Introduction
Pancreatic cancer is a significant health problem
worldwide (1), with an estimated
279,000 cases diagnosed globally in 2008 (2). The majority of patients are diagnosed
at an advanced stage, and the 5-year survival rate of patients
following surgery is poor, with 213,000 pancreatic
cancer-associated mortalities worldwide despite advancements in
surgery, radiation therapy and chemotherapy (3). Therefore, it is important to identify
the molecular mechanisms underlying the development and progression
of pancreatic cancer in order to develop novel strategies for its
early detection and treatment.
The present study investigated
serine/threonine-protein kinase-1 (SMG-1), which belongs to the
phosphatidylinositol 3-kinase-related kinase (PIKK) family
(4,5). Functionally, SMG-1 has been linked to
the nonsense-mediated decay (NMD) of mRNA, as a member of the mRNA
surveillance complex (4). SMG-1
also has NMD-independent functions (4), including cellular stress responses.
Previous studies (6,7) have demonstrated that hSMG-1 is an
important regulator of the cell cycle checkpoints by regulating the
synthesis and proteolysis of p21. SMG-1 is also considered to be a
potential cancer susceptibility gene. The catalogue of somatic
mutations in the COSMIC cancer database reveals that mutations in
SMG-1 are associated with breast, kidney and stomach cancer
(www.sanger.ac.uk/genetics/CGP/cosmic)
(8). Previous investigations have
demonstrated that SMG-1 is widely expressed in multiple tissues and
cell lines (5,9). It has been observed that SMG-1
regulates the G1/S checkpoint of the cell cycle via a p53-dependent
and a p53-independent pathway, and depletion of the SMG-1 protein
increases the cell growth of colorectal cancer cells, indicating
that SMG-1 is a tumor suppressor gene (10). Another study demonstrated that the
SMG-1-induced activation of the p53 pathway is associated with the
chemoprotective effects of tempol (11). However, additional studies have
revealed that the SMG-1 promoter hypermethylation-induced
downregulation of the expression of SMG-1 is associated with
improved survival rates in patients with human papaloma virus
(HPV)-positive head and neck squamous cell carcinoma (12). SMG-1 antagonizes tumor necrosis
factor-α-induced apoptosis in osteosarcoma cells (13). A kinome-wide screen identified
SMG-1 as an essential kinase for the survival of multiple myeloma
and SMG-1 knockdown with small interfering (si)RNA reduced the
survival of myeloma cell lines (14). Furthermore, SMG-1 mRNA has been
observed to be upregulated in acute myeloid leukemia (15). These studies indicated that SMG-1
may have different roles in cancer progression.
The present study detected the protein expression of
SMG-1 in pancreatic cancer tissue specimens and subsequently
assessed the effects of SMG-1 knockdown on the sensitivity of
pancreatic cancer cells to chemotherapy in vitro.
Materials and methods
Immunohistochemical analysis of the
expression of SMG-1 in pancreatic cancer tissue
The present study was approved by the ethics
committee of the Second Affiliated Hospital of Nanjing Medical
University (Nanjing, China). A pancreatic cancer tissue microarray
(TMA) was obtained from Alenabio (cat. no. PA2082; Xian, China).
The TMA contained 94 cases of pathologically diagnosed pancreatic
adenocarcinoma with certain additional clinicopathological data
from the patients. A pathologist inspected all the specimens and
confirmed the diagnosis of pancreatic adenocarcinoma, mucinous
adenocarcinoma, adenosquamous carcinoma, squamous cell carcinoma,
acinic cell carcinoma and neuroendocrine carcinoma, five cases of
normal pancreatic tissue and five cases of distant normal
pancreatic tissue. The age of the tissue donors ranged between 39
and 78 years, with a mean age of 57.5±9.9 years, and a female to
male ratio of 1.6. The tumor tissue was obtained from patients at
Tongxu People’s Hospital (Kaifeng, China).
For immunostaining of the SMG-1 protein, the TMA
sections were deparaffinized in xylene (Xilong Chemical Co., Ltd.,
Shenzhen, China) twice and rehydrated in a series of ethanol (100%
for 5 min, 95% for 5 min, 75% for 5 min) followed by ultrapure
water. Staining was performed by incubating the sections with mouse
monoclonal anti-SMG-1 antibody (1:300; cat. no. SAB1404950-100UG;
Sigma-Aldrich, St. Louis, MO, USA), at 4°C overnight. The intensity
of the SMG-1 staining was scored by a pathologist, in a blinded
manner, as negative (no signal), weak (weak intensity in <50% of
tumor cells), moderate (strong intensity in <40% of tumor cells)
or strong (strong intensity in the majority of tumor cells).
Cell lines and culture
The COLO-357, BxPc-3, Capan-1, Capan-2, SW1990 and
PANC-1 human pancreatic cancer cell lines were obtained from the
Shanghai Cell Bank (Shanghai, China) and cultured in Dulbecco’s
modified Eagle’s medium (DMEM), supplemented with 10% fetal bovine
serum (FBS), 100 μg/ml penicillin and 100 μg/ml
streptomycin in a humidified atmosphere of 5% CO2 at
37°C. All reagents were obtained from Wisent (St. Bruno, PQ,
Canada).
SMG-1 short hairpin (sh)RNA recombinant
lentiviral vectors and lentivirus
A lentiviral vector-mediated shRNA against SMG-1 was
designed and constructed by GenePharma (Shanghai, China). Four
pairs of shRNAs, each targeting different regions of the SMG-1
transcript (GenBank accession no. NM_015092), and one negative
control shRNA were constructed. The target mRNA sequences of SMG-1
were as follows: sh-SMG-1 #1 (no. 5252), 5′-GCAGAAAG
GTGGTTGACAATG-3′; sh-SMG-1 #2 (no. 6931), 5′-GCT
CGACACTATTCTGTAACA-3′; sh-SMG-1 #3 (no. 7512),
5′-GGGTGTAACTGGAGTAGAAGG-3′ and sh-SMG-1 #4 (no. 8877):
5′-GGAAGCGTCTGAGACAGTTCA-3′. The scrambled sequence,
5′-ACTACCGTTGTTATAGGTG-3′, was used as a negative control. These
lentiviral vectors were then used by GenePharma to produce the
lentivirus.
Lentivirus infection of pancreatic cancer
cell lines
To knock down the expression of SMG-1, the SW1990
and PANC-1 pancreatic cancer cell lines were infected with a
lentivirus. Briefly, the cells were seeded into six-well plates
(Corning Costar, Inc., Corning, NY, USA) at a density of 40% and
grown for 24 h at 37°C. The cells were then infected with a
lentivirus containing the shRNA targeting SMG-1[SW1990 at an
multiplicity of infection (MOI) of 10 and PANC-1 at an MOI of 15,
based on pre-experimental data], according to the manufacturer’s
instructions. The lentiviral infection efficiency was confirmed by
the immunofluorescence density of the enhanced green fluorescent
protein reporter gene, and the RNA interference (i) efficiency was
determined by analyzing the mRNA and protein expression of SMG-1.
Based on these investigations, SMG-1 shRNA lentivirus construct #1
was selected for the subsequent experiments (data not shown).
RNA isolation and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
The cells were washed with phosphate-buffered saline
(PBS; Beyotime Institute of Biotechnology, Haimen, China) and the
total RNA was isolated from the cells using RNAiso Plus (Takara
Bio, Inc., Dalian, China), according to the manufacturer’s
instructions. Primer-Script RT Master mix (Takara Bio, Inc.) was
used to synthesize cDNA from the RNA samples. qPCR was performed
using SYBR Green (Roche Diagnostics, Indianapolis, IN, USA) on a
7500 Real-Time-PCR System (Applied Biosystems, Foster City, CA,
USA). The qPCR amplification was performed on 10 ng cDNA (total
reaction volume, 20 μl) as follows: An initial cycle of 95°C
for 10 min, 40 cycles of 94°C for 30 sec, 60°C for 30 sec and 72°C
for 30 sec, and then a final extension at 72°C for 5 min. The mRNA
expression of β-actin was used as an internal control for
determining the relative mRNA expression of SMG-1. The cycle
threshold (Ct) comparative ΔΔCt method was used to calculate the
relative mRNA expression of SMG-1, and the fold-changes were
analyzed by 2−ΔΔCt (16). The primers used for RT-qPCR were as
follows: SMG-1, forward 5′-TTAATCGCCAAGAAACACCC-3′ and reverse
5′-AGGAATCTTGGGCCTTTTGT-3′ and β-actin, forward
5′-CTCCATCCTGGCCTCGCT-3′ and reverse 5′-GCTGTCACCTTCACCGTTCC-3′.
All the experiments were performed in triplicate and repeated three
times with independent RNA samples.
Protein extraction and western
blotting
The total cellular protein was extracted from the
SW1990 and PANC-1 cells using radioimmunoprecipitation buffer
(Beyotime Institute of Biotechnology), supplemented with 1%
phenylmethyl-sulfonylfluoride (Beyotime Institute of
Biotechnology). The protein concentration was estimated with a
(bicinchoninic acid) BCA kit (Nanjing KeyGen Biotech. Co., Ltd.,
Nanjing, China). Following quantification, the protein samples were
separated by 6% sodium dodecyl sulfate polyacrylamide gel
electrophoresis (Beyotime Institute of Biotechnology) and
transferred onto polyvinylidenedifluoride membranes (Beyotime
Institute of Biotechnology). The membranes were blocked with 5%
non-fat dry milk (Yili Industrial Group Co., Ltd., Inner Mongolia,
China) in Tris-buffered saline (TBS) and incubated with the
following primary antibodies: mouse monoclonal anti-SMG-1 (1:300;
cat. no. SAB1404950-100UG; Sigma-Aldrich) or mouse monoclonal
anti-GAPDH (1:500; cat. no. AG019; Beyotime Institute of
Biotechnology, Jiangsu, China) at 4°C overnight. The following day,
the membranes were washed with TBS-Tween-20 (TBS-T; Beyotime
Institute of Biotechnology) and further incubated with a secondary
horseradish peroxidase-coupled goat anti-mouse antibody (Santa Cruz
Biotechnology, Inc., Santa Cruz, CA, USA) for 2 h at room
temperature. The membranes were washed three times with TBS-T and
the color was developed using an electrochemiluminescence kit
(Pierce Biotechnology, Inc., Rockford, IL, USA). The membranes were
then exposed to X-ray film (GelDoc XR system; Bio-Rad Laboratories,
Inc., Hercules, CA, USA) to visualize the signals. The target
protein expression of GAPDH was used as an internal control for
determining the relative expression levels of SMG-1.
Cell proliferation assay
The cells were infected with either the negative
control lentivirus or the SMG-1 shRNA lentivirus for 3 days at 37°C
and then seeded into 96-well culture plates (Costar, Cambridge, UK)
at a density of 2×103 cells/well. The cells were
incubated at 37°C for up to 5 days. Cell proliferation was detected
daily using a Cell Counting kit-8 (CCK-8; Nanjing KeyGen Biotech.,
Co., Ltd.), according to the manufacturer’s instructions, for 5
days. Briefly, 10 μl CCK-8 solution was added to each well
and the optical density was detected using a microplate reader
(Sunrise™; Tecan, Grödig, Austria) at 450 nm, with a reference
wavelength of 650 nm. Each assay was performed in triplicate and
repeated independently three times.
Tumor cell migration and invasion
assay
The tumor cell migration and invasion capacities
were measured using a Transwell chamber assay with or without
Matrigel coating. The Transwell chambers were 6.5 mm in diameter
and had a 8 μm pore size (Corning Costar, Inc.). The SW1990
and PANC-1 cells were seeded into the upper chamber
(5.0×104 cells per Transwell) pre-coated with or without
1 mg/ml Matrigel (BD Biosciences, San Jose, CA, USA), and the lower
wells were filled with 500 μl 10% FBS-DMEM. Following
incubation for 24 h at 37°C, the non-invading cells were removed
using cotton swabs and the cells that had invaded into the
underside of the membrane were stained with 0.1% crystal violet
(Beyotime Institute of Biotechnology) for 15 min at 37°C. The
membranes were washed with PBS and the invading cells were counted
under an inverted microscope (Eclipse Ti-E; Nikon Corporation,
Tokyo, Japan). All the experiments were performed in triplicate and
repeated once.
Flow cytometric analysis of cell cycle
distribution and apoptosis
The cell cycle progression and apoptosis were
assessed by flow cytometry using a BD FACSCalibur (BD Biosciences).
The SW1990 and PANC-1 cells were grown and infected with sh-SMG-1
or with a control lentivirus at 37°C for 12 h, and then treated
with 10 mg/ml gemcitabine or cisplatin (Jiangsu Hansoh
Pharmaceutical Co., Ltd., Jiangsu, China). Cell cycle analysis was
conducted using PI/RNase Staining Buffer (BD Pharmingen, San Diego,
CA, USA). For cell cycle analysis, the cells were collected, washed
twice with PBS and fixed with 70% ethanol at −20°C overnight. The
cells were then washed twice with PBS and resuspended in 500
μl PBS containing 0.2% Triton-X-100, 10 mM EDTA, 100
μg/ml RNase A and 50 μg/ml propidium iodide. The
samples were incubated at room temperature for 30 min.
Apoptosis analysis was conducted using Annexin
V-FITC Apoptosis Detection kit (BD Pharmingen). For the detection
of apoptosis, the cells were collected and washed twice with PBS,
prior to suspending in 100 μl 1X binding buffer and staining
with 5 μl annexin-V 647 and 5 μl 7-aminoactinomycin D
at room temperature for 15 min in the dark. The samples were
analyzed using a flow cytometer (BD FACSCalibur; BD Biosciences).
All the experiments were performed in triplicate and repeated
once.
Statistical analysis
All data are expressed as the mean ± standard
deviation. Student’s t-test was used to analyze the differences
between groups using SPSS 16.0 software (SPSS, Inc., Chicago, IL,
USA). The differences among the strips from the western blot
analysis were inferred by comparing the gray level of the strips
using ImageJ software, version 1.43 (SeekBio, Huzhou, China).
P<0.05 was considered to indicate a statistically significant
difference.
Results
Differential protein expression of SMG-1
in pancreatic cancer tissues
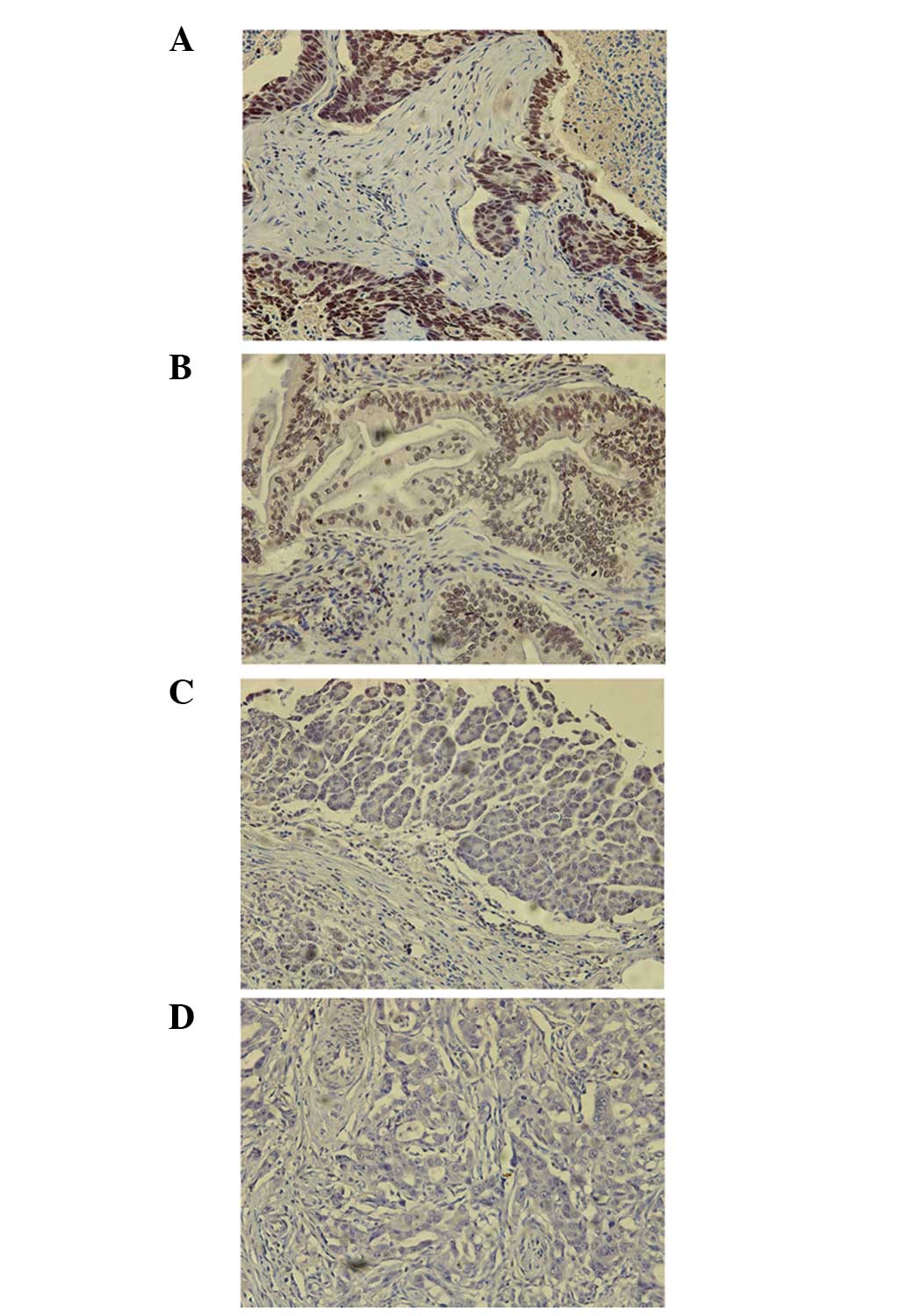
The present study first assessed the protein
expression levels of SMG-1 in normal and cancerous pancreatic
tissues. Immunohistochemical staining detected SMG-1 in the nuclei
and cytoplasm, but was more predominant in the nuclei of the tumor
cells (Fig. 1). According to the
expression of SMG-1, the pancreatic cancer cases were divided those
exhibiting high (moderate/strong) and low (negative/weak)
expression levels. In total, 51.1% (48/94) of the pancreatic cancer
tissues expressed a high level of SMG-1 protein, whereas 48.9%
(46/94) expressed a low level of SMG-1 protein (Table I). By contrast, 2/5 distant
pancreatic tissues and 1/5 normal pancreatic tissues expressed a
high level of SMG-1 protein. These results were consistent with
those in the Human Protein Atlas (http://www.protein-atlas.org/ENSG00000157106). Since
the TMA was obtained from a company with limited
clinicopathological data, the expression of SMG-1 was correlated
with the expression of SMG-1 among pancreatic cancer tissue,
distant pancreatic tissue and normal pancreatic tissue, which
revealed that the expression of SMG-1 was associated with an
advanced tumor stage, but not with age and gender (Table I).
 | Table IAssociation of the expression of
serine/threonine-protein kinase-1 with clinicopathological data
from the patients. |
Table I
Association of the expression of
serine/threonine-protein kinase-1 with clinicopathological data
from the patients.
| Group | Total (n) | Expression level of
SMG-1
|
|---|
| Low (n) | High (n) |
|---|
| Gender | | | |
| Female | 36 | 16 | 20 |
| Male | 58 | 30 | 28 |
| Age (years) | | | |
| 45 | 11 | 5 | 6 |
| 45–60 | 53 | 25 | 28 |
| 60 | 30 | 16 | 14 |
| Pathology type | | | |
| Ductal
adenocarcinoma | 83 | 43 | 40 |
| Others | 11 | 4 | 7 |
| TNM gradinga | | | |
| I | 31 | 21 | 10 |
| II | 45 | 18 | 27 |
| III | 7 | 3 | 4 |
| N/A | 11 | 4 | 7 |
Expression and knockdown of SMG-1 in
pancreatic cancer cell lines
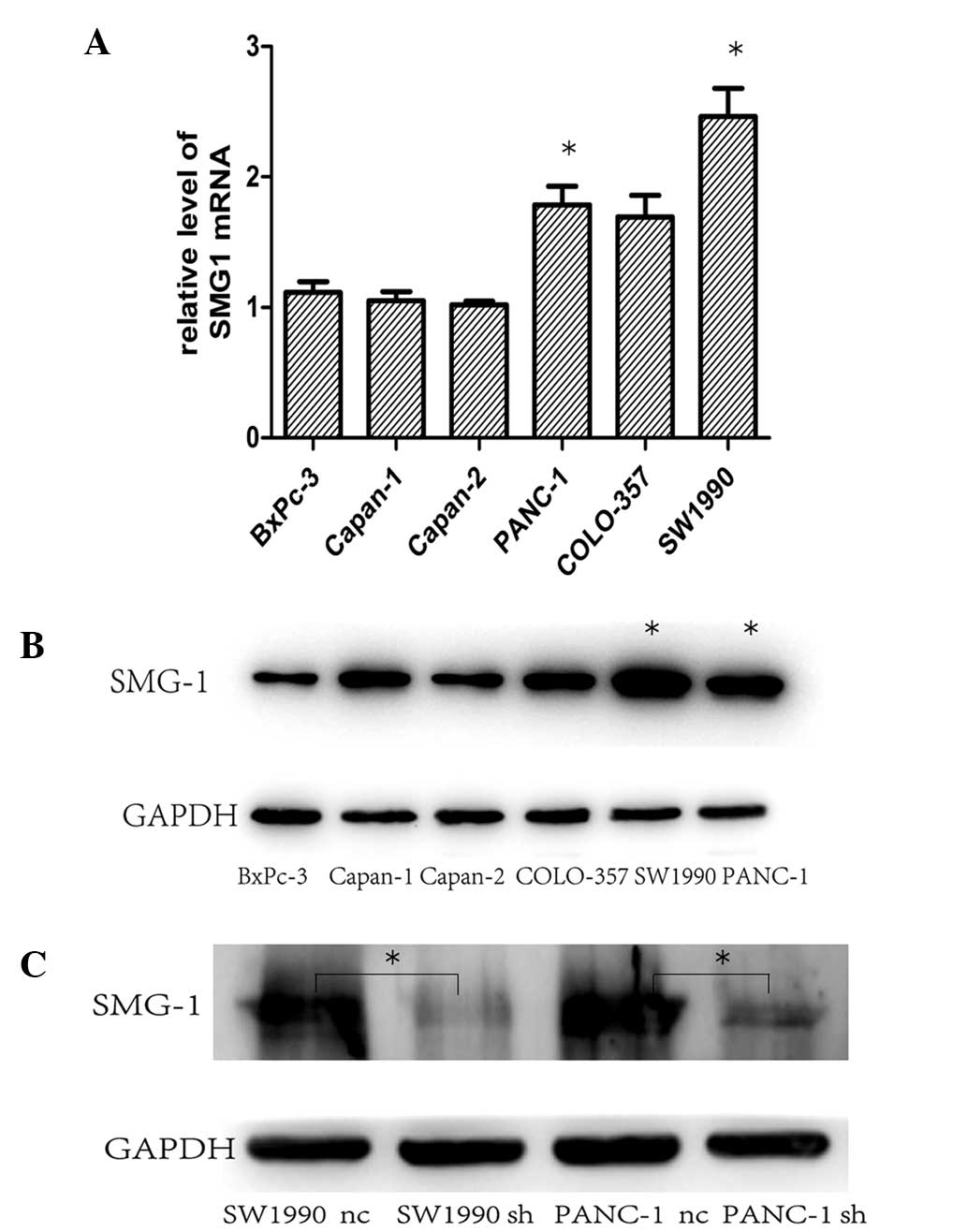
The expression levels of SMG-1 in six different
pancreatic cancer cell lines were assessed by western blot
analysis. The data revealed that all six pancreatic cancer cell
lines expressed high protein levels of SMG-1, and these levels were
highest in the PANC-1 and SW1990 cells (Fig. 2A).
Thus, PANC-1 and SW1990 cells were selected for the
subsequent knock down of SMG-1 expression using the shRNA
lentivirus. The data revealed that SMG-1 shRNA lentivirus #1
significantly reduced the protein expression levels of SMG-1 in the
twi cell lines (Fig. 2B) compared
with the negative control shRNA lentivirus.
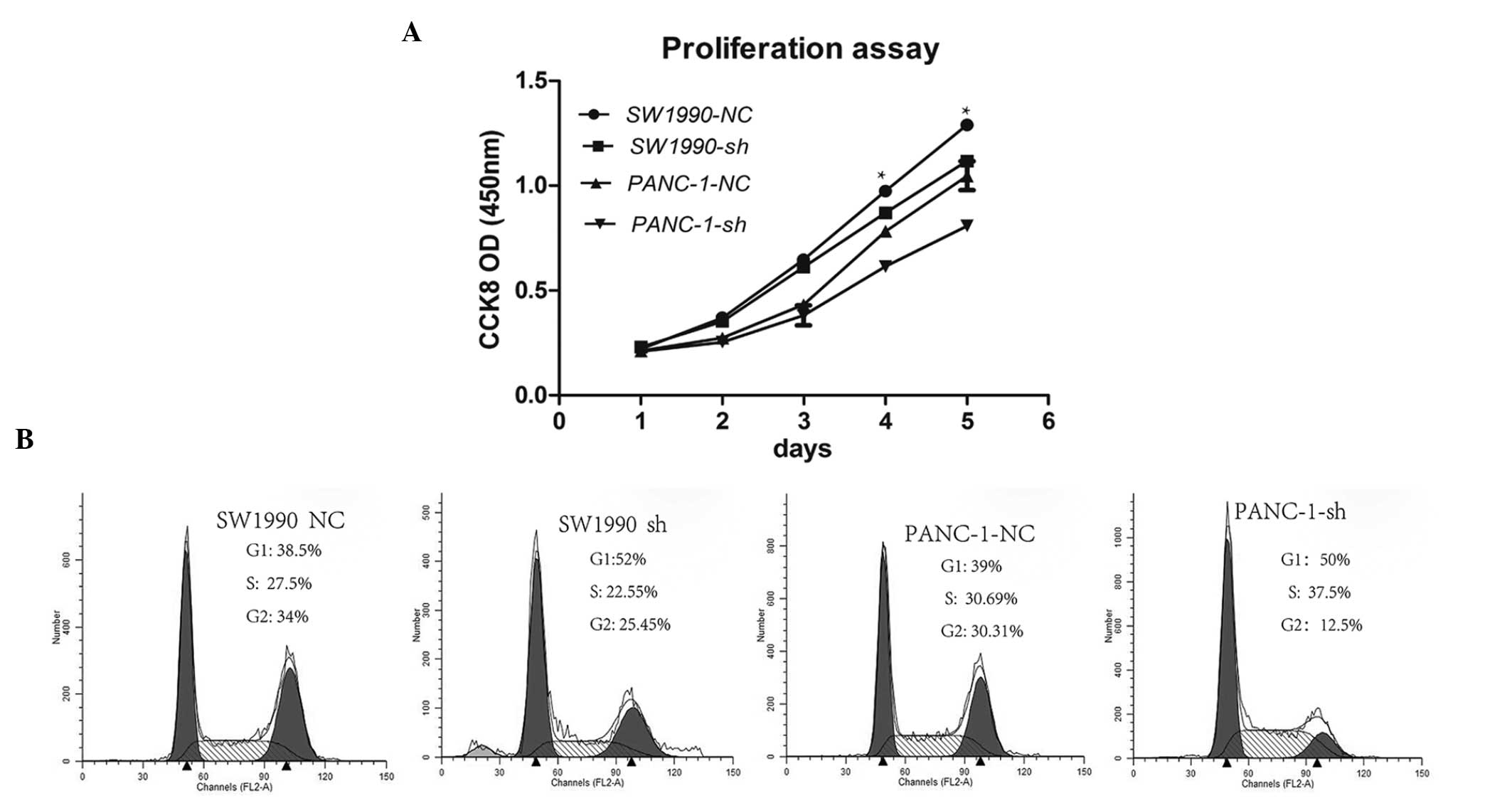
Knockdown of SMG-1 inhibits the
proliferation of pancreatic cancer cells
As shown in Fig.
3A, knockdown of SMG-1 significantly inhibited the
proliferation of PANC-1 and SW1990 cells compared with negative
control lentivirus-infected tumor cells (P<0.05, vs. control).
Flow cytometric analysis revealed that the knockdown of SMG-1
reduced the S/G2 phase of the cell cycle compared with the control
cells in the SW1990 cells (61.67±2.25, vs. 44.3±3.5%, P<0.05)
and PANC-1 cells (61±4, vs. 49.7±3.5%, P<0.05; Fig. 3B), while the G1 phase of the cell
cycle was increased in the SW1990 cells (38.3±2.25, vs. 52.3±3.5%,
P<0.05) and PANC-1 cells (39±4, vs. 50.3±3.5%, P<0.05;
Fig. 3B). These observations
demonstrated that loss of the expression of SMG-1 inhibited the
proliferation of pancreatic cancer cells via the induction of the
G1 phase of the cell cycle.
Effect of SMG-1-knockdown on the
regulation of pancreatic cancer cell invasion and migration
To determine whether SMG-1 knockdown affected the
invasion and migration of the SW1990 and PANC-1 cells, Matrigel
invasion and Transwell assays were performed. Infection with the
SMG-1 shRNA lentivirus #1 had no significant effect on the tumor
cell invasion and migration capacities (Fig. 4A and B) compared with the cells
infected with the control lentivirus.
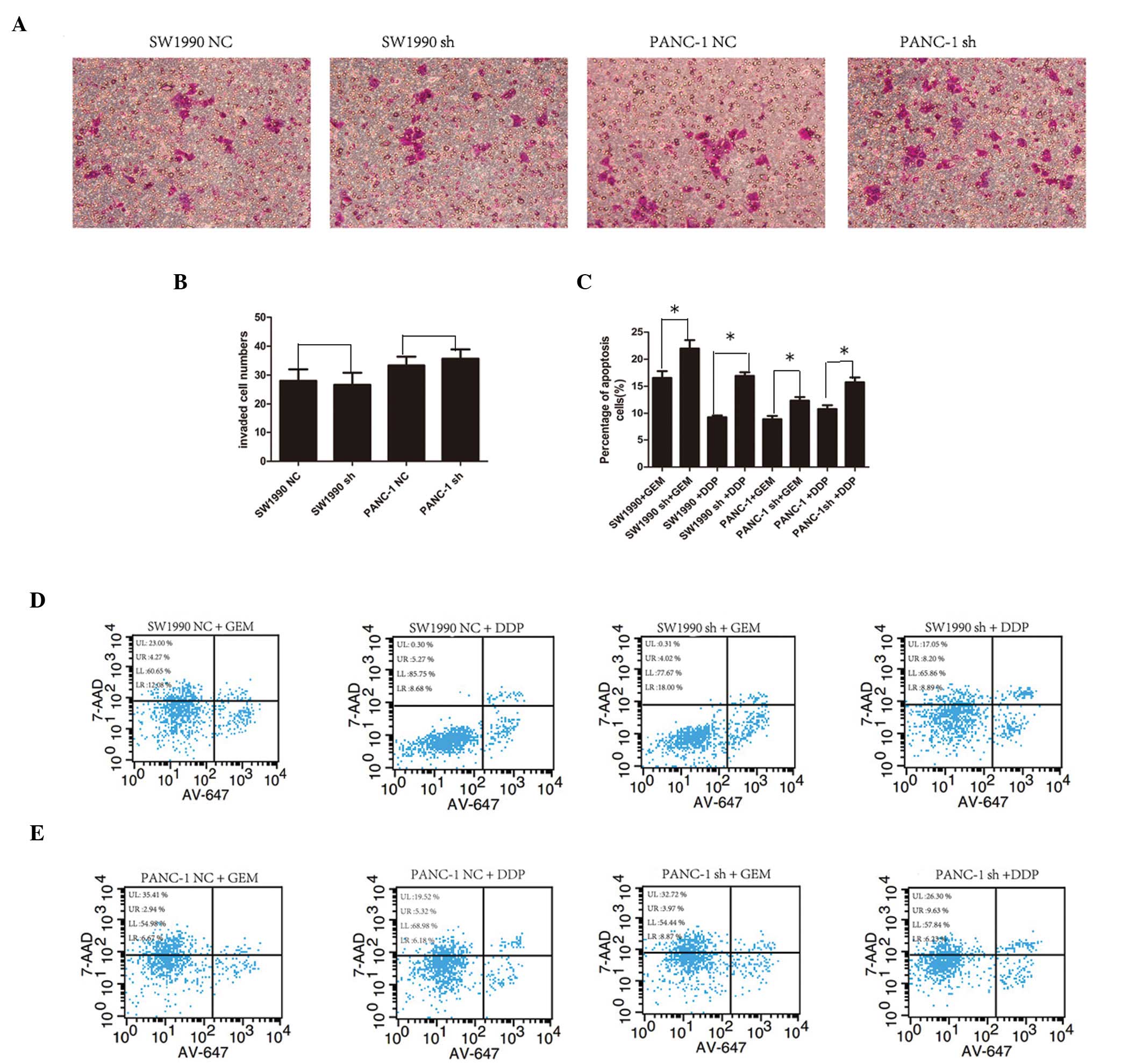 | Figure 4Effect of SMG-1 knockdown on the
regulation of pancreatic cancer cell invasion and sensitivity to
chemotherapeutic drugs. (A) Representative images of tumor cell
invasion and migration (magnification, ×200; 0.1% crystal violet
stain). (B) Summarized data from (A) based on three independent
experiments. (C) Apoptosis of the pancreatic cancer cell lines
following infection with the shSMG-1 or control lentivirus
(*P<0.05, vs. control). Data are expressed as the
mean ± standard deviation. (D and E) Changes in tumor cell
apoptotic rate. LR+UR is the total apoptotic rate. SMG-1,
serine/threonine-protein kinase 1; NC, negative control; sh,
short-hairpin; GEM, gemcitabine; DDP, cisplatin; 7-AAD,
7-aminoactinomycin D. UL, upper left; UR, upper right; LL, lower
left; LR, lower right. |
SMG-1 knockdown increases the
chemosensitivity of pancreatic cancer cells to treatment with
gemcitabine and cisplatin
Following infection of the pancreatic cancer cells
with SMG-1 shRNA and negative control lentiviruses for 72 h, the
cells were treated with 10 mg/ml gemcitabine or 10 mg/ml cisplatin
for an additional 24 h. Flow cytometric analysis revealed that the
levels of apoptosis in the SW1990 cells treated with
gemcitabine/cisplatin increased, between 16.55±1.26 and 21.97±1.55%
in the gemcitabine-treated cells compared with control cells, and
between 9.28±0.34 and 16.93±0.66% in the cisplatin-treated cells
(Fig. 4C and D). Similarly, the
PANC-1 cells demonstrated similar responses when treated with
gemcitabine (8.94±0.59, vs. 12.35±0.66% and cisplatin (10.8±0.7,
vs. 15.74±0.89%; Fig. 4C and
E).
Discussion
The present study detected the protein expression
levels of SMG-1 in pancreatic cancer and normal tissues, and found
that the expression levels of SMG-1 were increased in pancreatic
cancer tissue compared with normal tissue. This finding is
consistent with data from the Human Protein Atlas. It was also
revealed that the protein expression of SMG-1 was associated with
an advanced tumor stage. Subsequently, the expression of SMG-1 was
knocked down in pancreatic cancer cell lines and phenotypic changes
in the tumor cells were observed. The data demonstrated that knock
down of SMG-1 inhibited the proliferation of pancreatic cancer
cells and increased tumor cell chemosensitivity. However, the tumor
cell invasion and migration capacities remained unaltered.
SMG-1 is the newest member of the PIKK family
(4,5) due to its homology with CeSMG-1
(5). SMG-1 is involved in NMD of
mRNA, and previous studies have demonstrated that abnormal SMG-1
function is involved in human cancer (5,8,17).
Other investigations have revealed that SMG-1 is important in human
carcinogenesis and cancer progression (12,18,19),
and may be a tumor suppressor gene (5,8,9,20,21).
These studies demonstrated that SMG-1 is a stress-responsive enzyme
and regulates the cell cycle G1/S checkpoint, while silencing of
SMG-1 increases tumor cell growth (10,21).
By contrast, downregulation of the expression of SMG-1 is
associated with improved prognosis in head and neck cancer
(12).
In the present study, high protein expression levels
of SMG-1 were found in pancreatic cancer tissues and cell lines. In
addition, the protein expression of SMG-1 was associated with an
advanced tumor stage, although only limited clinicpathological data
was available from the TMA company. These findings were consistent
with data from the Human Protein Atlas.
Previous studies have demonstrated that SMG-1 is
involved in multiple biological processes, including cell
proliferation, apoptosis and stress responses (6,13,20,22,23).
The present study utilized an shRNA technique to knock down the
expression of SMG-1 to assess the functions of SMG-1 in pancreatic
cancer cell lines (24). The SMG-1
shRNA lentivirus #1 significantly reduced the expression of SMG-1
in two pancreatic cancer cell lines (Fig. 2B). SMG-1 knockdown inhibited cell
proliferation and increased the chemosensitivity of the cells to
gemcitabine and cisplatin treatment in vitro. Previous
studies have reported that the loss of SMG-1 function significantly
increases the rate and extent of apoptotic tumor cell death induced
by chemotherapy, irradiation or cytokine treatment (12,13).
Another study demonstrated that SMG-1-depleted human cells exhibit
an increased level of spontaneous DNA damage (7).
Since the majority pancreatic cancer patients (~75%)
are diagnosed at an advanced stage, chemotherapy is a tentative
treatment option (25). SMG-1
shRNA may assist in treating patients with chemotherapy-resistant
tumors. A previous study demonstrated that human papillomavirus
(HPV) head and neck cancer cells and tissues express SMG-1 at lower
levels compared with HPV-negative cancer tissues, and depletion of
SMG-1 in HPV-negative head and neck cancer cells increases the
sensitivity to radiation and chemotherapy (12). Another study reported that the
sensitivity of lung cancer cells to gemcitabine and cisplatin
increased following silencing of the expression of SMG-1 using
siRNA (26). The present study
demonstrated similar results in pancreatic cancer tissues and cell
lines. However, this was only proof-of-principle and further
studies are required to fully elucidate the role of SMG-1 protein
in the development and progression of pancreatic cancer, by
investigating the in vivo effects of SMG-1 knockdown on the
chemosensitivity of pancreatic cancer cells.
Acknowledgments
The authors would like to thank Dr Yi Miao and Dr
Min Tu of the Department of General Surgery (Nanjing Medical
University, Nanjing, China) for providing assistance and technical
support. The authors would also like to thank Medjaden Bioscience,
Ltd. (Hong Kong, China) for assisting in the preparation of this
manuscript.
References
|
1
|
Yadav D and Lowenfels AB: The epidemiology
of pancreatitis and pancreatic cancer. Gastroenterology.
144:1252–1261. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ghadirian P, Lynch HT and Krewski D:
Epidemiology of pancreatic cancer: an overview. Cancer Detect Prev.
27:87–93. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Han H and Von Hoff DD: SnapShot:
pancreatic cancer. Cancer Cell. 23:424. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yamashita A, Ohnishi T, Kashima I, Taya Y
and Ohno S: Human SMG-1, a novel phosphatidylinositol
3-kinase-related protein kinase, associates with components of the
mRNA surveillance complex and is involved in the regulation of
nonsense-mediated mRNA decay. Genes Dev. 15:2215–2228. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Denning G, Jamieson L, Maquat LE, Thompson
EA and Fields AP: Cloning of a novel phosphatidylinositol
kinase-related kinase: characterization of the human SMG-1 RNA
surveillance protein. J Biol Chem. 276:22709–22714. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Gehen SC, Staversky RJ, Bambara RA, Keng
PC and O’Reilly MA: hSMG-1 and ATM sequentially and independently
regulate the G1 checkpoint during oxidative stress. Oncogene.
27:4065–4074. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Brumbaugh KM, Otterness DM, Geisen C, et
al: The mRNA surveillance protein hSMG-1 functions in genotoxic
stress response pathways in mammalian cells. Mol Cell. 14:585–598.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Greenman C, Stephens P, Smith R, et al:
Patterns of somatic mutation in human cancer genomes. Nature.
446:153–158. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Uhlen M, Oksvold P, Fagerberg L, et al:
Towards a knowledge-based Human Protein Atlas. Nat Biotechnol.
28:1248–1250. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Gubanova E, Issaeva N, Gokturk C,
Djureinovic T and Helleday T: SMG-1 suppresses CDK2 and tumor
growth by regulating both the p53 and Cdc25A signaling pathways.
Cell Cycle. 12:3770–3780. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
Erker L, Schubert R, Yakushiji H, et al:
Cancer chemoprevention by the antioxidant tempol acts partially via
the p53 tumor suppressor. Hum Mol Genet. 14:1699–1708. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gubanova E, Brown B, Ivanov SV, et al:
Downregulation of SMG-1 in HPV-positive head and neck squamous cell
carcinoma due to promoter hypermethylation correlates with improved
survival. Clin Cancer Res. 18:1257–1267. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Oliveira V, Romanow WJ, Geisen C, et al: A
protective role for the human SMG-1 kinase against tumor necrosis
factor-α-induced apoptosis. J Biol Chem. 283:13174–13184. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Tiedemann RE, Zhu YX, Schmidt J, et al:
Kinome-wide RNAi studies in human multiple myeloma identify
vulnerable kinase targets, including a lymphoid-restricted kinase,
GRK6. Blood. 115:1594–1604. 2010. View Article : Google Scholar :
|
|
15
|
Neben K, Schnittger S, Brors B, et al:
Distinct gene expression patterns associated with FLT3- and
NRAS-activating mutations in acute myeloid leukemia with normal
karyotype. Oncogene. 24:1580–1588. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
17
|
Chen RQ, Yang QK, Chen YL, et al: Kinome
siRNA screen identifies SMG-1 as a negative regulator of
hypoxia-inducible factor-1α in hypoxia. J Biol Chem.
284:16752–16758. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Noensie EN and Dietz HC: A strategy for
disease gene identification through nonsense-mediated mRNA decay
inhibition. Nat Biotechnol. 19:434–439. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Perrin-Vidoz L, Sinilnikova OM,
Stoppa-Lyonnet D, Lenoir GM and Mazoyer S: The nonsense-mediated
mRNA decay pathway triggers degradation of most BRCA1 mRNAs bearing
premature termination codons. Hum Mol Genet. 11:2805–2814. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
González-Estévez C, Felix DA, Smith MD, et
al: SMG-1 and mTORC1 act antagonistically to regulate response to
injury and growth in planarians. PLoS Genet. 8:e10026192012.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Roberts TL, Ho U, Luff J, et al: Smg1
haploinsufficiency predisposes to tumor formation and inflammation.
Proc Natl Acad Sci USA. 110:E285–294. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Masse I, Molin L, Mouchiroud L, et al: A
novel role for the SMG-1 kinase in lifespan and oxidative stress
resistance in Caenorhabditis elegans. PLoS One. 3:e33542008.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Abraham RT: PI 3-kinase related kinases:
‘big’ players in stress-induced signaling pathways. DNA Repair
(Amst). 3:883–887. 2004. View Article : Google Scholar
|
|
24
|
Brummelkamp TR, Bernards R and Agami R: A
system for stable expression of short interfering RNAs in mammalian
cells. Science. 296:550–553. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Philip PA: Targeted therapies for
pancreatic cancer. Gastrointest Cancer Res. 2:S16–S19. 2008.
|
|
26
|
Xia QS, Li YZ, Li HY, et al: Effect of
inhibiting the expression of hSMG-1 on chemosensitivity of human
non-small cell lung cancer H1299 cells. Zhonghua Yi Xue Za Zhi.
91:554–559. 2011.In Chinese. PubMed/NCBI
|