Introduction
Klebsiella pneumoniae is a species of aerobic
gram-negative bacteria, which primarily inhabits the intestines,
respiratory system and urinary system of humans and animals
(1). It is well known as a cause
of community-acquired pneumonia (2). Resistance of K. pneumoniae to
antibiotics is an increasingly severe problem resulting in an
urgency for the development of novel types of antibiotic (3).
Invasion of host cells is the singular most
important mechanism that bacterial pathogens employ to avoid rapid
clearance by the host immune system (4). A common and recurring target of
pathogens is the cytoskeleton of host cells, which is utilized by
these microbes for purposes that include attachment, entry into
cells, movement within and between cells, vacuole formation and
remodelling, and avoidance of phagocytosis (5). Actin has two forms, one is a monomer
(G-actin) and the other a multimer (F-actin). Normally, the
progress of polymerization into F-actin (6) and depolymerization into G-actin is
balanced to regulate actin activity, as only polymeric actin has
biological effects (7,8). Diverse bacteria invade non-phagocytic
cells by stimulating the endogenous uptake processes, including
phagocytosis and macropinocytosis. Actin polymerization is central
to these two processes, driving plasma membrane extensions that
engulf external cargo (9).
Cytokines and chemokines are critical molecules in
response to pathogen invasion and are involved in almost every
facet of immunity and inflammation (10,11).
Interleukin (IL)-6 has been identified to enhance survival of
pneumonia sepsis caused by K. pneumoniae via increasing
neutrophil death (12). CXCL1 has
been demonstrated to be able to enhance resistance to K.
pneumonia in mice (13,14).
LCN2 may inhibit bacterial growth by sequestering the iron-binding
bacterial siderophores and blocking bacterial access to iron
(15,16).
Anti-microbial peptides (AMPs) are active at
physiological concentrations under conditions prevailing in the
tissues of origin (17).
High-mobility group nucleosome-binding domain 2 (HMGN2) protein, a
member of the HMGN family, binds specifically to nucleosomes
(18) in the cell nucleus of
vertebrate and invertebrate organisms and has a role in global DNA
repair (19). Previous studies by
our group revealed that HMGN2 protein had anti-bacterial activity
and is therefore considered as a potent AMP (20,21).
Is was able to inhibit the gene expression and replication of
hepatic B virus in vitro (22) and also exhibited anti-cancer
potency in HeLa cells (23).
Further studies by our group demonstrated that pre-treatment of
K. pneumoniae 03183 with 128 μg/ml HMGN2
significantly inhibited the internalization of K. pneumoniae
03183 into human bladder carcinoma T24 cells by decreasing K.
pneumoniae-induced actin polymerization (24,25).
HMGB1, a member of the HMG superfamily, stimulates
pro-inflammatory cytokine synthesis in human monocytes (26) and contributes to the occurrence of
rheumatoid arthritis, systemic lupus erythematosus and polymyositis
(27,28). HMGN1, like HMGN2, a member of the
HMGN subfamily, also induces pro-inflammatory cytokine production
in vitro and in vivo (29). In the present study, K.
pneumoniae 03183 bacteria were pre-treated with 128
μg/ml HMGN2 and the effects of HMGN2 on K. pneumoniae
03183-induced actin polymerization as well as on the expression of
the K. pneumoniae 03183-associated cytokines lipocalin-2
(LCN2), chemokine (C-X-C motif) ligand 1 (CXCL1) and IL-6 in mouse
lungs were assessed.
Materials and methods
Chemicals
Recombinant human HMGN2 was prepared as previously
described (24). 1% bovine serum
albumin (BSA) was obtained from Thermo Fisher Scientific (Waltham,
MA, USA).
Bacteria and culture conditions
K. pneumoniae strain 03183 was isolated from
a sample from a patient with a urinary tract infection at the
Department of Medicine, West China Hospital affiliated to Sichuan
University (Chengdu, China). K. pneumoniae 03183 was grown
in Luria-Bertani (LB) broth (Oxoid Ltd., Basingstoke, UK) for 14–16
h at 37°C until reaching the middle of the logarithmic growth
phase. The concentration of bacteria in the LB broth was determined
by measuring the absorbance at 650 nm. Later, K. pneumoniae
03183 was pre-treated with 128 μg/ml HMGN2 and 1% BSA
separately for 2 h at 37°C. The inoculum was pelleted by
centrifugation at 7,000 x g for 2 min and resuspended in
phosphate-buffered saline (PBS; Chengdu Kelong Chemical Co., Ltd.,
Chengdu, China) to obtain the desired concentration (109
colony-forming units).
Animals and treatment
A total of 24 female C57BL/6 mice (6–8 weeks old
with similar weights) were obtained from the Experimental Animal
Center of Sichuan University (Chengdu, Sichuan). The mice were fed
a normal diet and were kept at room temperature with a cycle of 12
h light and 12 h dark. They were randomly divided into four groups:
In the PBS group, mice were transtracheally inoculated with PBS; in
the HMGN2 group, mice were transtracheally inoculated with K.
pneumoniae 03183 pre-treated with 128 μg/ml HMGN2; in
the BSA group, mice were transtracheally inoculated with K.
pneumoniae 03183 pre-treated with 1% BSA; in the K.
pneumoniae group, mice were transtracheally inoculated with
untreated K. pneumoniae 03183. Each experimental group
contained six animals. The study was approved by the ethics
committee of West China College of Preclinical Sciences and
Forensic Medicine, Sichuan University.
In vivo invasion assay
Each animal was anesthetized with by i.p. injection
of 10% chloral hydrate (Chengdu Kelong Chemical Co., Ltd.). The
trachea was exposed, and the respective inoculum was administered
via a sterile 26-gauge needle. The skin incision was closed by
surgical suture. After 2 h, the lung was aseptically removed, 0.1
mg of gentamicin was applied for 30 min to kill all extracellular
bacteria, the lung was washed briefly using 100 μl PBS and
then homogenized in PBS according to a previously published
procedure (4). Homogenate
dilutions were plated at ~1×105 cells/ml on LB plates
and incubated overnight at 37°C for colony counts.
Histopathological examination and
immunofluorescence of lung sections
Each animal was anesthetized and inoculated
transtracheally with inoculum or PBS. After 2 h, the lung was
aseptically removed, immersed in 10% formalin fixative (Chengdu
Kelong Chemical Co., Ltd.) and processed for histological
examination. The lung tissue was embedded in paraffin wax (Chengdu
Kelong Chemical Co., Ltd.) and cut into 4–6 mm-thick sections using
a microtome. After this, the sections were stained with hematoxylin
and eosin (Beyotime Institute of Biotechnology, Nanjing, China) by
standard techniques (30). To
observe the expression of F-actin by immunohistochemistry, sections
of uninfected and infected lungs were prepared using standard
methods (31). Each section was
immerged in 30% H2O2 (Chengdu Kelong Chemical
Co., Ltd.)/distilled water (1:10) for 10 min at room temperature.
Each section was blocked in PBS containing 5% BSA for 10 min at
room temperature following microwave antigen retrieval, which
involved heating the sections in citrate buffer at 92–98°C for
10–15 min. The sections were then incubated with monoclonal
anti-mouse F-actin (1:100; ab130935; Abcam, Cambridge, UK) diluted
1:100 in PBS with 5% BSA overnight at 4°C. Sections were incubated
for 30 min at 37°C with biotinylated mouse anti-mouse
immunoglobulin IgG (1:100, ZB-2055; ZSGB-BIO, Beijing, China), then
streptavidin-horseradish peroxidase (HRP) (1:100; ZB2404; HongYue
ChuangXin Technology Co., Ltd., Bejing, China) was applied for 30
min at 37°C, using a diaminobenzidine kit (ZSGB-BIO) to display the
results. For immunostaining, sections were fixed and blocked in the
same manner as stated above and incubated with monoclonal mouse
F-actin antibody diluted 1:50 in PBS with 5% BSA overnight at 4°C.
Sections were then incubated with monoclonal biotinylated Cy3 goat
anti-mouse IgG (1:50; C5585; Sigma-Aldrich) for 60 min at 37°C and
DAPI (for nuclear staining; Sigma-Aldrich, St Louis, MO, USA) for
30 min at room temperature in the dark, mounted and viewed using an
Axio Scope A1 microscope (Zeiss, Oberkochen, Germany).
Quantification of F-actin by western blot
analysis
To examine F-actin protein by western blot analysis,
lung tissues were homogenized with lysis buffer (10 mM Tris, pH
8.0; 1 mM EDTA; 400 mM NaCl; 10% glycerol; 0.5% NP-40; 5 mM sodium
fluoride; 0.1 mM phenylmethylsulfonylfluoride; and 1 mM
dithiothreitol) (32,33). The lysates were centrifuged (12,000
× g, 20 min, 4°C) and the supernatants were collected. Protein
concentrations were determined using a BCA-1 bicinchoninic acid
protein assay kit (Sigma-Aldrich), and equal amounts of protein (50
μg) were separated on precast 10% SDS-polyacrylamide gels
(Beyotime Institute of Biotechnology), electrotransferred to
polyvinylidene fluoride membranes (Merck Millipore, Darmstadt,
Germany) and blocked with 5% skimmed milk (Sigma-Aldrich) at room
temperature for 1 h. The membranes were incubated with primary
antibody for F-actin (1:500) or GAPDH (1:500; AG019; Beyotime
Institute of Biotechnology, Haimen, China) at 4°C overnight.
Following repeated washing, the membranes were incubated with
HRP-conjugated anti-mouse secondary antibody (1:5,000; ZB-2305;
ZSGB-BIO). The expression of GAPDH was detected as an internal
control and the relative content of target proteins was analyzed
using an enhanced chemiluminescence system (BeyoECL Plus; Beyotime
Institute of Biotechnology, Nanjing, China).
RNA isolation and reverse transcription
quantitative polymerase chain reaction (RT-qPCR) amplification of
whole lung mRNA
To extract RNA from the lungs, 100 mg frozen lung
tissue was homogenized with 1 ml TRIzol (Ambion, Life Technologies,
Carlsbad, CA, USA) in a ribonuclease-free tube at 4°C. RNA
isolation was performed according to the manufacturer’s
instructions. Following extraction, total RNA was subjected to
reverse transcription by using a First Strand cDNA Synthesis kit
from Thermo Fisher Scientific, according to the manufacturer’s
instructions. The final product complementary DNA (cDNA) was stored
at −20°C. cDNA was amplified on a Bio-Rad CFX-96 real-time system
using SYBR Green qPCR Master mix from Thermo Fisher Scientific. The
following specific primers were used: LCN2,
5′-CCAGTTCGCCATGGTATTTTTC-3′ (sense) and
5′-CACACTCACCACCCATTCAGTT-3′ (anti-sense); CXCL1,
5′-GCTGGGATTCACCTCAAGAA-3′ (sense) and 5′-TCTCCGTTACTTGGGGACAC-3′
(anti-sense), IL-6, 5′-GAGGATACCACTCCCAACAGACC-3′ (sense) and
5′-AAGTGCATCATCGTTGTTCATACA-3′ (anti-sense), β-actin,
5′-CGGTTCCGATGCCCTGAGGCTCTT-3′ (sense) and
5′-CGTCACACTTCATGATGGAATTGA-3′ (anti-sense); all were purchased
from Sangon Biotech (Shanghai, China). Relative expression of GAPDH
was normalized to the β-actin levels.
Statistical analysis
Values are expressed as the mean ± standard
deviation. Statistical analysis was performed using SPSS 13.0
(SPSS, Inc., Chicago, IL, USA). Statistical analyses were performed
using the Student’s t-test. P<0.05 was considered to indicate a
significant difference between values.
Results
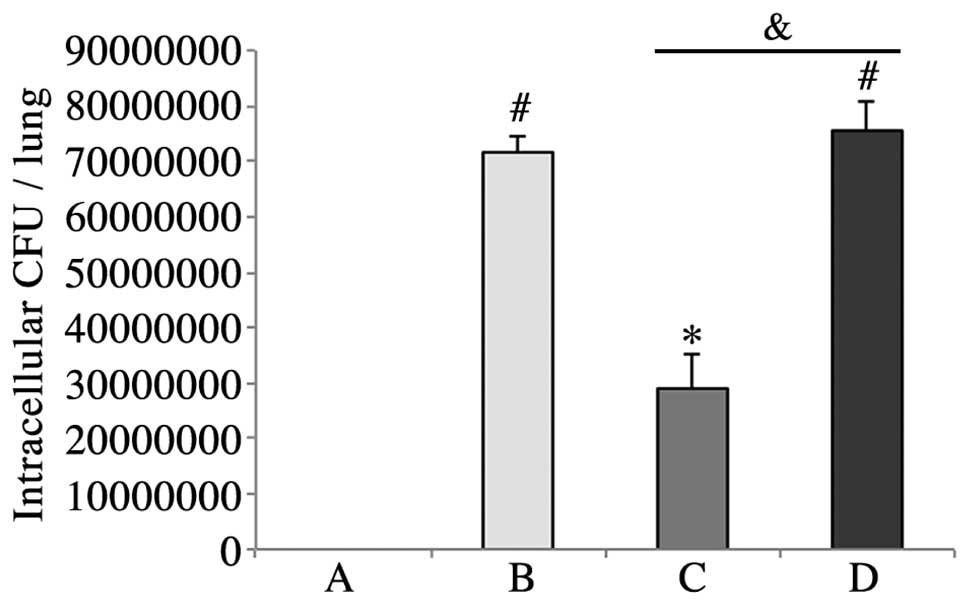
HMGN2 reduces K. pneumoniae 03183
invasion
128 μg/ml HMGN2 significantly inhibited the
invasion of K. pneumoniae 03183 into lungs. 1% BSA had no
effect on the invasion of K. pneumoniae 03183 into lungs as
compared with untreated K. pneumoniae 03183 (Fig. 1).
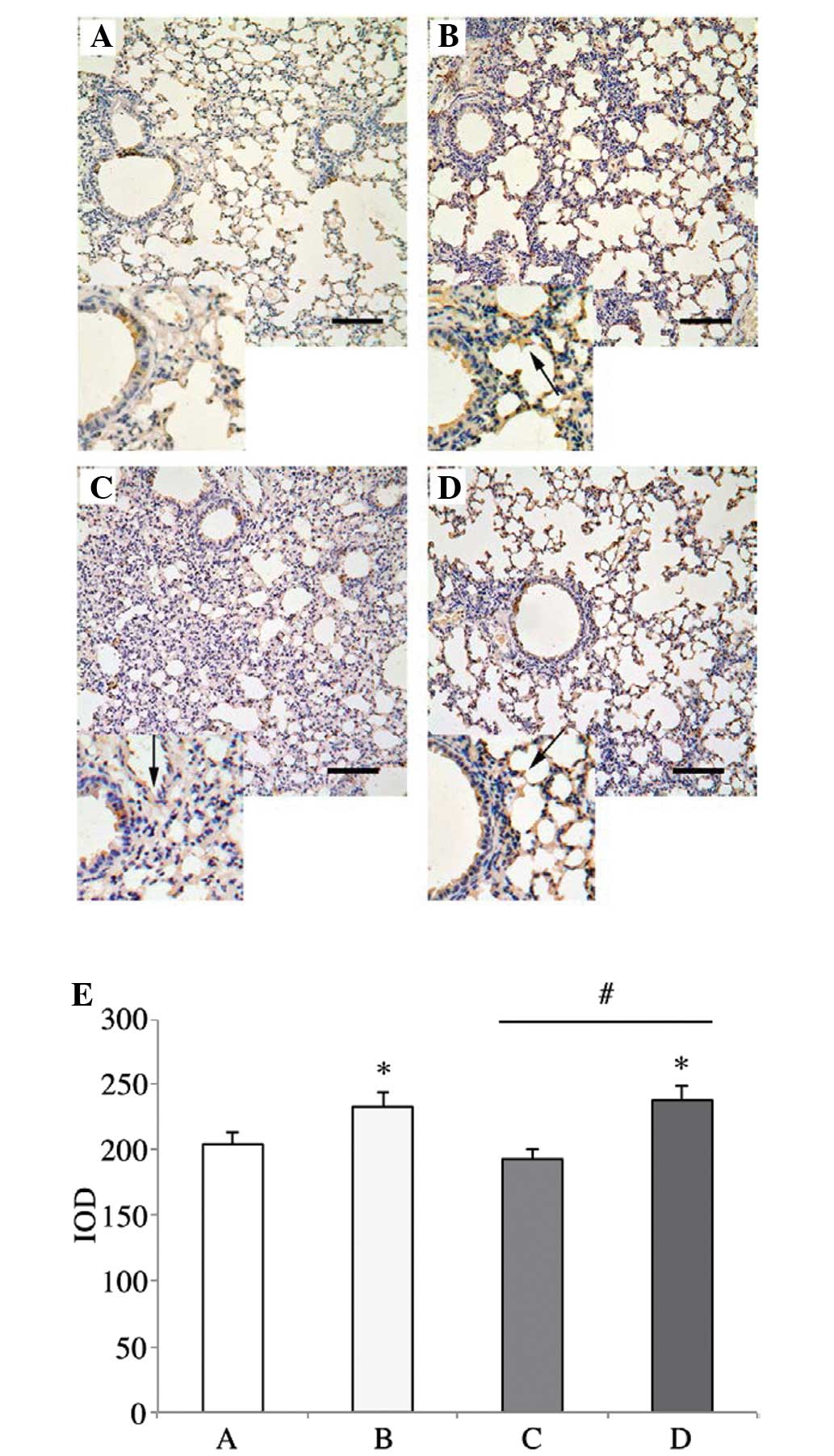
Pulmonary structure following K.
pneumoniae 03183-inoculation
As illustrated in Fig.
2A, the pulmonary structure in the mice of the PBS group was
normal. Although the alveoli of mice in the BSA group were
remaining, the pulmonary structure exhibited dilatation of alveolar
ducts, hyperemia and edema of alveolar walls, proliferation of
uncharacteristic cells but no evidence of inflammatory cells
(Fig. 2B). The condition of the
pulmonary structure in the HMGN2 group was in a worse condition
than that in the BSA group and exhibited excessive capillary
leakage, accumulation of white blood cells in the alveolar spaces
with diffuse damage of alveolar epithelial and endothelial cells
(Fig. 2C). Lungs of mice in the
K. pneumoniae group were similar to those in the BSA group;
however, normal alveoli were not present (Fig. 2D).
Expression of F-actin in lung tissue
Positive staining for F-actin expression was
significantly increased in the mice in the BSA and K.
pneumoniae groups, as compared with that in the mice in the
HMGN2 group (Fig. 3).
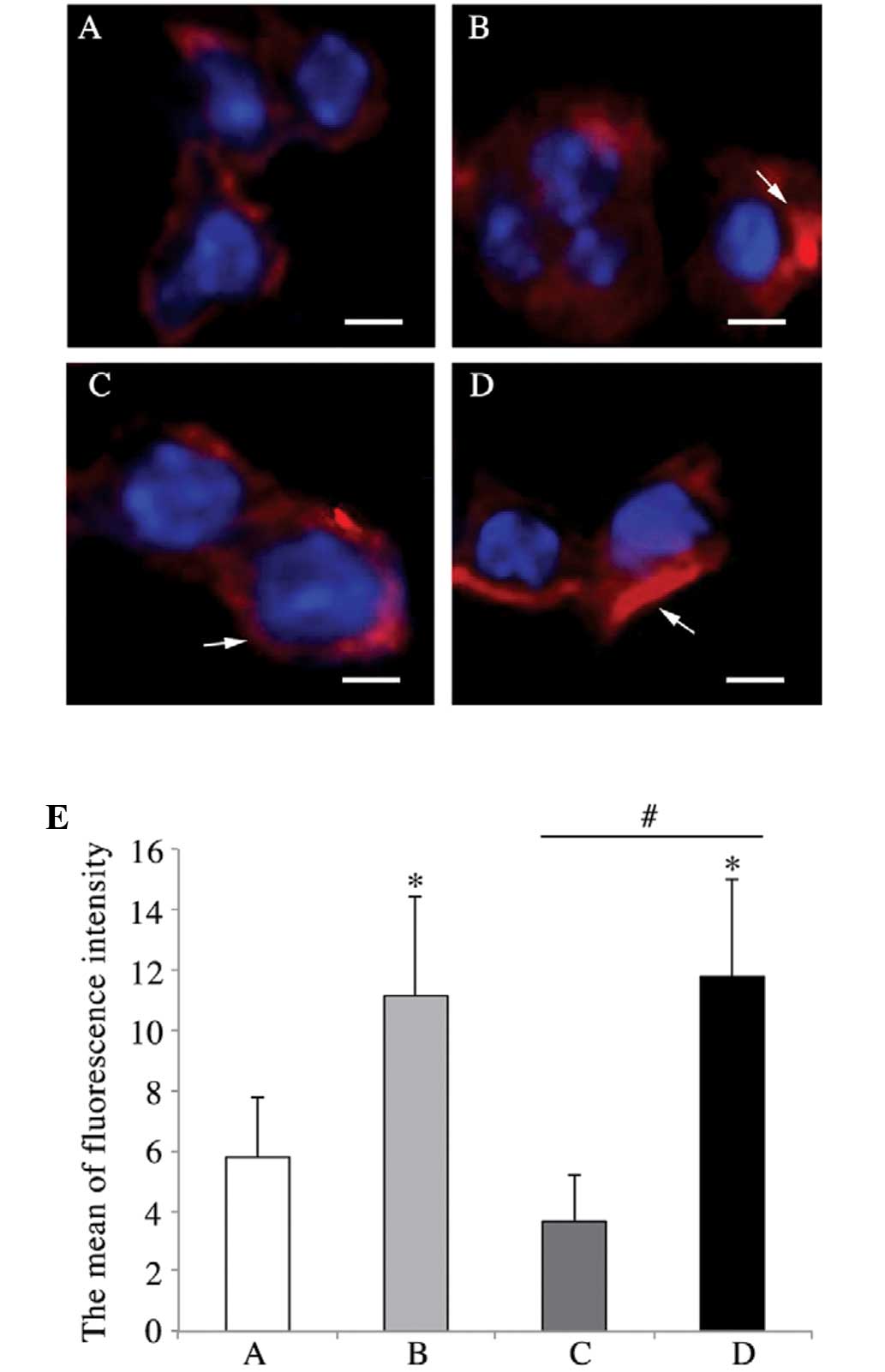
HMGN2 reduces K. pneumoniae-induced actin
fiber polymerization in pulmonary cells
As shown in Fig.
4A, in the pulmonary cells of mice in the PBS group,
well-organized actin filaments were present. Actin stress fibers
were observed at the cell periphery in most of the cells infected
with K. pneumoniae 03183 pre-treated with 1% BSA (Fig. 4B). By contrast, pre-treatment with
HMGN2 appeared to have eliminated the stress fibers and decreased
the overall content of F-actin; most of the actin filaments were
fragmented (Fig. 4C). Pulmonary
cells of mice inoculated with untreated K. pneumoniae 03183
showed a similar F-actin fiber distribution to that in the BSA
group (Fig. 4D). F-actin was
quantified as the mean fluorescence intensity (Fig. 4E).
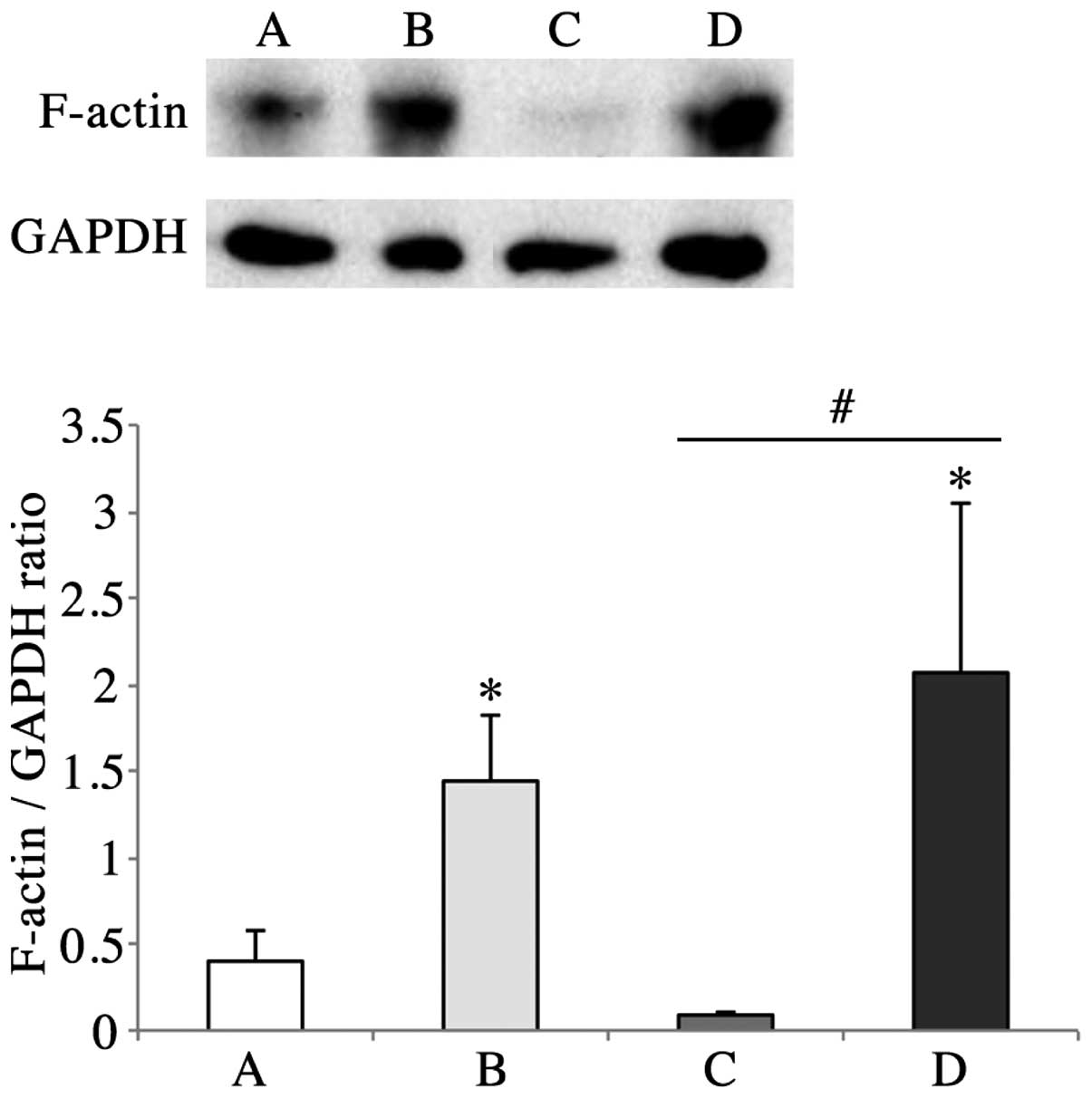
K. pneumoniae inoculation increases
F-actin protein levels in lung tissue
To illustrate the changes in F-actin expression at
the protein level, western blot analysis was employed to show the
quantity of F-actin in the four different groups. Consistent with
the results acquired by immunohistochemistry and
immunofluorescence, F-actin levels in the BSA and K.
pneumoniae groups were higher than those in the PBS group
(Fig. 5). However, F-actin levels
in the HMGN2 group were lower than those in the PBS group, which
may indicate that HMGN2 decreased the quantity of F-actin generated
by G-actin polymerization.
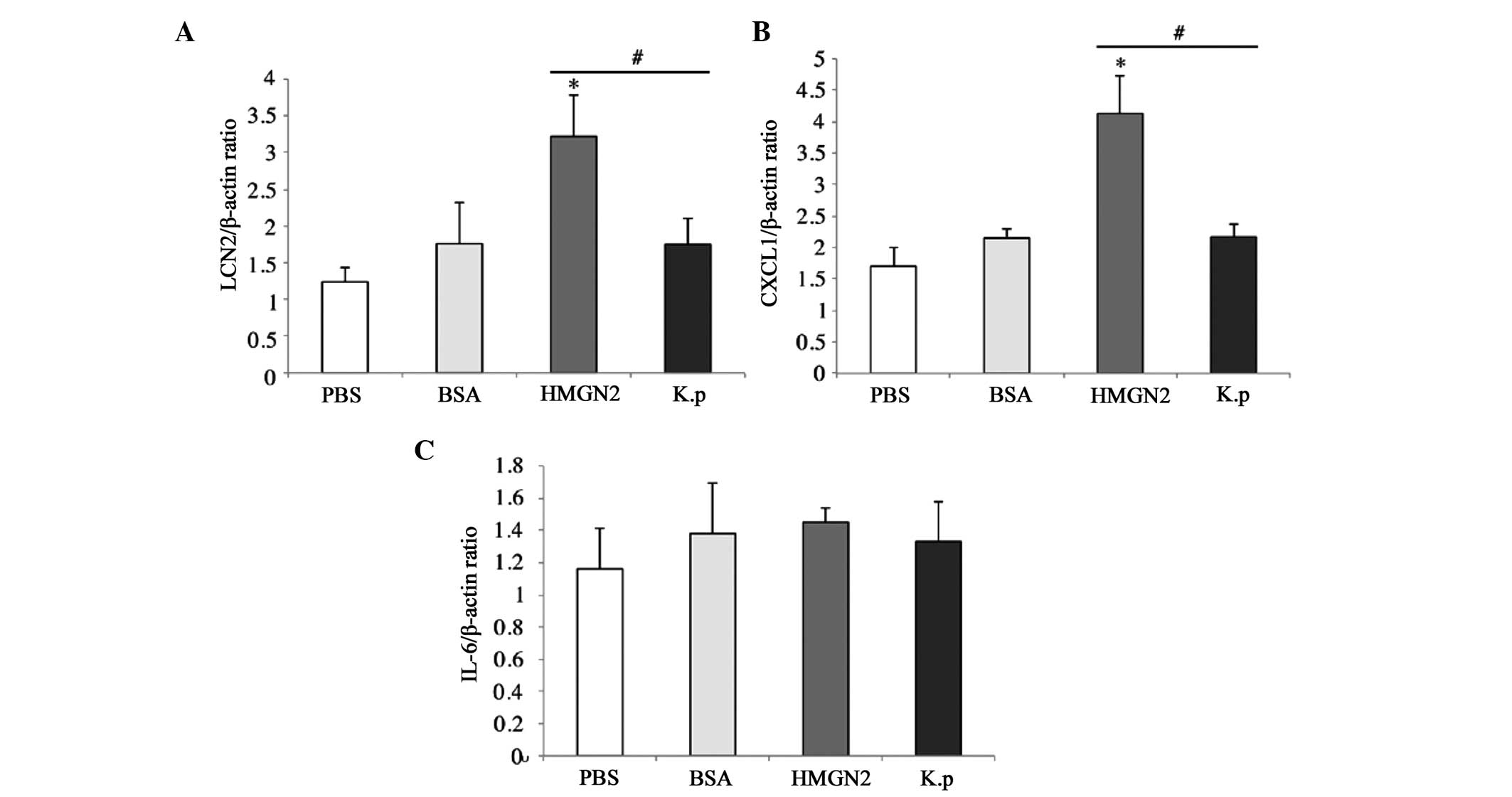
HMGN2 affects K. pneumoniae 03183-induced
inflammation-associated cytokines
To examine the effect of HMGN2 on K. pneumoniae
03183-induced inflammation-associated cytokine expression,
RT-qPCR was performed to measure mRNA levels of the cytokines LCN2,
CXCL1 and IL-6, which all have functions of restraining or killing
K. pneumoniae 03183 (Fig.
6) (12,13,15).
Compared with levels in the PBS group, LCN2 and CXCL1 mRNA levels
were higher in the BSA, HMGN2 and K. pneumoniae groups
(Fig. 6A and B). In the HMGN2
group, the mRNA levels of LCN2 and CXCL1 were more increased than
those in the BSA and K. pneumoniae groups, which indicated
that HMGN2 had an alleviating effect on K. pneumoniae
infection. In the BSA, K. pneumoniae and HMGN2 groups, IL-6
levels were higher than those in the PBS group; however, the
difference was not significant (Fig.
6C).
Discussion
The results of the present study indicated that
HMGN2 was able to reduce K. pneumoniae 03183 invasion of
host lung tissue by blocking K. pneumoniae 03183-induced
actin polymerization and up-regulating the secretion of the
inflammation-associated cytokines LCN2 and CXCL1.
Bacterial invasion to epithelial cells is a critical
step in the manifestation of bacterial infection, in addition to
evasion of the immune response of the organism and resistance to
antibiotics (34). In the present
study, bacteria adherent to epithelial cells were killed by the
bactericidal antibiotic gentamicin, which is unable to enter
eucaryotic cells, so that internalized organisms were protected
from its effects (35). The
results showed that HMGN2 significantly inhibited the invasion of
K. pneumoniae 03183 into lung tissue in vivo compared
with the invasion extent in the BSA and K. pneumoniae
groups. The effect HMGN2 was observed at a concentration of 128
μg/ml, which indicated that it inhibited the invasion of
K. pneumoniae 03183 neither by inhibiting the growth of
bacteria nor by inhibiting the physical progress of pre-treatment
with protein (24,25).
Altering the host cytoskeleton is crucial for
mediating pathogen adherence, invasion and intracellular
locomotion. Actin is the structure protein of the microfilament
cytoskeleton, existing in two forms, monomer (G-actin) and multimer
(F-actin). Normally, the progress of polymerization and
depolymerization of microfilaments is balanced for the purpose of
regulating actin activity, as only polymeric actin has biological
effects (7,8). In the present study,
immunohistochemical techniques, fluorescence microscopy and western
blot analysis were applied in order to observe the expression of
F-actin in lungs and to determine the extent of
depolymerization/polymerization of F-actin. The results of the
present study showed that the recomposition of the pulmonary
cytoskeleton was triggered by untreated K. pneumoniae 03183.
In the K. pneumoniae and BSA groups, F-actin expression and
stress fibers were increased and the average fluorescence intensity
of F-actin was significantly increased as compared with that in the
PBS group, which indicated that following infection, polymerized
F-actin was enhanced. Of note, in the HMGN2 group, F-actin
expression and average fluorescence intensity of F-actin were
decreased, which indicated that HMGN2 inhibited K. pneumoniae
03183 invasion of lung cells directly through reducing the
polymerization of F-actin, which is consistent with previous
studies by our group (24,25).
Cytokines and chemokines are critical molecules
expressed in response to the presence of invading pathogens
(10); they are involved in almost
every facet of immunity and inflammation (11). Inflammation is the first response
of the immune system to bacterial or viral infection (36). IL-6 has been found to enhance
survival of pneumonia sepsis caused by K. pneumoniae via
improving neutrophil killing (12). CXCL1, a member of the C-X-C
chemokine family, is broadly produced by numerous cells, including
fibroblasts, endothelial cells, as well as peritoneal and alveolar
MΦ (13). It was shown to be able
to enhance resistance to K. pneumonia in mice (13,14).
LCN2, produced by epithelial cells and macrophages, has been
identified as a potent bacteriostatic agent, inhibiting bacterial
growth by sequestering the iron-binding bacterial siderophores and
blocking bacterial access to iron (15,16).
In the present study, hematoxylin and eosin staining showed that
the lung structure in the HMGN2 group was in a worse condition than
that in the BSA and K. pneumoniae groups, which may indicate
that HMGN2 generates a brisk innate immune response to K.
pneumonia 03183 more rapidly. Furthermore, the results showed
the levels of LCN2 and CXCL1 mRNA in the HMGN2 group were higher
than those in the PBS, BSA, and K. pneumoniae groups, which
may indicate that pre-treatment of K. pneumoniae neumonia
with HMGN2 was able to improve the ability of the host to recognize
K. pneumoniae and to secrete associated cytokines. However,
no significant effect on IL-6 levels in the lung was noted.
In conclusion, HMGN2 inhibited the invasion of K.
pneumoniae 03183 into lungs through reducing the polymerization
of F-actin and increasing the levels of CXCL1 and LCN2 in
vivo, which indicated that HMGN2 may be an important defense
factor in the innate immune response.
Acknowledgments
China National Nature Science Fund 30671963 and
China Medical Board of New York INC98-681.
References
|
1
|
Jarvis WR and Martone WJ: Predominant
pathogens in hospital infections. J Antimicrob Chemother. 29:19–24.
1992. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Carpenter JL: Klebsiella pulmonary
infections: occurrence at one medical center and review. Rev Infect
Dis. 12:672–682. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Podschun R and Ullmann U: Klebsiella spp.
as Nosocomial pathogens: Epidemiology, taxonomy, typing methods,
and pathogenicity factors. Clin Microbiol Rev. 11:589–603.
1998.PubMed/NCBI
|
|
4
|
Song JM, Bishop BL, Li GJ, Duncan MJ and
Abraham SN: TLR4 initiated and cAMP mediated abrogation of
bacterial invasion of the bladder. Cell Host Microbe. 1:287–298.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Gruenheid S and Finlay BB: Microbial
pathogenesis and cytoskeletal function. Nature. 422:775–81. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Winder SJ and Ayscough KR: Actin-binding
proteins. J Cell Sci. 118:651–654. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tsai KW, Lai HT, Tsai TC, et al:
Difference in the regulation of IL-8 expression induced by
uropathogenic E. coli between two kinds of urinary tract epithelial
cells. J Biomed Sci. 16:912009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bhavsar AP, Guttman JA and Finlay BB:
Manipulation of host-cell pathways by bacterial pathogens. Nature.
449:827–834. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Haglund CM and Welch MD: Pathogens and
polymers: microbe-host interactions illuminate the cytoskeleton. J
Cell Biol. 195:7–17. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ye P, Garvey PB, Zhang P, et al:
Interleukin-17 and lung host defense against Klebsiella pneumoniae
infection. Am J Respir Cell Mol Biol. 25:335–340. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Borish LC and Steinke JW: Cytokines and
chemokines. J Allergy Clin Immunol. 111(2 Suppl): S460–S475. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sutherland RE, Olsen JS, McKinstry A, et
al: Mast cell IL-6 improves survival from Klebsiella pneumonia and
sepsis by enhancing neutrophil killing. J Immunol. 181:5598–5605.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tsai WC, Strieter RM, Wilkowski JM,
Bucknell KA, Burdick MD, Lira SA and Standiford TJ: Lung-specific
transgenic expression of KC enhances resistance to Klebsiella
pneumoniae in mice. J Immunol. 161:2435–2440. 1998.PubMed/NCBI
|
|
14
|
Cai S, Batra S, Lira SA, Kolls JK and
Jeyaseelan S: CXCL1 regulates pulmonary host defense to Klebsiella
infection via CXCL2, CXCL5, NF-kappaB, and MAPKs. J Immunol.
185:6214–6225. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Goetz DH, Holmes MA, Borregaard N, et al:
The neutrophil lipocalin NGAL is a bacteriostatic agent that
interferes with siderophore-mediated iron acquisition. Mol Cell.
10:1033–1043. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Flo TH, Smith KD, Sato S, et al:
Lipocalin2 mediates an innate immune response to bacterial
infection by sequestrating iron. Nature. 432:917–921. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ganz T: Defensins: antimicrobial peptides
of innate immunity. Nat Rev Immunol. 3:710–720. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Shirakawa H, Herrera JE, Bustin M and
Postnikov Y: Targeting of high mobility group-14/-17 proteins in
chromatin is independent of DNA sequence. J Biol Chem.
275:37937–37944. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Subramanian M, Gonzalez RW, Patil H, et
al: The nucleosome-binding protein HMGN2 modulates global genome
repair. FEBS J. 276:6646–6657. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Feng Y, Huang N, Wu Q and Wang B: HMGN2: a
novel antimicrobial effector molecule of human mononuclear
leukocytes? J Leukocyte Biol. 78:1136–1141. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Feng Y, Huang N, Wu Q, Bao L and Wang BY:
α-helical domain is essential for antimicrobial activity of high
mobility group nucleosomal binding domain 2 (HMGN2). Acta Pharmacol
Sin. 26:1087–1092. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Feng Y, He F, Zhang P, et al: Inhibitory
effect of HMGN2 protein on human hepatitis B virus expression and
replication in the HepG2.2.15 cell line. Antiviral Res. 81:277–282.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Xiong WB, Hung N, Feng Y, Wu Q and Wang
BY: Creation and anti-cancer potency in HeLa cells of a novel
chimeric toxin, HMGNCIDIN, composed of HMGN2 α-helical domain and
PE38 KDEL domain III. Chin Med J (Engl). 121:82–85. 2008.
|
|
24
|
Wu G, Cao Y, Fan B, et al: High-mobility
group protein N2 (HMGN2) inhibited the internalization of
Klebsiella pneumoniae into cultured bladder epithelial cells. Acta
Biochim Biophys Sin (Shanghai). 43:680–687. 2011. View Article : Google Scholar
|
|
25
|
Cao Y, Wu GX, Fan B, et al: High mobility
group nucleosomal binding domain 2 protein protects bladder
epithelial cells from Klebsiella pneumoniae invasion. Biol Pharm
Bull. 34:1065–1071. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Andersson U, Wang H, Palmblad K, et al:
High mobility group 1 protein (HMG-1) stimulates proinflammatory
cytokine synthesis in human monocytes. J Exp Med. 192:565–570.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ulloa L, Batliwalla FM, Andersson U, et
al: High mobility group box chromosomal protein 1 as a nuclear
protein, cytokine, and potential therapeutic target in arthritis.
Arthritis Rheum. 48:876–881. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Pisetsky DS, Erlandsson-Harris H and
Andersson U: High-mobility group box protein 1 (HMGB1): an alarmin
mediating the pathogenesis of rheumatic disease. Arthritis Res
Ther. 10:2092008. View
Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yang D, Postnikov YV, Li Y, et al:
High-mobility group nucleosome-binding protein 1 acts as an alarmin
and is critical for lipopolysaccharideinduced immune responses. J
Exp Med. 209:157–171. 2012. View Article : Google Scholar :
|
|
30
|
Mills B, Arrington JB and Sobin LH:
Laboratory methods in histotechnology. Washington, DC: American
registry of pathology; pp. 132–214. 1992
|
|
31
|
Deng LX, Wu GX, Cao Y, et al: The
chromosomal protein HMGN2 mediates the LPS-induced expression of
β-defensins in mice. Inflammation. 35:456–473. 2012. View Article : Google Scholar
|
|
32
|
Li QQ, Chen ZQ, Cao XX, et al: Involvement
of NF-κB/miR-448 regulatory feedback loop in chemotherapy-induced
epithelial-mesenchymal transition of breast cancer cells. Cell
Death Differ. 18:16–25. 2011. View Article : Google Scholar :
|
|
33
|
Nohara A, Okada S, Ohshima K, et al:
Cyclin-dependent kinase-5 is a key molecule in tumor necrosis
factor-α-induced insulin resistance. J Biol Chem. 286:33457–33465.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Lafont F and van der Goot FG: Bacterial
invasion via lipid rafts. Cell Microbiol. 7:613–620. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kihlström E and Andåker L: Inability of
gentamicin and fosfomycin to eliminate intracellular
Enterobacteriaceae. J Antimicrob Chemother. 15:723–728. 1985.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Snelgrove R, Gwyer E and Hussell T:
Modulation of immunity to respiratory viral infection. Future
Virol. 1:471–481. 2006. View Article : Google Scholar
|