et al:
Distribution of lipid nanocapsules in different cochlear cell
populations after round window membrane permeation. J Biomed Mater
Res B Appl Biomater. 87:10–18. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Roy S, Johnston AH, Newman TA, et al:
Cell-specific targeting in the mouse inner ear using nanoparticles
conjugated with a neurotrophin-derived peptide ligand: potential
tool for drug delivery. Int J Pharm. 390:214–224. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Tang MX and Szoka FC: The influence of
polymer structure on the interactions of cationic polymers with DNA
and morphology of the resulting complexes. Gene Ther. 4:823–832.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Shim MS and Kwon YJ: Controlled
cytoplasmic and nuclear localization of plasmid DNA and siRNA by
differentially tailored polyethylenimine. J Control Release.
133:206–213. 2009. View Article : Google Scholar
|
|
26
|
Boussif O, Lezoualch F, Zanta MA, et al: A
versatile vector for gene and oligonucleotide transfer into cells
in culture and-in vivo polyethylenimine. Proc Natl Acad Sci USA.
92:7297–7301. 1995. View Article : Google Scholar
|
|
27
|
Godbey WT, Barry MA, Saggau P, Wu KK and
Mikos AG: Poly(ethylenimine)-mediated transfection: a new paradigm
for gene delivery. J Biomed Mater Res. 51:321–328. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Patnaik S and Gupta KC: Novel
polyethylenimine-derived nanoparticles for in vivo gene delivery.
Expert Opin Drug Deliv. 10:215–228. 2013. View Article : Google Scholar
|
|
29
|
Behr JP: The proton sponge: a trick to
enter cells the viruses did not exploit. Chimia International
Journal for Chemistry. 51:34–36. 1997.
|
|
30
|
Choosakoonkriang S, Lobo BA, Koe GS, Koe
JG and Middaugh CR: Biophysical characterization of PEI/DNA
complexes. J Pharm Sci. 92:1710–1722. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ruponen M, Honkakoski P, Rönkkö S,
Pelkonen J, Tammi M and Urtti A: Extracellular and intracellular
barriers in non-viral gene delivery. J Control Release. 93:213–217.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wiethoff CM, Smith JG, Koe GS and Middaugh
CR: The potential role of proteoglycans in cationic lipid-mediated
gene delivery. Studies of the interaction of cationic lipid-DNA
complexes with model glycosaminoglycans. J Biol Chem.
276:32806–32813. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Artzner F, Zantl R and Rädler JO:
Lipid-DNA and lipid-polyelectrolyte mesophases: structure and
exchange kinetics. Cell Mol Biol (Noisy-le-grand). 46:967–978.
2000.
|
|
34
|
Mislick KA and Baldeschwieler JD: Evidence
for the role of proteoglycans in cation-mediated gene transfer.
Proc Natl Acad Sci USA. 93:12349–12354. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wiethoff CM and Middaugh CR: Barriers to
nonviral gene delivery. J Pharm Sci. 92:203–217. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Payne CK: Imaging gene delivery with
fluorescence microscopy. Nanomedicine (Lond). 2:847–860. 2007.
View Article : Google Scholar
|
|
37
|
De Smedt SC, Demeester J and Hennink WE:
Cationic polymer based gene delivery systems. Pharm Res.
17:113–126. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Pollard H, Remy JS, Loussouarn G,
Demolombe S, Behr JP and Escande D: Polyethylenimine but not
cationic lipids promotes transgene delivery to the nucleus in
mammalian cells. J Biol Chem. 273:7507–7511. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Wang T, Upponi JR and Torchilin VP: Design
of multifunctional non-viral gene vectors to overcome physiological
barriers: dilemmas and strategies. Int J Pharm. 427:3–20. 2012.
View Article : Google Scholar
|
|
40
|
Grosse S, Aron Y, Thévenot G, Monsigny M
and Fajac I: Cytoskeletal involvement in the cellular trafficking
of plasmid/PEI derivative complexes. J Control Release.
122:111–117. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Suh J, Wirtz D and Hanes J: Efficient
active transport of gene nanocarriers to the cell nucleus. Proc
Natl Acad Sci USA. 100:3878–3882. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Bausinger R, von Gersdorff K, Braeckmans
K, et al: The transport of nanosized gene carriers unraveled by
live-cell imaging. Angew Chem Int Ed Engl. 45:1568–1572. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Kulkarni RP, Wu DD, Davis ME and Fraser
SE: Quantitating intracellular transport of polyplexes by
spatio-temporal image correlation spectroscopy. Proc Natl Acad Sci
USA. 102:7523–7528. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Pante N and Kann M: Nuclear pore complex
is able to transport macromolecules with diameters of about 39 nm.
Mol Biol Cell. 13:425–434. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Grosse S, Aron Y, Thévenot G, François D,
Monsigny M and Fajac I: Potocytosis and cellular exit of complexes
as cellular pathways for gene delivery by polycations. J Gene Med.
7:1275–1286. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Godbey WT, Wu KK and Mikos AG: Tracking
the intracellular path of poly(ethylenimine)/DNA complexes for gene
delivery. Proc Natl Acad Sci USA. 96:5177–5181. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Abdallah B, Hassan A, Benoist C, Goula D,
Behr JP and Demeneix BA: A powerful nonviral vector for in vivo
gene transfer into the adult mammalian brain: polyethylenimine. Hum
Gene Ther. 7:1947–1954. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Suzuki M, Kitamura K and Nomura Y: Anionic
sites of the basement membrane of the labyrinth. Acta Otolaryngol
Suppl. 481:112–115. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Suzuki M and Kaga K: Effect of cisplatin
on the negative charge barrier in strial vessels of the guinea
pigs. A transmission electron microscopic study using
polyethyleneimine molecules. Eur Arch Otorhinolaryngol.
253:351–355. 1996. View Article : Google Scholar
|
|
50
|
Yamasoba T, Suzuki M and Kaga K: Influence
of chronic kanamycin administration on basement membrane anionic
sites in the labyrinth. Hear Res. 102:116–124. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Suzuki M, Yamasoba T and Kaga K:
Development of the blood-labyrinth barrier in the rat. Hear Res.
116:107–112. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Breunig M, Lungwitz U, Liebl R, et al:
Gene delivery with low molecular weight linear polyethylenimines. J
Gene Med. 7:1287–1298. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Godbey WT, Wu KK and Mikos AG: Size
matters: molecular weight affects the efficiency of
poly(ethylenimine) as a gene delivery vehicle. J Biomed Mater Res.
45:268–275. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Wightman L, Kircheis R, Rössler V, et al:
Different behavior of branched and linear polyethylenimine for gene
delivery in vitro and in vivo. J Gene Med. 3:362–372. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Goula D, Remy JS, Erbacher P, et al: Size,
diffusibility and transfection performance of linear PEI/DNA
complexes in the mouse central nervous system. Gene Ther.
5:712–717. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Goula D, Benoist C, Mantero S, Merlo G,
Levi G and Demeneix BA: Polyethylenimine-based intravenous delivery
of transgenes to mouse lung. Gene Ther. 5:1291–1295. 1998.
View Article : Google Scholar
|
|
57
|
Bragonzi A, Dina G, Villa A, et al:
Biodistribution and transgene expression with nonviral cationic
vector/DNA complexes in the lungs. Gene Ther. 7:1753–1760. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Maynard AD, Aitken RJ, Butz T, et al: Safe
handling of nanotechnology. Nature. 444:267–269. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Boeckle S, von Gersdorff K, van der Piepen
S, Culmsee C, Wagner E and Ogris M: Purification of
polyethylenimine polyplexes highlights the role of free polycations
in gene transfer. J Gene Med. 6:1102–1111. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Thomas M, Ge Q, Lu JJ, Chen J and Klibanov
AM: Cross-linked small polyethylenimines: while still nontoxic,
deliver DNA efficiently to mammalian cells in vitro and in vivo.
Pharm Res. 22:373–380. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Forrest ML, Koerber JT and Pack DW: A
degradable polyethylenimine derivative with low toxicity for highly
efficient gene delivery. Bioconjug Chem. 14:934–940. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Xiong MP, Forrest ML, Ton G, Zhao A,
Davies NM and Kwon GS: Poly(aspartate-g-PEI800), a polyethylenimine
analogue of low toxicity and high transfection efficiency for gene
delivery. Biomaterials. 28:4889–4900. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Fischer D, Li Y, Ahlemeyer B, Krieglstein
J and Kissel T: In vitro cytotoxicity testing of polycations:
influence of polymer structure on cell viability and hemolysis.
Biomaterials. 24:1121–1131. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Kabanov AV: Polymer genomics: an insight
into pharmacology and toxicology of nanomedicines. Adv Drug Deliv
Rev. 58:1597–1621. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Moghimi SM, Symonds P, Murray JC, Hunter
AC, Debska g and Szewczyk A: A two-stage
poly(ethylenimine)-mediated cytotoxicity: implications for gene
transfer/therapy. Mol Ther. 11:990–995. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Florea BI, Meaney C, Junginger HE and
Borchard G: Transfection efficiency and toxicity of
polyethylenimine in differentiated Calu-3 and nondifferentiated
COS-1 cell cultures. AAPS PharmSci. 4:E122002. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Klemm AR, Young D and Lloyd JB: Effects of
polyethyleneimine on endocytosis and lysosome stability. Biochem
Pharmacol. 56:41–46. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Lin CW, Jan MS, Kuo JH, Hsu LJ and Lin YS:
Protective role of autophagy in branched polyethylenimine (25K)-
and poly(L-lysine) (30–70K)-induced cell death. Eur J Pharm Sci.
47:865–874. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Park MV, Lankveld DP, van Loveren H and de
Jong WH: The status of in vitro toxicity studies in the risk
assessment of nanomaterials. Nanomedicine (Lond). 4:669–685. 2009.
View Article : Google Scholar
|
|
70
|
Fako VE and Furgeson DY: Zebrafish as a
correlative and predictive model for assessing biomaterial
nanotoxicity. Adv Drug Deliv Rev. 61:478–486. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Rizzo LY, Golombek SK, Mertens ME, et al:
In vivo nanotoxicity testing using the zebrafish embryo assay. J
Mater Chem B Mater Biol Med. 1:3918–3925. 2013. View Article : Google Scholar
|
|
72
|
Caruthers SD, Wickline SA and Lanza GM:
Nanotechnological applications in medicine. Curr Opin Biotechnol.
18:26–30. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Lai DY: Toward toxicity testing of
nanomaterials in the 21st century: a paradigm for moving forward.
Wiley Interdiscip Rev Nanomed Nanobiotechnol. 4:1–15. 2012.
View Article : Google Scholar
|