Introduction
A large number of myocardial cells die after acute
myocardial infarction (MI), which can result in ventricular
remodeling and heart failure (1).
Apoptosis is the predominant form of cardiomyocyte death, which
occurs in acute or chronic MI (2,3).
Therefore, greater understanding of the mechanism of apoptosis
following MI may aid in the development of effective therapy to
treat patients with MI.
Recent studies have shown that endoplasmic reticulum
stress (ERS) is an important pathway in cell apoptosis in addition
to the mitochondria pathway and death receptor pathway in heart
failure (4,5). ERS can be caused by a number of
disturbances, including hypoxia, nutritional deprivation, infection
and drug toxicity (6). When a
large amount of unfolded proteins accumulate in the ER, three ER
receptors are disassociated with GRP78 and then unfolded protein
response (UPR) is triggered (7).
Two independent pathways mediated by ER transmembrane proteins are
involved in the UPR protection mechanism. One pathway involves IRE1
and ATF6 restoring the unfolded or mis-folded protein to a
correctly folded protein by inducing the expression of chaperone
protein in the ER, which halts ERS and thus prevents cell apoptosis
(8). The other involves the
phosphorylation of eIF2α at serine residue 51 by pancreatic
endoplasmic reticulum eIF2α kinase (PERK) and phosphorylated eIF2α
decreases the unfolded protein level by preventing protein
synthesis (9,10). When these two pathways were
inhibited, the apoptosis-related proteins, such as caspase-12,
GADD/CHOP and ASK1/JNK, were activated and ERS-induced apoptosis
was then triggered (9). The
ER-related apoptotic pathway has been reported to be one of
mechanisms underlying the induction of MI injury in cardiomyocytes
(11).
Salubrinal is a small molecular compound that can
inhibit the dephosphorylation and maintain the phosphorylation
state of eIF2α, resulting in the suppression of protein translation
and a decrease in protein synthesis to maintain homestasis in the
ER (12,13). The protective effect produced by
salubrinal has been verified in a number of studies (14–16).
Salubrinal protects against cell death, which was mediated by
tunicamycin-induced ERS at certain concentrations in the PC-12 rat
cell line (17). Gasparetto et
al (18) found that salubrinal
can be used to reduce nephrotoxicity induced by
cyclosporine-mediated ERS in rats. In addition, salubrinal has also
been used for the treatment of urinary system diseases (19) and diabetes (20). Recently, Liu et al (21) have suggested that salubrinal can
protect against tunicamycin- and hypoxia-induced cardiomyocyte
apoptosis via the PERK-eIF2α signaling pathway. In the heart
failure rat model, salubrinal treatment reduced apoptosis and
increased the levels of eIF2α and caspase-12 (16). However, to the best of our
knowledge, there has been no study investigating the effects of
salubrinol in MI.
Thus, in the present study, ERS was observed in a
rat MI model. Salubrinal was used as an in vivo treatment
and eIF2α phosphorylation was detected and its role in MI was
evaluated.
Materials and methods
All studies were approved by the Chinese PLA General
Hospital Ethics Committee and performed in accordance with the
ethical standards of the Authorization for Practicals on Animals at
our hospital. All the experiments were conducted in accordance with
the Guide for the Care and Use of Laboratory Animals and approved
by the ethical committee of Academy of Military Medical
Science.
MI rat model
Male Wistar rats weighing 220–260 g (Animal Center
affiliated to Academy of Military Medical Science, Beijing, China)
were randomly divided into the MI group (n=75) and sham group
(n=75). Each group was further divided into five subgroups (15 rats
per group), which were anaesthetized and sacrificed at 1, 3, 6, 12
and 24 h, respectively. In the salubrinal-treated experiments, an
additional 60 rats were equally and randomly divided into the sham
group, MI group and salubrinal-treated group (n=20). Heart tissue
analysis in the sham group, MI group and salubrinal group was
performed after 24 h administration.
Rats were anaesthetized with intraperitoneal (ip)
injection of 2% pentobarbital natrium (40 mg/kg, Shanghai Second
Chemical Reagent Company, Shanghai, China). The rats were placed in
the supine position. A left thoracotomy was performed and the left
anterior descending (LAD) coronary artery was ligated at the root
of the left coronary artery as previously reported (22). Successful coronary occlusion was
confirmed by a typical S-T segment elevation on the
electrocardiogram, whitened myocardial tissue and decreased
regional myocardial velocity, as determined by magnetic resonance
imaging, which was performed on a Picker 1.5-T scanner (Picker
International, Highland Heights, OH, USA), as described previously
(23). No LAD coronary artery
ligation was performed in the sham group. Furthermore, in the
salubrinal treated group, rats received ip injection of salubrinal
(1 mg/kg body weight, Sigma-Aldrich, St. Louis, MO, USA) for 30 min
prior to LAD ligation. Salubrinal was dissolved in dimethy
sulfoxide (Amresco, Solon, OH, USA) and then in saline. The MI
group with ip injection of equal volume of DMSO and saline was set
as the control.
Hematoxylin and eosin (H&E), terminal
deoxynucleotidyl transferase-mediated dUTP nick end labeling
(TUNEL) and triphenyltetrazolium chloride (TTC) staining
The heart tissues from each group were fixed in 10%
neutral formaldehyde and then stained with H&E (Beijing
Chemical Reagent Company, Beijing, China). Images were captured
using a microscope (BX51; Olympus Corporation, Tokyo, Japan).
The pretreated heart tissue slices were processed
with TUNEL staining according to the manufacturer’s instructions
(Roche, Basel, Switzerland). The slices were analyzed under a light
microscope with a magnification of ×400. Five non-overlapping
fields were randomly selected and the cells with brown particles
observed in the nucleus were recognized as positive cells. The
following equation was used for the apoptotic index (AI)
calculation: AI=(Positive cell numbers in one field/Total cell
numbers in one field) × 100.
The hearts was rapidly removed and cooled in
ice-cold saline for 10 min. Left ventricle (2 mm) was cut and
immersed in 1% TTC (Sigma-Aldrich) at 37°C for 30 min, and then
transferred to 4% paraformaldehyde in 0.1 M PBS (pH 7.4) for 24 h
fixation. Normal hearts were red while MI hearts were white. The
heart slices were photographed and analyzed with Image-Pro Plus 6.0
(Media Cybernetics, Inc., Rockville, MD, USA).
Lactate dehy drogenase (LDH), creatine
kinase (CK) and superoxide dismutase (SOD) activity
LDH, CK and SOD activity in myocardial tissue were
measured using LDH, CK and SOD Detection kits (Nanjing Jiancheng
Bioengineering Institute, Nanjing, China) according to the
manufacturer’s instructions.
Reverse transcription-quantitative
polymerase chain reaction
Total RNA was extracted from myocardial tissue by
TRIzol reagent (Invitrogen Life Technologies, Carlsbad, CA, USA)
and DNA was removed by DNase (Sigma-Aldrich). The reverse
transcription reaction was performed according to the
manufacturer’s instructions. The GRP78, caspase-12 and CHOP
specific sequences were amplified during 45 cycles of 20 sec
denaturing at 95°C, 25 sec annealing at 59°C, and 30 sec extension
at 72°C, with the primers listed in Table I. GAPDH was used as an internal
control and the 2−ΔΔCt method was used to calculate
expression levels.
 | Table ISequence of the primers. |
Table I
Sequence of the primers.
| Primer name | Forward (5′-3′) | Reverse (5′-3′) |
|---|
| Caspase-12 |
CTCTTCATTTCCAAACTCGTTGACT |
GGGCATCTGGGTCAGTTCAC |
| GADD153/CHOP |
AGGAGGTCCTGTCCTCAGATGA |
ATGTGCGTGTGACCTCTGTTG |
| GRP78 |
TGACCAAAACCGCCTGACA |
TCTCCAATCTGGTTCTTGAGAGAA |
| GAPDH |
GGTGAAGGTCGGTGTGAACG |
CTCGCTCCTGGAAGATGGTG |
Western blot analysis
The frozen myocardial tissues were lysed in
radioimmunoprecipitation cell lysis solution (Beyotime
Biotechnology, Shanghai, China), followed by high speed
centrifugation (13,000 × g, 5 min) and β-cyanoalanine
quantification. Cellular protein was separated by electrophoresis
on SDS-PAGE gel and then transferred onto polyvinylidene difluoride
membrane (Merck Millipore, Darmstadt, Germany). After blocking, the
blots were incubated with antibodies against GRP78 (N-20) (cat. no.
sc-1050; 1/600; Santa Cruz Biotechnology Inc., Santa Cruz, CA,
USA), p-eIF2α (Ser49) (cat. no. sc-293100; 1/200; Santa Cruz
Biotechnology Inc.), caspase-12 (A-14) (cat. no. sc-12395; 1/400;
Santa Cruz Biotechnology Inc.) and GADD153 (F-168) (cat. no.
sc-575; 1/500; Santa Cruz Biotechnology Inc.) at 4°C overnight. The
eIF2α (FL-315) (cat. no. sc-11386; 1/500; Santa Cruz Biotechnology
Inc.) and actin (C-11) (cat. no. sc-1615; 1/500; Santa Cruz
Biotechnology Inc.) were used as loading controls. The appropriate
horseradish peroxidase-conjugated secondary antibodies were added
and the samples were incubated for 1 h at room temperature (1:6,000
dilution for p-eIF2a and caspase-12 and 1:10,000 dilution for
GRP78, eIF2a, GADD153 and actin). The protein bands were detected
with SuperSignal Ultra Chemiluminescent Substrate (Pierce,
Rockford, IL, USA) on X-ray films (Koda).
Statistical analysis
Statistical analysis was performed by SPSS11.5
software (SPSS Inc., Chicago, IL, USA). The data are presented as
the mean ± standard deviation. One way-analysis of variance was
used to examine the differences between three or more groups.
P<0.05 was considered to indicate a statistically significant
difference.
Results
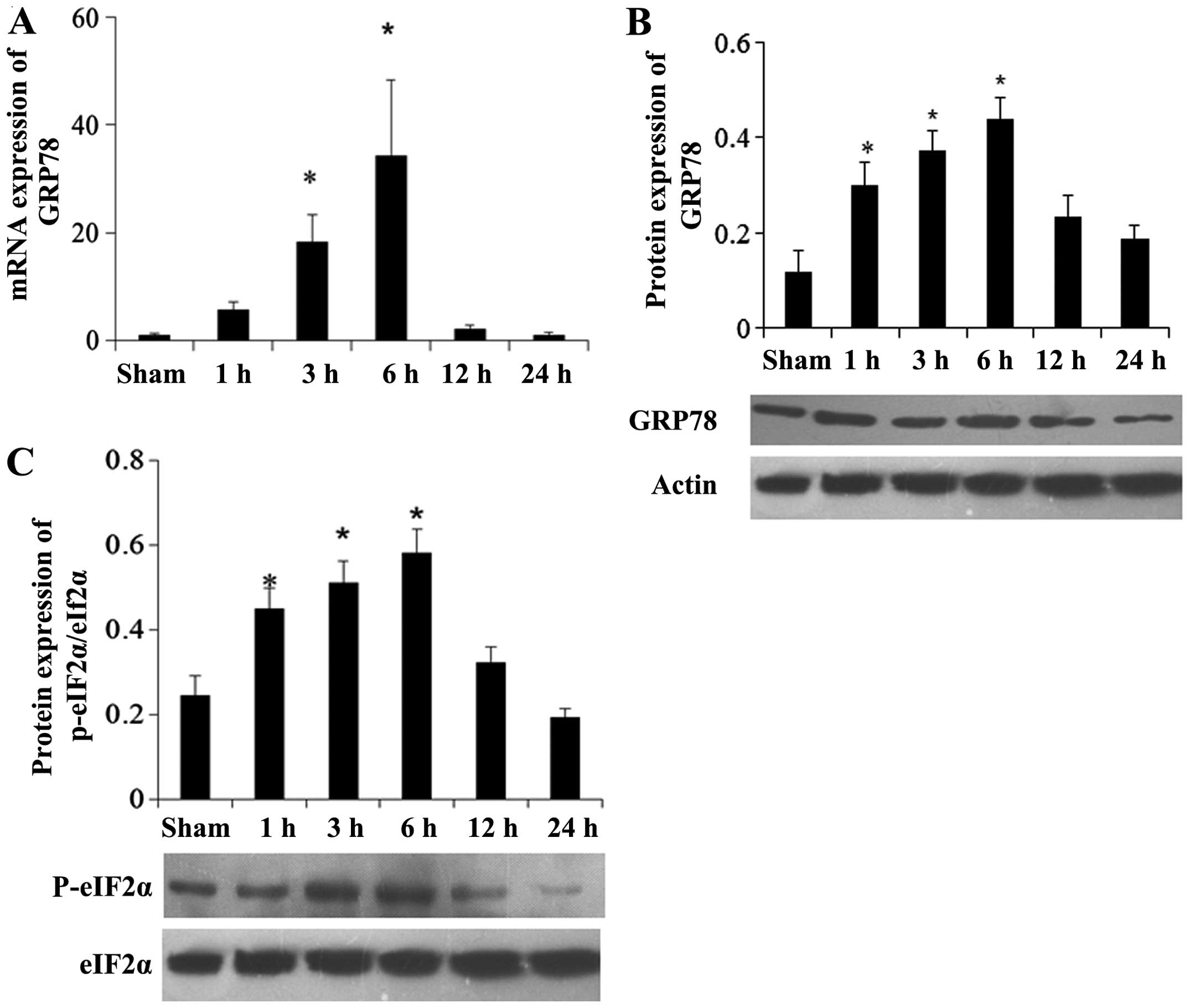
UPR of ER responds to MI
After successful establishment of the MI rat model,
RT-qPCR and western blot analysis were used to analyze the
expression of GRP78 in the myocardial tissues. Compared with the
sham group, the mRNA expression level of GRP78 in the MI group was
increased during the early period after MI and decreased after 6 h
of MI (Fig. 1A). Furthermore, the
western blot analysis results showed that no significant difference
in the GRP78 protein level was identified among the different time
points in the sham group; thus, the GRP78 protein level at the 24 h
time point in sham group was used as a reference. Compared with the
sham group, GRP78 protein in the MI group was expressed 1 h
following the establishment of MI and reached a peak after 6 h
(Fig. 1B). A similar outcome was
demonstrated via immunohistochemistry (data not shown).
Furthermore, the phosphorylation level of eIF2α was
detected by western blot analysis and immunohistochemistry. Western
blot analysis results showed that p-eIF2α protein expression was
increased and reached a peak 6 h after MI (Fig. 1C), indicating that UPR was induced
at early stages following establishment of MI. Immunohistochemistry
experiments showed similar results (data not shown).
ER stress-induced apoptosis pathway is
activated after MI
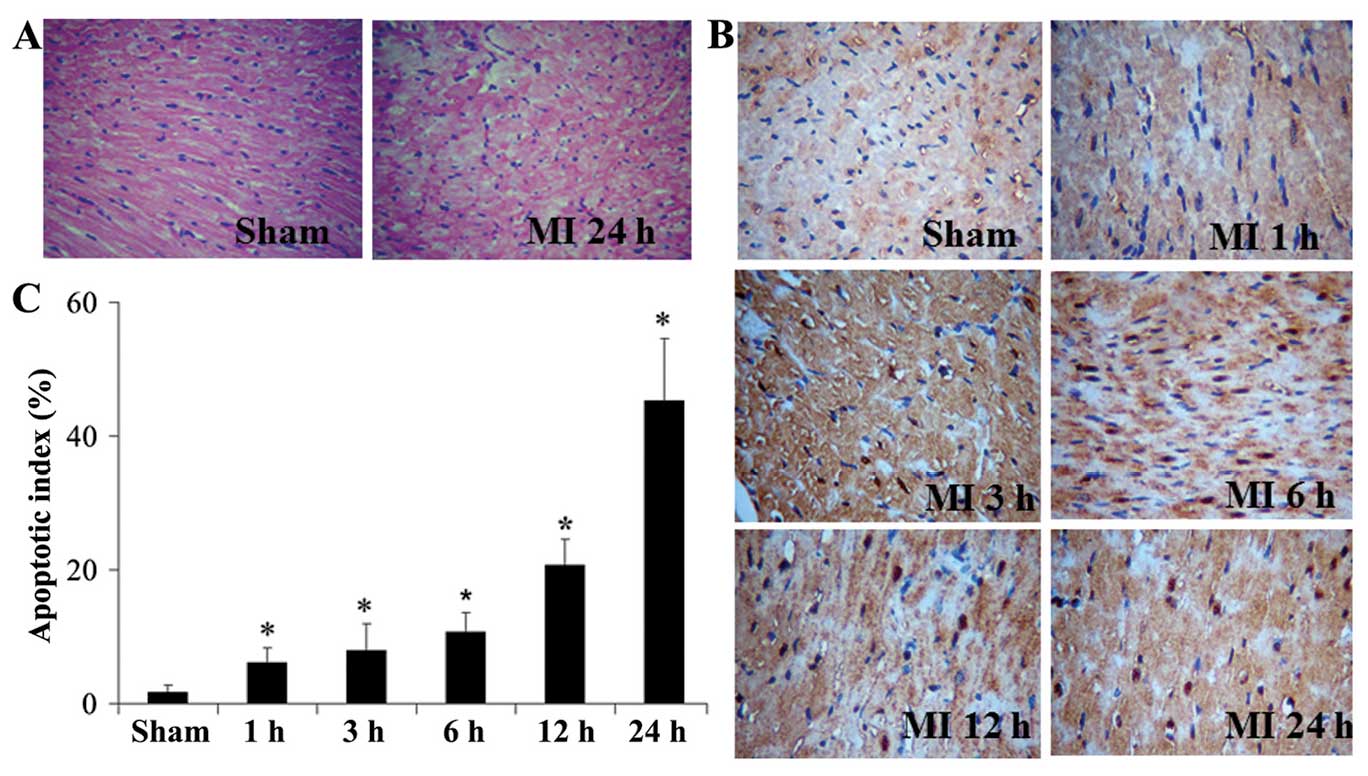
According to the H&E staining results,
myocardial injury was observed in the MI group, while no
significant difference was observed among different time points in
the sham group (Fig. 2A). TUNEL
staining also showed no difference in AI among different time
points in the sham group (P>0.05). However, in the MI group, AI
was increased with time and was significantly higher compared with
that in the sham group following MI (P<0.05, Fig. 2B and C).
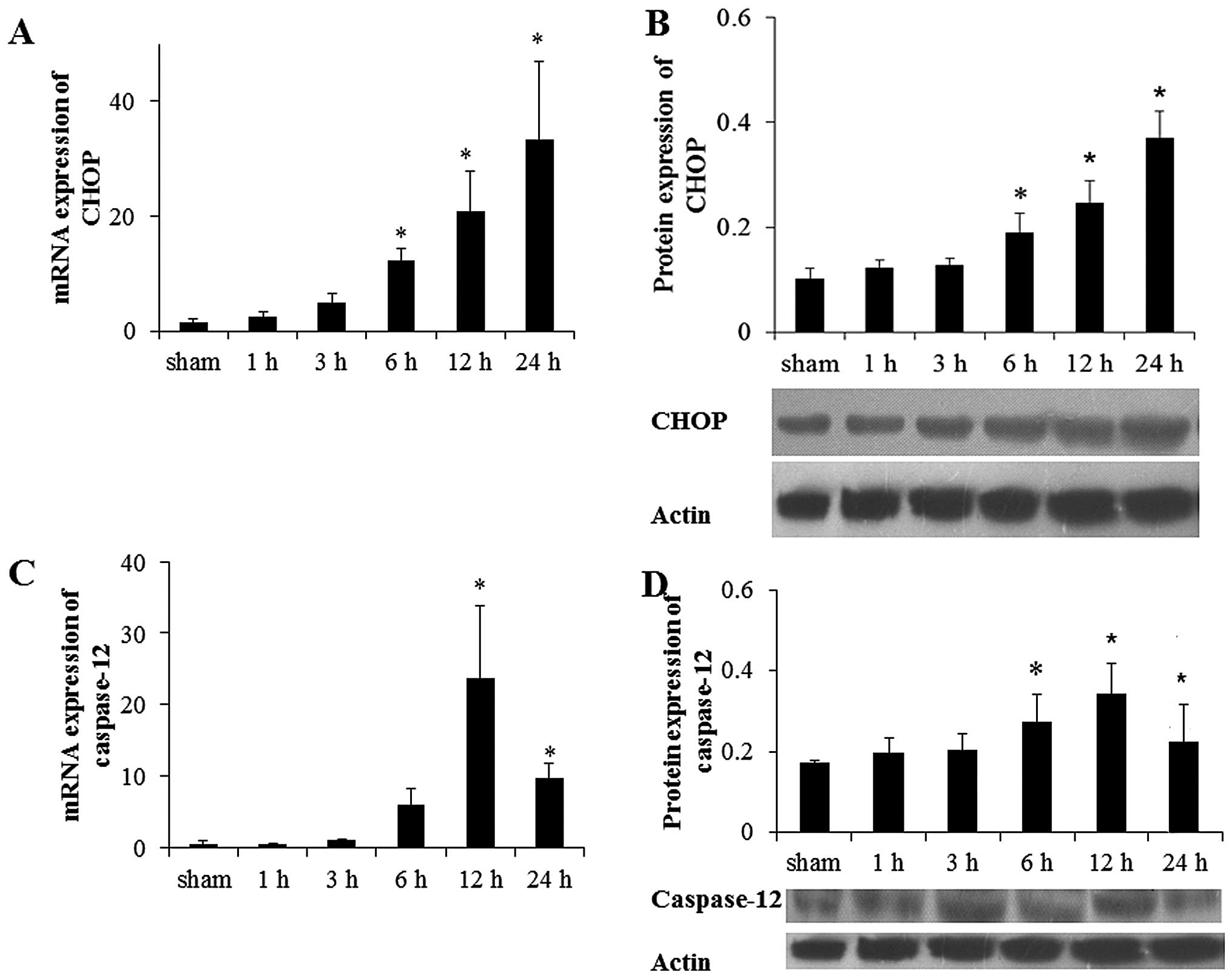
Subsequently, the mRNA and protein expression levels
of CHOP and caspase-12 protein, which were involved in the
apoptotic pathway induced by ER were investigated. Compared with
the sham group, the mRNA level of CHOP and caspase-12 were
increased 6 h after MI and reached a peak at 24 and 12 h after MI,
respectively (Fig. 3A and B).
Compared with the sham group, the CHOP protein level was increased
significantly at 6 h after MI and reached a peak at 24 h
(P<0.05; Fig. 3C). The protein
level of caspase-12 was increased 6 h after MI and declined
significantly at 24 h after MI (Fig.
3D). Similar results were detected via immunohistochemistry
(P<0.05; data not shown).
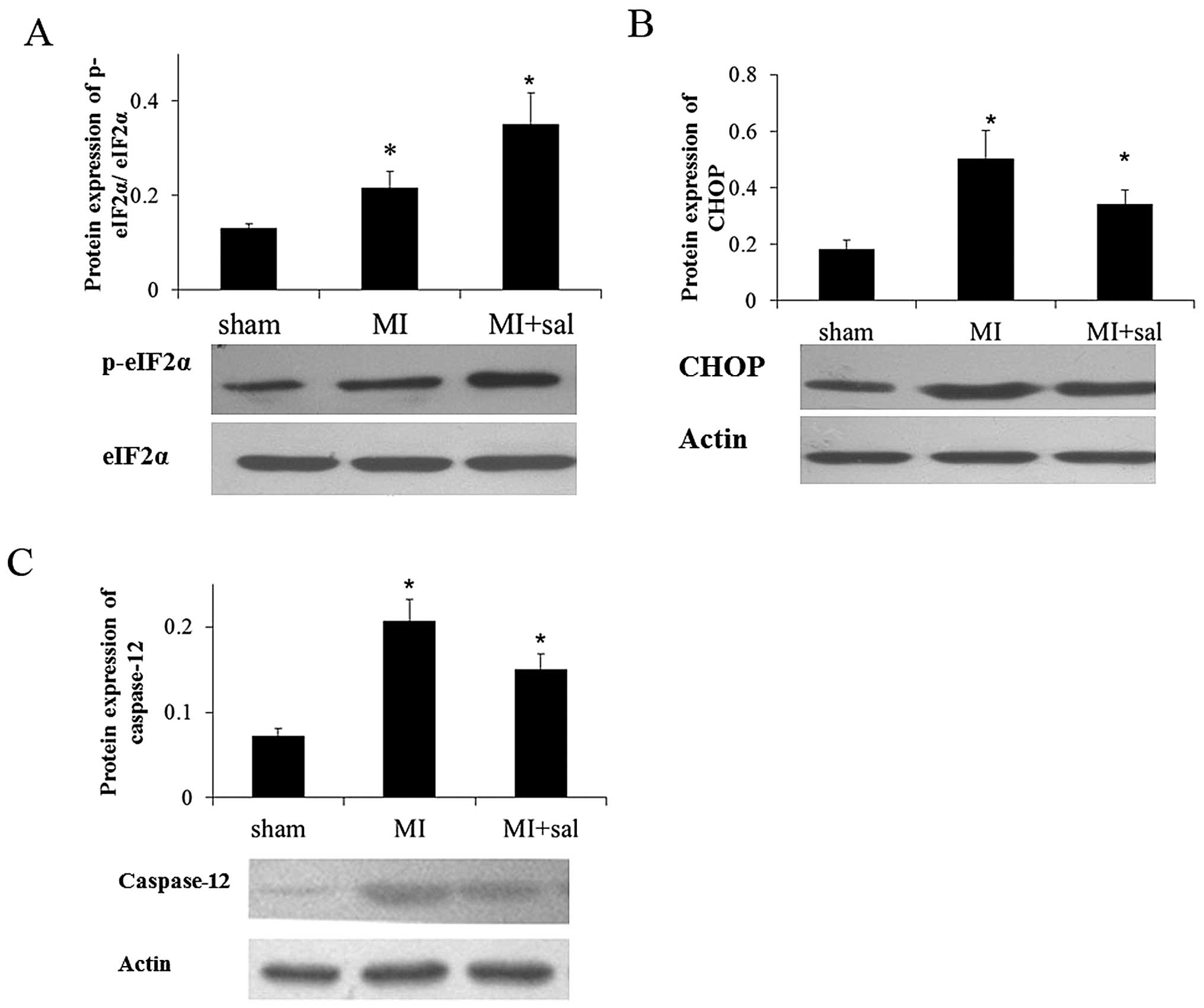
Salubrinal increases the phosphorylation
level of eIF2α and decreases the protein level of CHOP and
caspase-12
Salubrinal was administered to MI rats. In the
salubrinal treated group, the p-eIF2α expression level was
increased significantly compared with the MI group, which was
verified by western blot analysis (P<0.05) (Fig. 4A). Furthermore, the protein levels
of CHOP and caspase-12 were decreased significantly in the
salubrinal treated group compared with the MI group (P<0.05)
(Fig. 4B and C).
Immunohistochemistry experiments demonstrated the same results
(data not shown).
Ischemic damage induced by MI is
alleviated by suppression of eIF2α phosphorylation
According to H&E staining, the myocardial damage
in the salubrinal treated group was alleviated compared with the MI
group. The MI size assessed by the TTC staining was decreased
significantly in the salubrinal treated group compared with the MI
group (P<0.05, Fig. 5A). TUNEL
staining showed similar results (Fig.
5B). The activities of CK and LDH were decreased and SOD was
increased in the salubrinal treated group, compared with the MI
group (Fig. 5C).
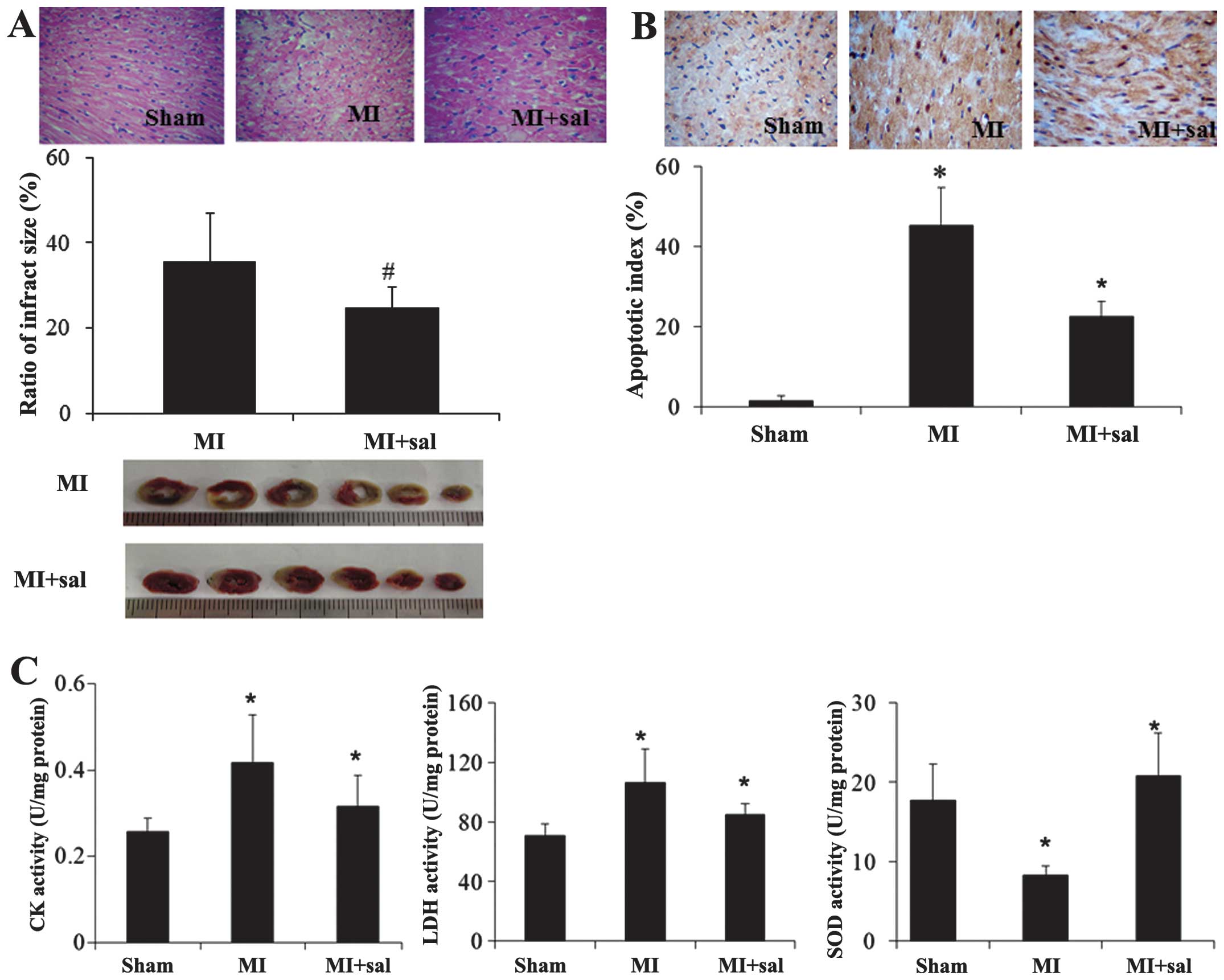 | Figure 5Ischemic damage induced by MI was
decreased by suppressing eIF2α. The effect of salubrinal (sal)
treatment on the myocardium was observed by (A) hematoxylin-eosin
staining and triphenyltetrazolium chloride staining, (B) terminal
deoxynucleotidyl transferase-mediated dUTP nick end labeling
(original magnification, ×400), and (C) myocardial enzyme detection
(CK, LDH, SOD). The representative images from three independent
experiments were presented. Data were presented as mean ± standard
deviation (*P<0.05 vs. sham; #P<0.05
vs. MI). MI, myocardial infarction; eIF2α, eukaryotic translation
initiation factor 2α; LDH, lactate dehy drogenase; CK, creatine
kinase; SOD, superoxide dismutase. |
Discussion
To the best of our knowledge, the present study is
the first to demonstrate that salubrinal protects cardiomyocytes
against apoptosis induced by MI. It was shown that administration
of salubrinal protected cardiomyocytes from injury induced by LAD
ligation in a rat model. Enhanced eIF2α phosphorylation and
decreased MI size were observed in the salubrinal treated group.
These findings indicated that the protective effect of salubrinal
in the rat MI model was mediated by suppressing the
dephosphorylation of eIF2α.
When ER dysfunction occurs, GRP78 is released and
interacts with unfolded or mis-folded protein to prevent protein
accumulation in the ER. As GRP78 is induced in response to ER
dysfunction, it was used as a marker to indicate the events of ERS
and UPR (24). In the present
study, the mRNA level of GRP78 increased significantly and reached
a peak at 6 h, and the GRP78 protein was expressed 1 h after MI and
also reached a peak at 6 h in the MI group. These results suggested
that UPR was induced early after MI. The results were consistent
with a previous study, which demonstrated that GRP48 expression was
increased in cardiac myocytes near the MI (25). In addition, p-eIF2α protein
expression was increased and reached a peak 6 h after MI. The
increase in p-eIF2α expression was also observed following heart
failure (16). All the results
showed that the ER was activated after MI.
When ER stress occurred, caspase-12 was activated,
which mediates ER-specific apoptosis (26–28).
Therefore, caspase-12 can be recognized as a specific marker of
ERS-induced apoptosis. It was demonstrated that the expression of
caspase-12 was elevated 6 h after MI and gradually increased with
time. These results indicated that UPR was induced to respond to
ERS during the early stage after MI, in which no caspase-12 was
detected. However, with the processing of ERS, caspase-12 was
activated and cell death was observed. Similar results were shown
in a previous study where the protein expression of caspase-12 was
increased in a heart failure rat model, which was induced by MI
(16).
CHOP is also termed GADD153, the activation of which
is one of the ER apoptosis pathways (29). Studies have demonstrated that CHOP
can be used as a specific marker in ERS induced apoptosis (30). In the present study, it was
demonstrated that the protein level of CHOP began to increase 6 h
after MI and reached a peak at 24 h. No CHOP expression was
observed during the early stages of MI as UPR was induced. With the
development of MI, CHOP was activated and apoptosis was
triggered.
When MI model rats were treated with salubrinal,
infarct size was reduced and apoptosis was alleviated. Similarly,
the activities of LDH and CK, which have high specificity for
cardiac injury and infarction, were decreased following
administration of salubrinal. Antioxidant enzyme SOD activity was
decreased after MI and increased after salubrinal treatment, which
has been reported to be impaired in MI (31). The results showed that salubrinal
alleviated the ischemic damage. In salubrinal treatment, expression
levels of CHOP and caspase-12 were decreased. In addition, the
phosphorylation level of eIF2α was increased following salubrinal
treatment. It has been reported that salubrinal could inhibit the
dephosphorylation of eIF2α (32).
In HK-2 human renal proximal tubular cells exposed to cadmium
chloride, apoptosis was induced and salubrinal could reduce the
expression of CHOP expression (33). In heart failure, caspase-12 was
increased in apoptotic cardiac myocytes and significantly decreased
after salubrinal treatment (16).
In heart failure, the phosphorylation of eIF2α was decreased and
increased further after salubrinal treatment (16,34).
Thus, salubrinal could protect cardiac myocytes against ERS
apoptosis after MI.
In conclusion, using the rat LAD ligation model, it
was found that ERS was induced in the cardiomyocyte apoptosis
following MI. UPR was found to maintain ER homestasis at the early
stages of MI and then the apoptosis related pathway was activated.
eIF2α may therefore be a candidate gene for an MI therapeutic
target as the eIF2α inhibitor salubrinal in the rat model can
alleviate the damage of MI. However, these results require further
confirmation.
Acknowledgments
This study was supported by the Ministry Science
Foundation of the Chinese People’s Liberation Army during the 12th
Five-Year Plan Period (grant no. BWS12J048) and Major International
Science and Technology Cooperation Projects (grant no.
2013DFA31170).
References
|
1
|
Gajarsa JJ and Kloner R: Left ventricular
remodeling in the post-infarction heart: A review of cellular,
molecular mechanisms, and therapeutic modalities. Heart Fail Rev.
16:13–21. 2011. View Article : Google Scholar
|
|
2
|
Abbate A, Salloum FN, Vecile E, et al:
Anakinra, a recombinant human interleukin-1 receptor antagonist,
inhibits apoptosis in experimental acute myocardial infarction.
Circulation. 117:2670–2683. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Takemura G and Fujiwara H: Role of
apoptosis in remodeling after myocardial infarction. Pharmacol
Ther. 104:1–16. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Schröder M: Endoplasmic reticulum stress
responses. Cell Mol Life Sci. 65:862–894. 2008. View Article : Google Scholar
|
|
5
|
Miyazaki Y, Kaikita K, Endo M, et al:
C/EBP homologous protein deficiency attenuates myocardial
reperfusion injury by inhibiting myocardial apoptosis and
inflammation. Arterioscler Thromb Vasc Biol. 31:1124–1132. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Xu C, Bailly-Maitre B and Reed JC:
Endoplasmic reticulum stress: cell life and death decisions. J Clin
Invest. 115:2656–2664. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lee AS: The ER chaperone and signaling
regulator GRP78/BiP as a monitor of endoplasmic reticulum stress.
Methods. 35:373–381. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lin JH, Li H, Yasumura D, et al: IRE1
signaling affects cell fate during the unfolded protein response.
Science. 318:944–949. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lai EWS: Diabetes and endoplasmic
reticulum stress in pancreatic β-cells: effects on insulin
biosynthesis and β-cell apoptosis. Master of Science dissertation.
University of Toronto; Toronto, Canada: 2008
|
|
10
|
Ron D: Translational control in the
endoplasmic reticulum stress response. J Clin Invest.
110:1383–1388. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Mitra A, Basak T, Datta K, Naskar S,
Sengupta S and Sarkar S: Role of α-crystallin B as a regulatory
switch in modulating cardiomyocyte apoptosis by mitochondria or
endoplasmic reticulum during cardiac hypertrophy and myocardial
infarction. Cell Death Dis. 4:e5822013. View Article : Google Scholar
|
|
12
|
Boyce M, Bryant KF, Jousse C, et al: A
selective inhibitor of eIF2alpha dephosphorylation protects cells
from ER stress. Science. 307:935–939. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wiseman RL and Balch WE: A new
pharmacology-drugging stressed folding pathways. Trends Mol Med.
11:347–350. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhu Y, Fenik P, Zhan G, Sanfillipo-Cohn B,
Naidoo N and Veasey SC: Eif-2a protects brainstem motoneurons in a
murine model of sleep apnea. J Neurosci. 28:2168–2178. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Sokka AL, Putkonen N, Mudo G, et al:
Endoplasmic reticulum stress inhibition protects against
excitotoxic neuronal injury in the rat brain. J Neurosci.
27:901–908. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu Y, Wang J, Qi SY, et al: Reduced
endoplasmic reticulum stress might alter the course of heart
failure via caspase-12 and JNK pathways. Can J Cardiol. 30:368–375.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Paschen W: Endoplasmic reticulum: a
primary target in various acute disorders and degenerative diseases
of the brain. Cell Calcium. 34:365–383. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Gasparetto M, Gentry T, Sebti S, et al:
Identification of compounds that enhance the anti-lymphoma activity
of rituximab using flow cytometric high-content screening. J
Immunol Methods. 292:59–71. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Katsoulieris E, Mabley JG, Samai M, Green
IC and Chatterjee PK: alpha-Linolenic acid protects renal cells
against palmitic acid lipotoxicity via inhibition of endoplasmic
reticulum stress. Eur J Pharmacol. 623:107–112. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Cnop M, Ladriere L, Hekerman P, et al:
Selective inhibition of eukaryotic translation initiation factor 2
alpha dephosphorylation potentiates fatty acid-induced endoplasmic
reticulum stress and causes pancreatic beta-cell dysfunction and
apoptosis. J Biol Chem. 282:3989–3997. 2007. View Article : Google Scholar
|
|
21
|
Liu CL, Li X, Hu GL, et al: Salubrinal
protects against tunicamycin and hypoxia induced cardiomyocyte
apoptosis via the PERK-eIF2α signaling pathway. J Geriatr Cardiol.
9:258–268. 2012.PubMed/NCBI
|
|
22
|
Martindale JJ, Wall JA, Martinez-Longoria
DM, et al: Overexpression of mitogen-activated protein kinase
kinase 6 in the heart improves functional recovery from ischemia in
vitro and protects against myocardial infarction in vivo. J Biol
Chem. 280:669–676. 2005. View Article : Google Scholar
|
|
23
|
Karwatowski SP, Mohiaddin RH, Yang GZ,
Firmin DN, St John Sutton M and Underwood SR: Regional myocardial
velocity imaged by magnetic resonance in patients with ischaemic
heart disease. Br Heart J. 72:332–338. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Schröder M and Kaufman RJ: The mammalian
unfolded protein response. Annu Rev Biochem. 74:739–789. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Thuerauf DJ, Marcinko M, Gude N, Rubio M,
Sussman MA and Glembotski CC: Activation of the unfolded protein
response in infarcted mouse heart and hypoxic cultured cardiac
myocytes. Circ Res. 99:275–282. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ron D and Walter P: Signal integration in
the endoplasmic reticulum unfolded protein response. Nat Rev Mol
Cell Biol. 8:519–529. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
27
|
Walter P and Ron D: The unfolded protein
response: from stress pathway to homeostatic regulation. Science.
334:1081–1086. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Nakagawa T, Zhu H, Morishima N, et al:
Caspase-12 mediates endoplasmic-reticulum-specific apoptosis and
cytotoxicity by amyloid-beta. Nature. 403:98–103. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Oyadomari S and Mori M: Roles of
CHOP/GADD153 in endoplasmic reticulum stress. Cell Death Differ.
11:381–389. 2004. View Article : Google Scholar
|
|
30
|
Benavides A, Pastor D, Santos P, Tranque P
and Calvo S: CHOP plays a pivotal role in the astrocyte death
induced by oxygen and glucose deprivation. Glia. 52:261–275. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Prabhu S, Jainu M, Sabitha KE and Devi CS:
Role of mangiferin on biochemical alterations and antioxidant
status in isoproterenol-induced myocardial infarction in rats. J
Ethnopharmacol. 107:126–133. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Boyce M, Bryant KF, Jousse C, et al: A
selective inhibitor of eIF2alpha dephosphorylation protects cells
from ER stress. Science. 307:935–939. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Komoike Y, Inamura H and Matsuoka M:
Effects of salubrinal on cadmium-induced apoptosis in HK-2 human
renal proximal tubular cells. Arch Toxicol. 86:37–44. 2012.
View Article : Google Scholar
|
|
34
|
George I, Sabbah HN, Xu K, Wang N and Wang
J: β-Adrenergic receptor blockade reduces endoplasmic reticulum
stress and normalizes calcium handling in a coronary embolization
model of heart failure in canines. Cardiovasc Res. 91:447–455.
2011. View Article : Google Scholar : PubMed/NCBI
|