Introduction
Renal interstitial fibrosis (RIF) is considered to
be the outcome of all types of chronic kidney disease (CKD) and the
common pathway that leads to end-stage renal failure (1). It has been demonstrated that in the
process of RIF, renal fibroblasts, the effector cells for collagen
secretion, underwent excessive proliferation and phenotypic
transformation into myofibroblasts (2,3),
resulting in abnormal proliferation of fibroblasts and accumulation
of extracellular matrix (ECM) (4–6).
It is widely accepted that the loss of peritubular
capillaries leads to local hypoxic ischemia during RIF. Adenosine
(ADO) is a key factor in hypoxic ischemia-induced signal
transduction via binding to the ADO receptor (AR) on the cell
surface (7,8). A previous study in the mouse
unilateral ureteral obstruction (UUO) model demonstrated that in
addition to occlusion, the kidney exhibited hypoxic aggravation,
ADO elevation and interstitial collagen accumulation, finally
resulting in RIF (9). It was
hypothesized that the binding of ADO with AR under hypoxia
triggered a series of pathological alterations in fibroblasts,
further inducing the progression of RIF. In the present study, an
in vitro mouse fibroblast model was used to investigate the
effects and related mechanism of the ADO signaling pathway in RIF
development.
Materials and methods
Cell culture
Cells from the mouse renal fibroblast cell line
NIH3T3 (third/fourth passage) (Promab Biotechnologies, Inc.,
Changsha, China) were divided into the following four groups: i)
Control; ii) 5′-N-ethylcarboxamidoadenosine, NECA (cat. no. 1691,
Tocris Bioscience, Minneapolis, MN, USA); iii) PT, NECA +
8-phenyltheophylline (PT) (cat. no. 2795, Tocris Bioscience); and
iv) MRS, NECA + N-(4-cyanopheny
l)-2-[4-(2,3,6,7-tetrahydro-2,6-dioxo-1,3-dipropyl-1H-purin-8-yl)phenoxy]-acetamide
(MRS1754) (cat. no. 2752, Tocris Bioscience). NECA was used as an
analogue of ADO with in vitro instability. The cells were
detected following incubation for 12, 24, 48 and 72 h. The drug
concentrations were as follows: NECA, analogue of ADO, 20 μM; 8-PT,
AR blocker, 20 μM, and MRS1754, A2BR antagonist, 20
μM.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
RT-qPCR primers were designed by Premier, version
3.0 software (Premier Biosoft International, Palo Alto, CA, USA)
and synthesized by ProMab Biotechnologies, Inc. The primer
sequences are listed in Table
I.
 | Table IPrimer sequences. |
Table I
Primer sequences.
| Gene | Primer sequence (5′
to 3′) | Amplified fragment
length (bp) | Annealing temperature
(°C) |
|---|
| α1 (I)
procollagen-F |
GTTCTCCTGGCAAAGACGGA | 199 | 58 |
| α1 (I)
procollagen-R |
CGGCCACCATCTTGAGACTT | | |
| TGF-β1-F |
AGGGCTACCATGCCAACTTC | 168 | 58 |
| TGF-β1-R |
CCACGTAGTAGACGATGGGC | | |
| α-SMA-F |
GGACTCTGGAGATGGTGTGAC | 167 | 58 |
| α-SMA-R |
CAATCTCACGCTCGGCAGTA | | |
| GAPDH
(mouse)-F |
AACTTTGGCATTGTGGAAGG | 132 | 58/59 |
| GAPDH
(mouse)-R |
GGATGCAGGGATGATGTTCT | | |
Fibroblast total RNA was isolated by the single-step
method using TRIzol (15596-026; Invitrogen Life Technologies,
Carlsbad, CA, USA). RiboLock™ Ribonuclease Inhibitor (EO0381;
Thermo Fisher Scientific, Pittsburgh, PA, USA) was used to remove
genomic DNA. The reverse transcription reaction was performed using
the RevertAid H Minus First Strand cDNA Synthesis kit (K1631;
Thermo Fisher Scientific). SYBR® Green master mix
(4309155; Applied Biosystems, Life Technologies, Foster City, CA,
USA) was used for the RT-qPCR assay of target genes.
Fibroblast cell surface AR types were identified
under hypoxia (1% O2, 5% CO2 and 94%
N2) by TaqMan probe-based analysis using JumpStart Taq
Ready Mix kit (P2893; Sigma-Aldrich, St. Louis, MO, USA). Primer
and probe sequences of A1R, A2AR,
A2BR, A3R and β-actin mRNA are shown in
Table II (10–12).
| Table IIAR primer and probe sequences. |
Table II
AR primer and probe sequences.
| Gene | Primer sequence (5′
to 3′) | Annealing
temperature (°C) |
|---|
| A1R |
F-GTTTGGCTGGAACAACCTGA
R-ACACTTGATCACGGGCTCC
Probe: FAM-AACAAGCCTGGATAGCCAACGGCA | 57 |
|
A2AR |
F-CCCCTTCATCTACGCCTACAG
R-GTGGGTTCGGATGATCTTCC
Probe: FAM-TCCGGGAGTTCCGCCAGACCT | 56 |
|
A2BR |
F-GCGAGAGGGATCATTGCTG
R-CAGGAACGGAGTCAATCCAA
Probe: FAM-CCTCTGGGTCCTTGCCTTTGGC | 56 |
| A3R |
F-ATACCAGATGTCGCAATGTGC
R-GCAGGCGTAGACAATAGGGTT
Probe: FAM-CATGGAGTTCGCGTGGGACAACAG | 56 |
| β-actin |
F-GCTCTGGCTCCTAGCACCAT
R-CCACCGATCCACACAGAGTAC
Probe: FAM-ATCAAGATCATTGCTCCTCCTGAGCGC | 60 |
The PCR cycling conditions were as follows: 40
cycles of 94°C (20 sec), 58°C (20 sec) and 72°C (30 sec), using an
ABI Prism® 7900HT sequence detection system (Applied
Biosystems Life Technologies). PCR quantification was conducted as
previously described (13).
Cell proliferation assay
The
3-(4,5-dimethylthi-azol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
assay (MTT Cell Proliferation and Cytotoxicity Detection kit;
Keygen Biotech Co., Ltd., Nanjing, China) was performed to
determine the cell growth curve. Cells were seeded into a 96-well
plate with 1×104 cells/well and were subjected to
hypoxia (1% O2, 5% CO2 and 94% N2)
for 24 h. Following incubation with the drugs described above, MTT
and dimethyl sulfoxide were added sequentially and the optical
density (OD) was measured at 570 nm (DNM-9606 Enzyme Mark Analyzer;
Perlong Medical, Beijing, China).
Cell-survival rate calculation: Cell-survival rate
(%) = (ODsample −
ODblank)/(ODcontrol − ODblank) ×
100. ODblank refers to the value of culture medium mixed
with MTT without cells. In addition, the cells were visualized
microscopically (GX51; Olympus Corporation, Tokyo, Japan).
Statistical analysis
Statistical analysis was performed using GraphPad
Prism 5 software (GraphPad Software, Inc., La Jolla, CA, USA), and
all data are presented as the mean ± standard error of the mean. To
analyze the differences between groups, Student’s t-test for two
groups, analysis of variance for multiple group comparisons, and
Tukey’s test for repeated measures were conducted. P<0.05 was
considered to indicate a statistically significant difference.
Results
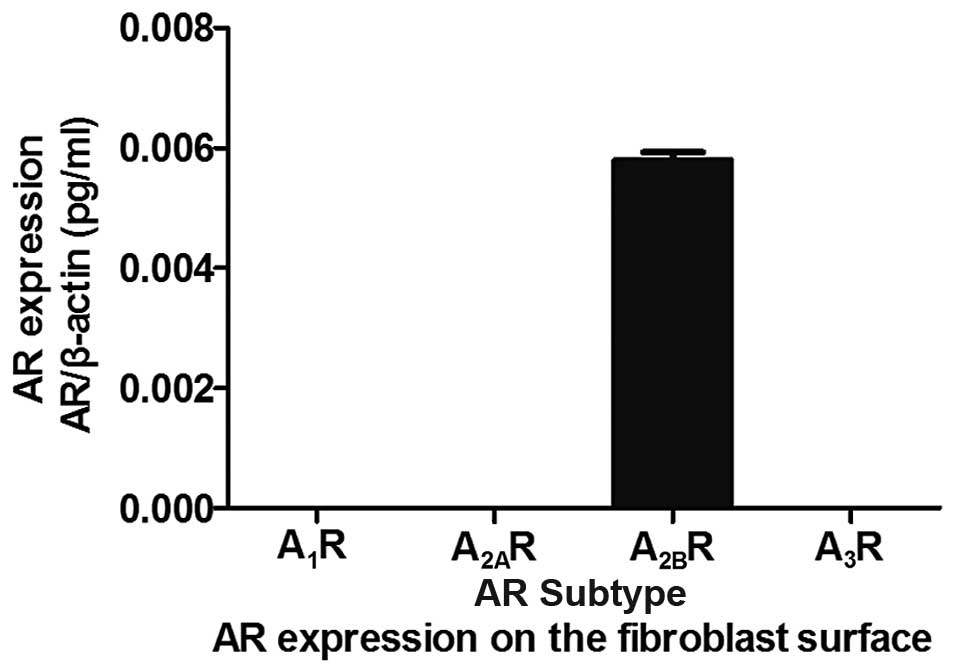
AR expression under hypoxia
The TaqMan probe-based assay demonstrated that the
predominantly expressed AR on mouse renal fibroblasts under
conditions of hypoxia was the A2BR subtype (Fig. 1).
Expression of transforming growth
factor-β1 (TGF-β1), procollagen α1 (I) and α-smooth muscle actin
(α-SMA)
TGF-β1 mRNA expression
Fig. 2A
demonstrates that a significant increase in the levels of TGF-β1
mRNA was observed in fibroblasts under hypoxia in the NECA group
(P<0.01). However, the PT and MRS groups were identified to have
significantly lower expression levels than the NECA group
(P<0.05) at days 2 and 3. No significant differences were
observed between the PT and MRS treatment groups (P>0.05).
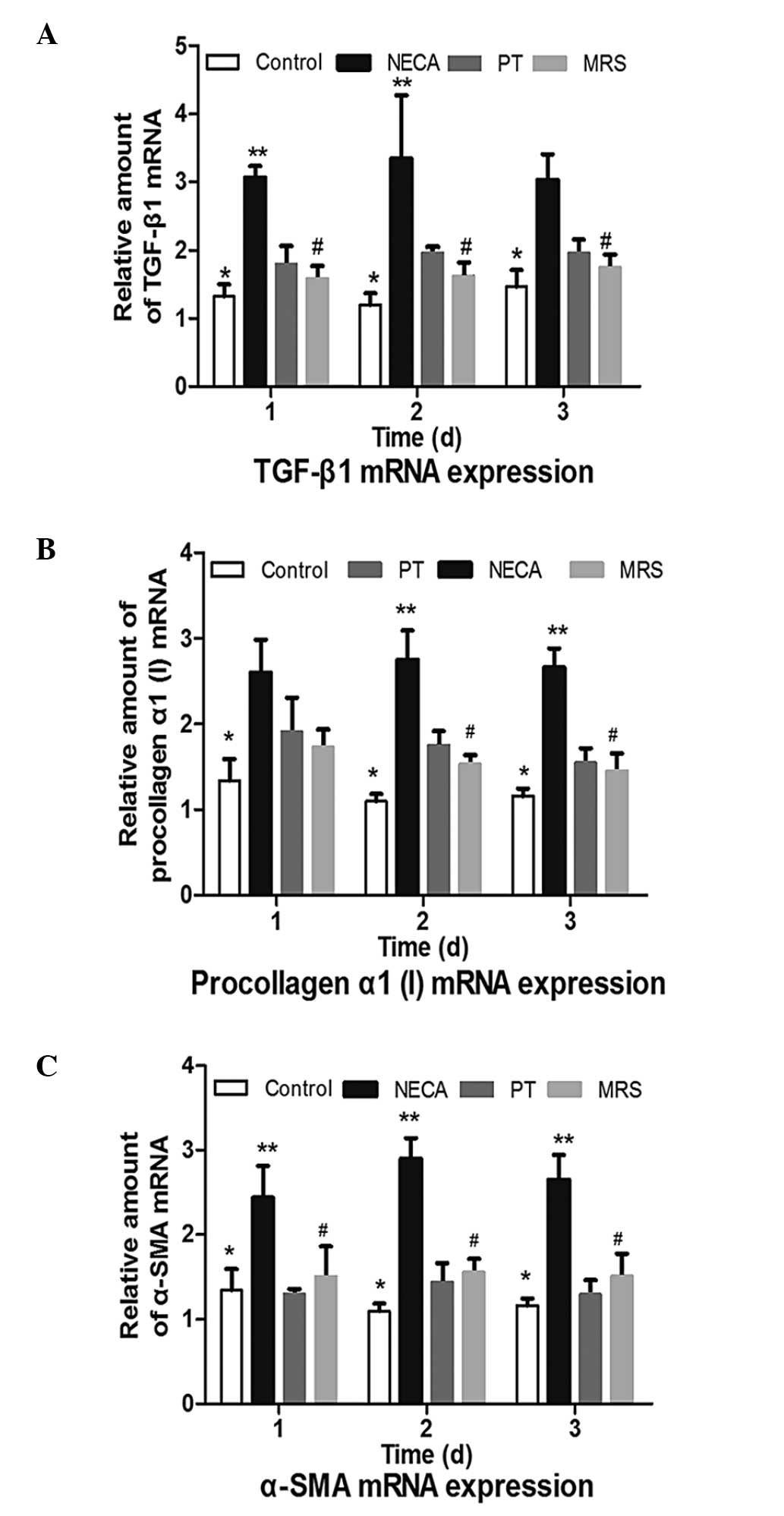 | Figure 2Expression of TGF-β1, procollagen α1
(I) and α-SMA mRNA expression levels in the control, PT, NECA and
MRS groups. (A) TGF-β1 mRNA expression. ADO significantly
upregulated TGF-β1 mRNA expression, while blocking A2BR
reversed TGF-β1 mRNA overexpression. (B) Procollagen α1 (I) mRNA
expression. ADO significantly upregulated procollagen α1 (I) mRNA
expression, while blocking A2BR reversed procollagen α1
(I) mRNA overexpression. (C) α-SMA mRNA expression. ADO
significantly upregulated α-SMA mRNA expression, while blocking
A2BR reversed the α-SMA mRNA overexpression.
*P<0.01 vs. the NECA group; **P<0.01 vs. the PT
group; #P<0.05 vs. the NECA group. TGF-β1,
transforming growth factor-β1; α-SMA, α-smooth muscle actin; ADO,
adenosine; NECA, 5′-N-ethylcarboxamidoadenosine; PT,
8-phenyltheophylline; MRS, MRS1754. |
Procollagen α1 (I) mRNA
expression
Fig. 2B
demonstrates that NECA significantly increased the levels of
procollagen α1 (I) mRNA expression (P<0.01). Compared with NECA,
procollagen α1 (I) mRNA expression was significantly reduced
following PT or MRS treatment on days 2 and 3 (P<0.05). PT and
MRS however, were not identified to significantly influence
procollagen α1 (I) mRNA expression levels (P>0.05), and no
differences were observed between the PT and MRS groups
(P>0.05).
α-SMA mRNA expression
As presented in Fig.
2C, the stimulation of NECA significantly induced the
expression of α-SMA (P<0.01). Compared with the control group,
there was no effect of PT or MRS on α-SMA mRNA expression levels
(P>0.05), which were significantly lower than in the NECA group
(P<0.05). No significant differences were identified between the
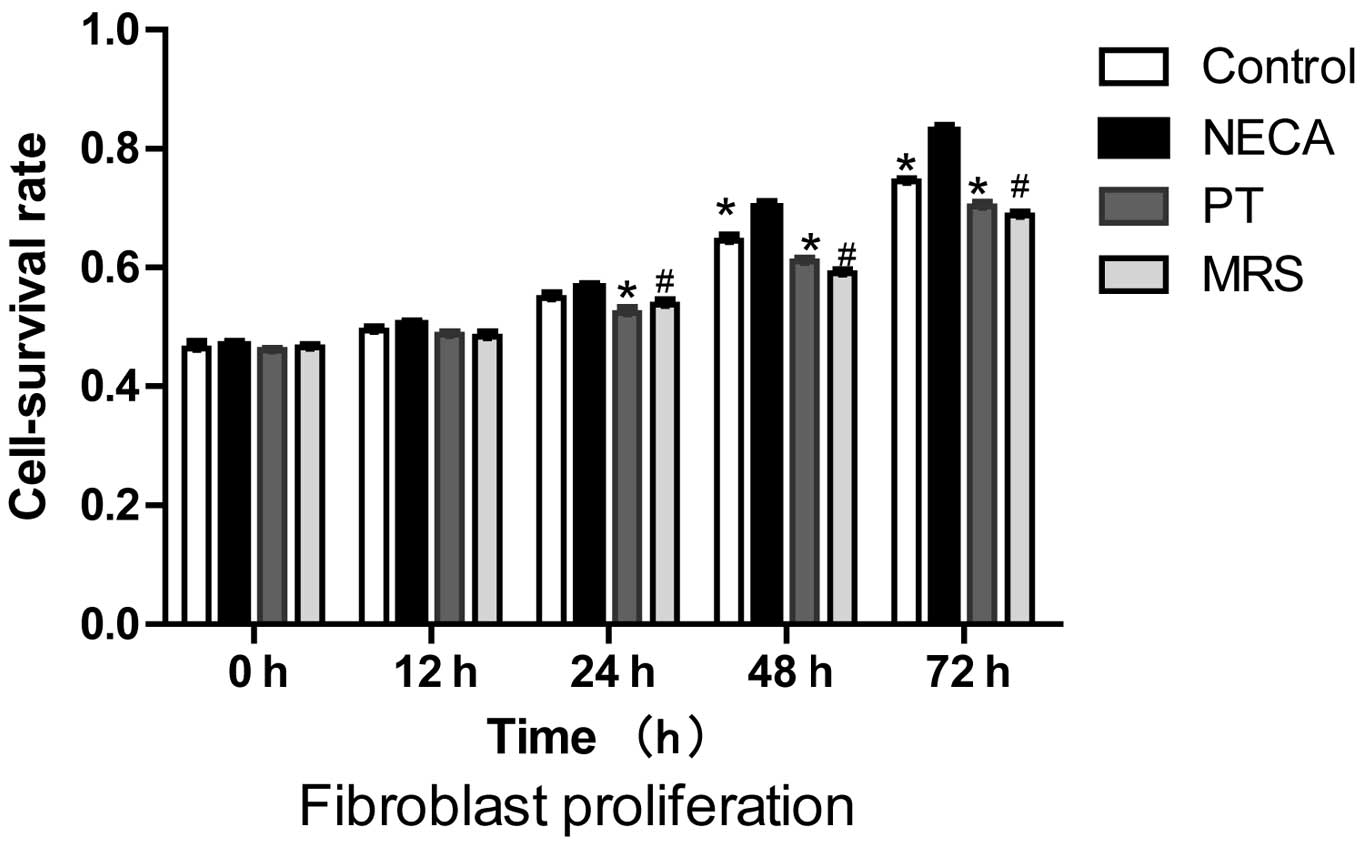
PT and MRS treatment groups (P>0.05). Proliferation of
fibroblasts. Proliferation of renal fibroblasts was measured
following 0, 12, 24, 48 and 72 h of hypoxia in triplicate (Fig. 3).
The cell survival rates of each group are presented
in Fig. 4. Compared with the
control group, NECA significantly induced renal fibroblast
proliferation at 48 and 72 h (P<0.001). The addition of 8-PT or
MRS 1754 was able to reverse the NECA-induced increase in renal
fibroblast proliferation at 24, 48 and 72 h (P<0.01).
Discussion
Fibroblast proliferation and activation are closely
associated with RIF development. Fibroblasts embedded within the
renal interstitium synthesize various components of the ECM,
including type I collagen, type III collagen and fibronectin
(14). The majority of fibroblasts
exist in a quiescent state and a small proportion remain active in
order to repair damaged tissues (15). However, in pathological conditions,
including hypoxic ischemia, inflammation and tissue damage, the
excretion of cytokines and ECM results in excessive fibroblast
proliferation and transdifferentiation to activated myofibroblasts.
As a result of the enhanced collagen production by myofibroblasts,
the ECM accumulation hinders normal renal function and results in
the development of RIF. In this pathological process, fibroblast
proliferation and activation are key to the development of RIF.
ADO is a chemical that is naturally present in all
cells and an increase in the cytoplasmic ADO concentration under
hypoxic ischemia is considered as a response to stress (16–18).
There are two key sources of cellular ADO: The degradation of ADO
triphosphate, and the dephosphorylation of adenosine monophosphate,
which is produced by the extracellular adenine nucleotide
metabolism (19). Extracellular
ADO exerts its biological effects by binding with AR on the cell
surface. Three types and four subtypes of AR have been identified,
including A1R, A2AR, A2BR and
A3R (20). ADO has been
reported to have vasodilatory effects in the majority of organs
(21), and protects against tissue
damage during acute ischemic injury in the heart (22), brain (23), liver (24) and kidney (25). However, extended exposure to a high
concentration of ADO has been demonstrated to lead to tissue damage
and increased organ dysfunction (26). In previous studies, it has been
identified that extracellular ADO continually increases during RIF,
and ADO serves an important function in RIF development and kidney
dysfunction (9,27). AR inhibitors, such as PT, have been
demonstrated to protect kidney function in UUO mice (9,27,28).
In order to further understand the function of the AR in RIF
development, a hypoxia mouse renal fibroblast model was generated
in the current study, in order to dynamically investigate the
function of ADO and the associated AR. In the present study, it was
observed that A2BR was the predominant receptor type on
the surface of the fibroblasts under hypoxia. The specific
A2BR inhibitor MRS1754 produced similar effects to the
global AR inhibitor 8-PT in renal fibroblast proliferation
inhibition (P>0.05), indicating that A2BR was the
predominant AR type during hypoxia and ADO increased fibroblast
proliferation by binding with A2BR.
During the development of RIF, fibroblasts exhibit
smooth muscle cell-like characteristics following a specific
transformation, producing myofibroblasts (29). In addition, α-SMA is a known marker
of this fibroblast phenotypic transformation (30). Data obtained from animals and
humans have clarified the common process of phenotypic
transformation of fibroblasts expressing α-SMA into myofibroblasts
during CKD development (31–35).
As the active form, myofibroblast are important in the progression
of fibrosis, by inducing excessive ECM accumulation (36,37).
The data from the current study demonstrated that compared with the
control group, NECA significantly increased α-SMA mRNA expression
levels (P<0.01). Compared with the NECA group, NECA combined
with 8-PT or MRS1754 reversed the NECA-induced upregulation of
α-SMA mRNA (P<0.05). The inhibition of global AR or
A2BR was able to reduce transformation of the fibroblast
phenotype, leading to the delay of RIF development and protection
of kidney function. No significant differences were observed
between 8-PT and MRS1754 in α-SMA expression, indicating that
A2BR was the predominant AR type responsible for
fibroblast transdifferentiation during hypoxia.
A previous study investigated the involvement of the
cytokine regulation network in the development of RIF (38). Under hypoxic ischemia, large
quantities of cytokines and chemotactic factors are excreted from
stressed cells, resulting in structural alterations and dysfunction
of the renal tissue. In the current study, two verified profibrotic
cytokines, TGF-β1 and procollagen α1 (I), were investigated. The
results demonstrated that compared with the control group, NECA
significantly increased TGF-β1 mRNA expression levels (P<0.01),
while 8-PT and MRS1754 were able to reverse NECA-induced
upregulation of TGF-β1 mRNA (P<0.05). Similar results were
observed in the evaluation of procollagen α1 (I) levels. The
results suggested that under hypoxia, ADO may accelerate RIF
development by inducing the excretion of pro-fibrotic cytokines.
However, the inhibition of global AR or A2BR was only
able to effectively alleviate disease by inhibiting cytokine
synthesis. No significant differences were observed between 8-PT
and MRS1754 treatment in the levels of TGF-β1 and procollagen α1
(I) regulation, indicating that A2BR was the predominant
receptor type for fibroblast profibrotic cytokine excretion under
hypoxia.
In conclusion, the current study demonstrated that
ADO was important in the regulation of biological behavior in
fibroblasts, and A2BR was identified as the predominant
receptor type on the surface of fibroblasts under conditions of
hypoxia. The inhibition of A2BR significantly reduced
the proliferation and activation of fibroblasts, and reduced the
excretion of profibrotic cytokines, thus preventing RIF
development. The current study suggests that A2BR may be
a novel target in the treatment of RIF.
Acknowledgments
The current study was supported by the Science and
Technology program of Hunan Scientific Committee (grant no.
2012FJ3134) and the National Natural Science Foundation of China
(grant no. 81470925).
References
|
1
|
Noronha IL, Fujihara CK and Zatz R: The
inflammatory component in progressive renal disease - are
interventions possible? Nephrol Dial Transplant. 17:363–368. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Badid C, Vincent M, Fouque D, Laville M
and Desmoulière A: Myofibroblast: A prognostic marker and target
cell in progressive renal disease. Ren Fail. 23:543–549. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Roberts IS, Burrows C, Shanks JH, Venning
M and McWilliam LJ: Interstitial myofibroblasts: Predictors of
progression in membranous nephropathy. J Clin Pathol. 50:123–127.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Razzaque MS and Taguchi T: Cellular and
molecular events leading to renal tubulointerstitial fibrosis. Med
Electron Microsc. 35:68–80. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zeisberg M, Strutz F and Müller GA: Renal
fibrosis: An update. Curr Opin Nephrol Hypertens. 10:315–320. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Eddy AA: Molecular basis of renal
fibrosis. Pediatr Nephrol. 15:290–301. 2000. View Article : Google Scholar
|
|
7
|
Fredholm BB: Adenosine, an endogenous
distress signal, modulates tissue damage and repair. Cell Death
Differ. 14:1315–1323. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Eltzschig HK, Thompson LF, Karhausen J,
Cotta RJ, Ibla JC, Robson SC and Colgan SP: Endogenous adenosine
produced during hypoxia attenuates neutrophil accumulation:
Coordination by extracellular nucleotide metabolism. Blood.
104:3986–3992. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Tang J, Jiang X, Zhou Y, Xia B and Dai Y:
Increased adenosine levels contribute to ischemic kidney fibrosis
in the unilateral ureteral obstruction model. Exp Ther Med.
9:737–743. 2015.PubMed/NCBI
|
|
10
|
Chunn JL, Mohsenin A, Young HW, Lee CG,
Elias JA, Kellems RE and Blackburn MR: Partially adenosine
deaminase-deficient mice develop pulmonary fibrosis in association
with adenosine elevations. Am J Physiol Lung Cell Mol Physiol.
290:L579–L587. 2006. View Article : Google Scholar
|
|
11
|
Chunn JL, Molina JG, Mi T, Xia Y, Kellems
RE and Blackburn MR: Adenosine-dependent pulmonary fibrosis in
adenosine deaminase-deficient mice. J Immunol. 175:1937–1946. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chunn JL, Young HW, Banerjee SK, Colasurdo
GN and Blackburn MR: Adenosine-dependent airway inflammation and
hyperresponsiveness in partially adenosine deaminase-deficient
mice. J Immunol. 167:4676–4685. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
14
|
Strutz F and Zeisberg M: Renal fibroblasts
and myofibroblasts in chronic kidney disease. J Am Soc Nephrol.
17:2992–2998. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chen CZ and Raghunath M: Focus on
collagen: in vitro systems to study fibrogenesis and antifibrosis
state of the art. Fibrogenesis Tissue Repair. 2:72007. View Article : Google Scholar
|
|
16
|
Blackburn MR, Volmer JB, Thrasher JL,
Zhong H, Crosby JR, Lee JJ and Kellems RE: Metabolic consequences
of adenosine deaminase deficiency in mice are associated with
defects in alveogenesis, pulmonary inflammation, and airway
obstruction. J Exp Med. 192:159–170. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Linden J: Adenosine in tissue protection
and tissue regeneration. Mol Pharmacol. 67:1385–1387. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Van Belle H, Goossens F and Wynants J:
Formation and release of purine catabolites during hypoperfusion,
anoxia, and ischemia. Am J Physiol. 252:H886–H893. 1987.PubMed/NCBI
|
|
19
|
Rego AC, Santos MS and Oliveira CR:
Adenosine triphosphate degradation products after oxidative stress
and metabolic dysfunction in cultured retinal cells. J Neurochem.
69:1228–1235. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wei CJ, Li W and Chen JF: Normal and
abnormal functions of adenosine receptors in the central nervous
system revealed by genetic knockout studies. Biochim Biophys Acta.
1808:1358–1379. 2011. View Article : Google Scholar
|
|
21
|
Mi T, Abbasi S, Zhang H, et al: Excess
adenosine in murine penile erectile tissues contributes to priapism
via A2B adenosine receptor signaling. J Clin Invest. 118:1491–1501.
2008. View
Article : Google Scholar : PubMed/NCBI
|
|
22
|
Rudolphi KA, Schubert P, Parkinson FE and
Fredholm BB: Adenosine and brain ischemia. Cerebrovasc Brain Metab
Rev. 4:346–369. 1992.PubMed/NCBI
|
|
23
|
Schubert P, Rudolphi KA, Fredholm BB and
Nakamura Y: Modulation of nerve and glial function by adenosine -
role in the development of ischemic damage. Int J Biochem.
26:1227–1236. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhu JF, Tang LM and Li DC: Activation of
adenosine 2A receptor attenuating oxidative stress on
small-for-size liver transplantation. Chin J Bases And Clin In Gen
Surg. 17:243–247. 2010.
|
|
25
|
Grenz A, Osswald H, Eckle T, Yang D, Zhang
H, Tran ZV, Klingel K, Ravid K and Eltzschig HK: The reno-vascular
A2B adenosine receptor protects the kidney from ischemia. PLoS Med.
5:e1372008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lu Q, Sakhatskyy P, Newton J, et al:
Sustained adenosine exposure causes lung endothelial apoptosis: a
possible contributor to cigarette smoke-induced endothelial
apoptosis and lung injury. Am J Physiol Lung Cell Mol Physiol.
304:L361–L370. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yap SC and Lee HT: Adenosine and
protection from acute kidney injury. Curr Opin Nephrol Hypertens.
21:24–32. 2012. View Article : Google Scholar :
|
|
28
|
Mubagwa K and Flameng W: Adenosine,
adenosine receptors and myocardial protection: an updated overview.
Cardiovasc Res. 52:25–39. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Leask A: Towards an anti-fibrotic therapy
for scleroderma: targeting myofibroblast differentiation and
recruitment. Fibrogenesis Tissue Repair. 3:82010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Malmström J, Lindberg H, Lindberg C, et
al: Transforming growth factor-beta 1 specifically induce proteins
involved in the myofibroblast contractile apparatus. Mol Cell
Proteomics. 3:466–477. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ohtaka A, Ootaka T, Sato H and Ito S:
Phenotypic change of glomerular podocytes in primary focal
segmental glomerulosclerosis: Developmental paradigm. Nephrol Dial
Transplant. 17(Suppl 9): 11–15. 2002. View Article : Google Scholar
|
|
32
|
Jinde K, Nikolic-Paterson DJ, Huang XR,
Sakai H, Kurokawa K, Atkins RC and Lan HY: Tubular phenotypic
change in progressive tubulointerstitial fibrosis in human
glomerulonephritis. Am J Kidney Dis. 38:761–769. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Johnson RJ, Iida H, Alpers CE, Majesky MW,
Schwartz SM, Pritzi P, Gordon K and Gown AM: Expression of smooth
muscle cell phenotype by rat mesangial cells in immune complex
nephritis. Alpha-smooth muscle actin is a marker of mesangial cell
proliferation. J Clin Invest. 87:847–858. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Ng YY, Fan JM, Mu W, Nikolic-Paterson DJ,
Yang WC, Huang TP, Atkins RC and Lan HY: Glomerular
epithelial-myofibroblast transdifferentiation in the evolution of
glomerular crescent formation. Nephrol Dial Transplant.
14:2860–2872. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Eddy AA: Molecular insights into renal
interstitial fibrosis. J Am Soc Nephrol. 7:2495–2508.
1996.PubMed/NCBI
|
|
36
|
Zhang G, Moorhead PJ and el Nahas AM:
Myofibroblasts and the progression of experimental
glomerulonephritis. Exp Nephrol. 3:308–318. 1995.PubMed/NCBI
|
|
37
|
Hewitson TD, Wu HL and Becker GJ:
Interstitial myofibroblasts in experimental renal infection and
scarring. Am J Nephrol. 15:411–417. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Ling H, Li X, Jha S, Wang W, Karetskaya L,
Pratt B and Ledbetter S: Therapeutic role of TGF-beta-neutralizing
antibody in mouse cyclosporin A nephropathy: Morphologic
improvement associated with functional preservation. J Am Soc
Nephrol. 14:377–388. 2003. View Article : Google Scholar : PubMed/NCBI
|