Introduction
Infantile spasms, also known as the West syndrome,
constitute a unique, age-specific epileptic encephalopathy of early
infancy and are characterized by epileptic spasms with psychomotor
regression and electroencephalographic (EEG) indication of
hypsarrhythmia (1,2). The incidence of infantile spasms has
been estimated to be ~0.31 per 1,000 live births (3). The peak age of onset of infantile
spasms is three to seven months, and the condition is slightly more
common in males (4). A number of
underlying disorders, including perinatal brain injury, metabolic
disorders and chromosome anomaly, may be the etiologies of
infantile spasms (5). However, in
~20% of the cases, no causes are identified (6).
While >200 types of etiologies have been involved
in the pathogenesis of infantile spasms, the pathophysiology of
infantile spasms remains largely elusive (7). Previous studies showed that pre-natal
adverse stress is associated with infantile spasms in animal models
and clinical epidemiological analyses (8,9).
Shang et al (8) found that
the onset risk of infantile spasms correspondingly increased with
the degree of maternal pre-natal stress. The stress may act on the
developing brain and results in the dyssecretosis of the
hypothalamic-pituitary-adrenal (HPA) axis and high levels of the
corticotrophin-releasing hormone (CRH), thereby causing spasms
(10). Baram and Schultz (11) established a CRH rat model to verify
the HPA dysfunction hypothesis. CRH administered into the cerebral
ventricles of rats during the first post-natal week caused a
specific and stereotyped sequence of behaviors: Rhythmic chewing
and licking were followed by ‘limbic’-type seizures. This condition
suggested that CRH may be a strong inducer of spasms, and its
excessive release may be the final common pathway through which
infantile spasms were induced by various factors. On the other
hand, 50–80% of infantile spasm cases can be successfully treated
with adrenocorticotropic hormone (ACTH), although infantile spasms
are difficult to control with most conventional anti-epileptic
drugs (12). The clinical response
of infantile spasms to ACTH can be explained by the suppression of
CRH production and release via a negative feedback mechanism
(13).
The effect of CRH is mediated via CRH receptors in
the cell membrane of effector organs (14). Two major CRH receptors, CRHR1 and
CRHR2, have been identified and have functional differences between
them (15). CRHR1 is a G
protein-coupled receptor localized in frontal cortical areas,
forebrain, brainstem, amygdala, cerebellum and the anterior
pituitary (16). CRHR1 has a key
role in the regulation of the HPA axis in response to stressful
events, mediating the action of CRH on the pituitary gland to
release ACTH that stimulates the production of cortisol in the
adrenal cortex (17). Previous
studies have shown that polymorphisms in the coding and regulatory
regions of CRHR1 gene are associated with the onset of certain
neuropsychiatric disorders, including major depression, bipolar
disorder, alcoholism and others (18–20).
However, whether genetic variants of CRHR1 are associated with
infantile spasms, has not been studied in depth. In the present
study, the CRHR1 gene was analyzed as a candidate to test its
association with the onset of infantile spasms, as well as its
responsiveness to ACTH treatment.
Materials and methods
Study subjects
A total of 128 patients with infantile spasms and
131 healthy controls were recruited to perform the case-control
analysis. The cases were unrelated Chinese individuals (83 boys, 45
girls; mean age, 6.8±2.9 months), recruited between January 2006
and May 2010 from the Chinese PLA General Hospital and Beijing
Children’s Hospital affiliated to Capital Medical University
(Beijing, China). The diagnostic criteria for infantile spasms were
as follows: 1) The presence of epileptic spasms; 2) hypsarrhythmia
or modified hypsarrhythmia indicated by EEG prior to treatment; and
3) onset of the infantile spasms at an age between one month and
two years (12). According to
etiologies, infantile spasms were classified as symptomatic or
cryptogenic, as defined by the International League Against
Epilepsy (21). Clinical
evaluation was performed by experienced pediatric neurologists.
Individuals were excluded from the study if they had received
hormone therapy within 28 days prior to recruitment or if they
displayed contraindications to hormone therapy. The control
subjects were all unrelated Chinese individuals (87 boys, 44 girls;
mean age, 7.5±3.7 months) without any nervous system diseases,
particularly epilepsy, or family history of epilepsy. No
statistical difference existed between the patients and controls in
terms of gender and age (Table I).
All subjects of the present study were Han Chinese, and their
parents were asked to sign written informed consent prior to
enrolement in this study. The study was approved by the Committee
on Human Study of Beijing Children’s Hospital affiliated to Capital
Medical University (Beijing, China).
 | Table IDistribution of selected variables in
cases (n=128) and controls (n=131). |
Table I
Distribution of selected variables in
cases (n=128) and controls (n=131).
| Variable | Cases | Controls | P-value |
|---|
| Gender | | | 0.79a |
| Male, n (%) | 83 (64.8) | 87 (66.4) | |
| Female, n (%) | 45 (35.2) | 44 (33.6) | |
| Age (months) | 6.8±2.9 | 7.5±3.7 | 0.88b |
| ACTH
responsiveness | | | |
| Response (n) | 62 | 63.9 | |
| No response (n) | 35 | 36.1 | |
| Etiology | | | |
| Symptomatic (n) | 79 | 70.5 | |
| Cryptogenic (n) | 33 | 29.5 | |
Evaluation of effectiveness of ACTH
treatment
Patients with infantile spasms received ACTH
(Shanghai First Biochemical Pharmaceutical Co., Shanghai, China)
treatment 25 U/day intravenously for 3 weeks. ACTH response was
evaluated mainly based on changes in seizure frequency. Baseline
seizure frequency was assessed by parents or carers. Three weeks
after ACTH treatment initiation, the seizure frequency was assessed
again. Changes in seizure frequency were classified as seizure-free
(cessation of seizure for ≥7 days), reduced (reduction of the
baseline frequency by 50-<100%), unchanged (reduction of the
baseline frequency by <50%), or increased (>100%). The rate
of response, defined as a cessation and reduction of the baseline
seizure frequency by ≥50%, was calculated in each group. No change
or increased frequency was defined as no response.
Selection of single nucleotide
polymorphisms (SNPs)
The human CRHR1 gene (NM_004382) contains 13 exons
and spans 51.5 kb of the sequence on chromosome 17. Based on the
HapMap Genome Browser (Phase 1 and 2 – full dataset; http://hapmap.ncbi.nlm.nih.gov/), the tagSNP and
functional SNP strategy with a minor allele frequency (MAF)>0.1
were used on a Chinese population, and five SNPs were selected for
genotyping in the present study. One SNP, rs28364026, in the 3′
untranslated region (3′UTR), one SNP, rs242948, in the 3′ neargene
region, and all other three SNPs were in introns.
Genotyping assay
Genomic DNA was extracted from peripheral blood
lymphocytes using the Qiagen QIAamp DNA Blood mini kit (Qiagen
Inc., Hilden, Germany) according to the manufacturer’s
instructions. Genotyping primers were designed using the Primer3
software (http://frodo.wi.mit.edu/cgi-bin/primer3/primer3_results.cgi),
and the sequences are listed in Table
II. Multiplex polymerase chain reactions (PCR) were used to
amplify all fragments in a total reaction volume of 20 μl.
The amplification reactions contained 1X HotStarTaq buffer, 3.0 mM
Mg2+, 0.3 mM desoxynucleoside triphosphate, 1U
HotStarTaq polymerase (Qiagen Inc.), 10 ng sample DNA and 1
μl multiple PCR primer. All primers were supplied by Beijing
SaiBaiSheng Gene Technology Co., Ltd. (Beijing, China). The PCR
amplification was performed using a PTC-200 thermocycler (MJ
Research, Watertown, MA, USA). The cycling conditions for PCR in
all reactions included an initial activation step at 95°C for 15
minutes, followed by 11 cycles of denaturation at 94°C for 20
seconds, touchdown annealing at 65°C for 40 seconds −0.5°C per
cycle, extension at 72°C for 90 seconds, followed by another 24
cycles of denaturation at 94°C for 20 seconds, annealing at 59°C
for 30 seconds, extension at 72°C for 90 seconds, and finally,
extension at 72°C for 4 minutes. After the completion of PCR
amplification, phosphatase alkaline shrimp (SAP) and exonuclease I
(Exo I) were used to purify the PCR products. One unit of SAP and 1
unit of Exo I were added to 5 μl PCR mixture. The mixture
was incubated at 37°C for 1 hour, followed by inactivation at 75°C
for 15 minutes. Snap-shot multiplex single-base extension was
performed using the SnaPshot Multiplex kit (Applied Biosystems,
Life Technologies, Thermo Fisher Scientific, Waltham, MA, USA) with
five different extension primers. The 10-μl extension mix
contained 5 μl SnaPshot Multiplex reagent, 2 μl
purified multiplex PCR products, 1 μl extension primer mix
and 2 μl double distilled H2O. The extension PCR
cycling conditions included an initial activation step at 96°C for
1 minute, followed by 28 cycles of denaturation at 96°C for 10
seconds, annealing at 50°C for 5 seconds and extension at 60°C for
30 seconds. The 10-μl extension PCR product obtained by
extension PCR was purified with 1U SAP at 37°C for 1 hour, followed
by an inactivation at 75°C for 15 minutes. Pyrosequencing was
performed on the ABI3130XL sequencer (Applied Biosystems). Data
were collected and analyzed using GeneMapper 4.0 (Applied
Biosystems).
| Table IIPrimer information of NR3C1 for
snapshot assay. |
Table II
Primer information of NR3C1 for
snapshot assay.
| rs number | Direction | Primer
sequence | Tm (°C) | Product size
(bp) |
|---|
| rs171440 | Forward
Reverse |
GGATGGGTCTGTTCCAGGGTGT
TGGTCCCCTGCTCTGTAGCCTAA | 66.09
65.96 | 214 |
| rs4458044 | Forward
Reverse |
TGGGCTCCCCTCTTCTGTGAG
CTGCCTTCCCTTCCCTCCTCTT | 65.66
64.42 | 106 |
| rs17689966 | Forward
Reverse |
CCTTCTGCCAGGGTTGGAATTG
ATCATGGGGCCCTGGTAGATGT | 64.63
64.92 | 212 |
| rs28364026 | Forward
Reverse |
CTGCCTGTGGAGGTGACCTGTT
CCCCATTTCAATTCATTCCCATGT | 63.68
64.11 | 158 |
| rs242948 | Forward
Reverse |
ATGGGTCAGGGGAAGGAACAAA
TGGAGACAGCTGCATTCACAGC | 64.59
64.70 | 176 |
Statistical analysis
Differences in gender as well as frequencies of
alleles and genotypes between cases and controls were tested using
Pearson’s χ2 test. An independent-samples T test was
used to examine the difference in age between controls and cases.
Pearson’s χ2 test was also employed to examine the
different allelic frequencies of ACTH effectiveness and etiology in
cases. The Hardy-Weinberg equilibrium test was performed for every
SNP in the controls. The linkage disequilibrium (LD) blocks were
reconstructed with D′ and r2 using Haploview software (http://www.broadinstitute.org/haploview)
and pairwise linkage disequilibrium values D’ and r2 were
calculated in the control population using the maximum likelihood
method. Haplotype blocks were defined using the method of Gabriel
et al (22). Each samples’
haplotype and dihaplotype was estimated from the genotype data
using PHASE 2.0.2 software (available on request). Odds ratios (OR)
and 95% confidence intervals (95% CI) were calculated using
unconditional logistic regression analysis to evaluate the
association between the risk of infantile spasms and each SNP or
haplotype or dihaplotype, and adjusted to age and gender. All
statistical tests were performed using SPSS 20.0 software
(International Business Machines, Armonk, NY, USA). Two-sided
P-values<0.05 were considered to indicate statistically
significant differences between values.
Results
In the present study, 128 cases with infantile
spasms and 131 controls were examined who were all unrelated Han
Chinese children. There was no significant difference between cases
and control group regarding gender and age (P>0.05). Table I shows basic data on the
subjects.
The positions and MAFs of five SNPs of the CRHR1
gene in the National Center for Biotechnology Information database
for Han Chinese in Beijing are shown in Table III. Each SNP was examined with
Hardy-Weinberg equilibrium among controls, and no significant
difference was observed (P>0.05). Hence, the population was
qualified for further analyses. The genotyping rates of five SNPs
were all >99.6%.
| Table IIIInformation on five genotyped SNPs of
CRHR1 (OMIMa no. 122561; locus
17ql2-q22). |
Table III
Information on five genotyped SNPs of
CRHR1 (OMIMa no. 122561; locus
17ql2-q22).
| NCBI SNP ID | Chromosome
positionb | Location in gene
region | Base change | MAF
| P-valued | P-value for
HWEe | Genotyping rate
(%) |
|---|
| Databasec | Cases | Controls |
|---|
| rs4458044 | 43873727 | intron | G>C | 0.15 | 0.305 | 0.294 | 0.79 | 0.58 | 100 |
| rs171440 | 43893487 | intron | C>T | 0.267 | 0.125 | 0.126 | 0.97 | 1 | 100 |
| rs17689966 | 43910455 | intron | G>A | 0.238 | 0.172 | 0.181 | 0.79 | 1 | 99.6 |
| rs28364026 | 43912294 | 3′UTR | G>A | 0.261f | 0.207 | 0.202 | 0.89 | 1 | 100 |
| rs242948 | 43913544 | 3′neargene | C>A | 0.227f | 0.172 | 0.164 | 0.81 | 0.94 | 100 |
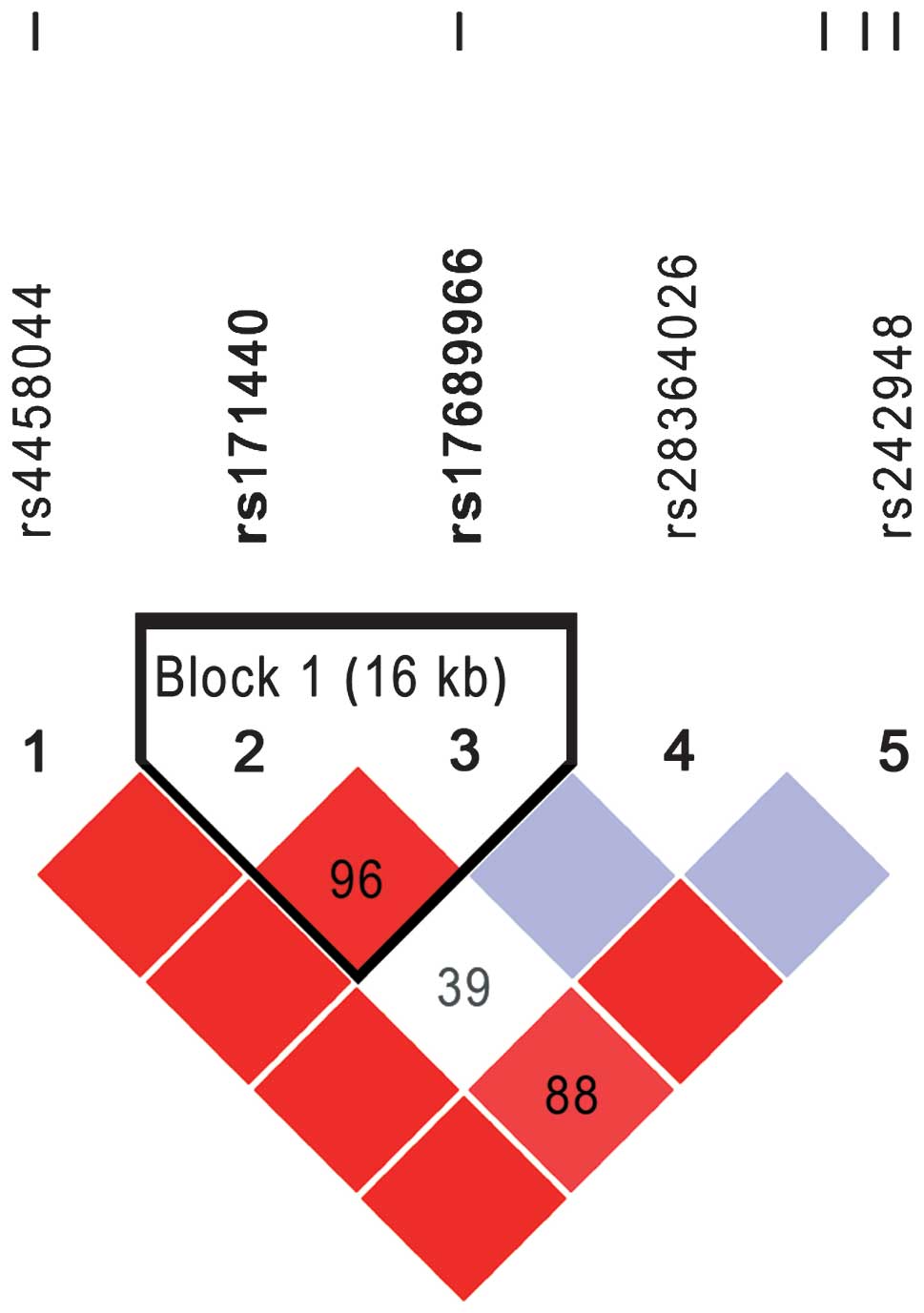
The association of five SNPs in the CRHR1 gene was
examined in a dominant model adjusted to age and gender, but no
significant differences were identified by Pearson’s χ2
test between cases and controls regarding each SNP (Table IV). The LD block of the five SNPs
was reconstructed using the genotyping data of the controls with
the haploview software. Fig. 1
shows the LD blocks among five SNPs, where two SNPs, rs171440 and
rs17689966, were able to form an LD block. PHASE software was used
to estimate the possible haplotypes and diplotypes between rs171440
and rs17689966. There were three haplotypes and three common
diplotypes, but none of them was found to be associated with the
risk of infantile spasms (Table
V).
| Table IVGenotypes of five single nucleotide
polymorphisms of CRHR1 and their associations with infantile
spasms. |
Table IV
Genotypes of five single nucleotide
polymorphisms of CRHR1 and their associations with infantile
spasms.
| Genetic model | Genotype | Controls, n
(%) | Cases, n (%) | P-valuea | Logistic
regressionb
|
|---|
| OR (95%
CI)c |
P-valued |
|---|
| rs4458044 | | | | | | |
| Co-dominant | GG
GC
CC | 67 (51.1)
51 (38.9)
13 (10) | 57 (44.5)
64 (50)
7 (5.5) | 0.13 | 1.00
1.47 (0.88–2.45)
0.63 (0.23–1.68) | 0.14
0.35 |
| Dominant | GC+CC | 64 (48.9) | 71 (55.5) | 0.29 | 1.30
(0.80–2.12) | 0.29 |
| rs171440 | | | | | | |
| Co-dominant | AA
AG
GG | 100
(76.3)
29 (22.1)
2 (1.6) | 96 (75)
32 (25)
0 | 0.33 | 1.00
1.15 (0.65–2.05) | 0.64
0.99 |
| Dominant | AG+GG | 31 (23.7) | 32 (25) | 0.80 | 1.08
(0.61–1.91) | 0.80 |
| rs17689966 | | | | | | |
| Co-dominant | AA
AG
GG | 87 (66.9)
39 (30)
4 (3.1) | 88 (68.8)
36 (28.1)
4 (3.1) | 0.78 | 1.00
0.92 (0.53–1.60)
0.98 (0.24–4.06) | 0.77
0.97 |
| Dominant | AG+GG | 43 (33.1) | 40 (31.2) | 0.75 | 0.93
(0.55–1.57) | 0.78 |
| rs28364026 | | | | | | |
| Co-dominant | GG
GA
AA | 83 (63.4)
43 (32.8)
5 (3.8) | 79 (61.7)
45 (35.2)
4 (3.1) | 0.90 | 1.00
1.10 (0.66–1.86)
0.85 (0.22–3.29) | 0.71
0.81 |
| Dominant | GA+AA | 48 (36.6) | 49 (38.3) | 0.79 | 1.08
(0.65–1.78) | 0.77 |
| rs242948 | | | | | | |
| Co-dominant | TT
TG
GG | 92 (70.2)
35 (26.7)
4 (3.1) | 88 (68.8)
36 (28.1)
4 (3.1) | 0.97 | 1.00
1.09 (0.63–1.90)
1.03 (0.25–4.26) | 0.76
0.97 |
| Dominant | TG+GG | 39 (29.8) | 40 (31.2) | 0.80 | 1.08
(0.63–1.85) | 0.77 |
| Table VAssociation of haplotypes and
diplotypes with risk of infantile spasms. |
Table V
Association of haplotypes and
diplotypes with risk of infantile spasms.
| Types | Controls, n
(%) | Cases, n (%) | Logistic
regressiona
|
|---|
| OR (95% CI) | P-valueb |
|---|
| Haplotypesc | | | | |
| AA | 214 (82) | 212 (82.8) | 1.00 | |
| AG | 15 (5.7) | 12 (4.7) | 0.82
(0.37–1.80) | 0.62 |
| GG | 32 (12.3) | 32 (12.5) | 1.01
(0.60–1.71) | 0.98 |
| Diplotypesc | | | | |
| AA/AA | 88 (67.2) | 88 (68.8) | 1.00 | |
| AA/AG | 12 (9.2) | 8 (6.3) | 0.68
(0.26–1.78) | 0.43 |
| AA/GG | 26 (19.8) | 28 (21.9) | 1.08
(0.59–2.00) | 0.80 |
| Othersd | 5 (3.8) | 4 (3) | 0.80
(0.21–3.08) | 0.74 |
A total of 97 patients with infantile spasms
received ACTH treatment for three weeks, among which 62 cases
(63.9%) showed responses and 35 cases (36.1%) were unresponsive.
The association of ACTH responsiveness and the genotypes of SNPs
was analyzed. Under the assumption of a co-dominant model, there
was no association of the effectiveness of ACTH and the etiology of
infantile spasms with any single SNP (Table VI).
| Table VIAssociation of the effectiveness of
ACTH and etiology of infantile spasms with single nucleotide
polymorphisms of CRHR1. |
Table VI
Association of the effectiveness of
ACTH and etiology of infantile spasms with single nucleotide
polymorphisms of CRHR1.
| Genetic model | Genotype | ACTH
| P-valuea | Logistic
regressionb OR (95% CI) |
P-valuec | Etiology
| P-valuea | Logistic
regressionb OR(95% CI) |
P-valuec |
|---|
| Effective n(%) | Ineffective
n(%) | Symptomatic
n(%) | Cryptogenic
n(%) |
|---|
| rs4458044 | | | | | | | | | | | |
| Co-dominant | GG
GC
CC | 27 (43.5)
32(51.6)
3 (4.9) | 16 (45.7)
18 (51.4)
1 (2.9) | 0.89 | 1.00
0.97 (0.42–2.28)
0.53 (0.05–5.64) | 0.95
0.60 | 37 (46.8)
38(48.1)
4(5.1) | 15 (45.5)
17(51.5)
1 (3.0) | 0.87 | 1.00
1.14(049–2.62)
0.65 (0.07–647) | 0.76
0.71 |
| Dominant | GC+CC | 35 (56.5) | 19 (54.3) | 0.84 |
0.93(0.40–2.16) | 0.87 | 42(53.1) | 18 (54.5) | 0.90 | 1.09(048–248) | 0.84 |
| rs171440 | | | | | | | | | | | |
| Co-dominant | AA
AG | 14 (22.6)
48 (77.4) | 9 (25.7)
26 (74.3) | 0.73 | 1.00
1.16(0.44–3.05) | 0.77 | 58 (73.4)
21 (26.6) | 26 (78.8)
7(21.2) | 0.55 | 1.00
0.69(0.26–1.84) | 0.46 |
| rs17689966 | | | | | | | | | | | |
| Co-dominant | AA
AG
GG | 45 (72.6)
14 (22.6)
3 (4.8) | 23 (65.7)
12 (34.3)
0 | 0.23 | 1.00
1.66(0.65–423)
0 | 0.29
0.99 | 53(67.1)
24 (30.4)
2 (2.5) | 24 (72.7)
7(21.2)
2(6.1) | 0.44 | 1.00
0.56(0.21–1.53)
2.19(0.28–17.13) | 0.26
0.46 |
| Dominant | AG+GG | 17 (27.4) | 12 (34.3) | 0.48 |
1.35(0.55–3.32) | 0.59 | 26 (32.9) | 9 (27.3) | 0.57 |
0.68(0.27–1.72) | 0.42 |
| rs28364026 | | | | | | | | | | | |
| Co-dominant | GG
GA
AA | 41 (66.1)
19 (30.6)
2 (3.3) | 19 (54.3)
14 (40)
2 (5.7) | 0.49 | 1.00
1.58(0.65–3.83)
242(0.31–18.94) | 0.31
0.40 | 48 (60.8)
29 (36.7)
2 (2.5) | 21 (63.6)
10 (30.3)
2(6.1) | 0.57 | 1.00
0.76(0.31–1.85)
2.69 (0.35–20.97) | 0.54
0.35 |
| Dominant | GA+AA | 21 (33.9) | 16 (45.7) | 0.25 |
1.66(0.79–3.89) | 0.25 | 31 (39.2) | 12 (36.9) | 0.78 | 0.87
(0.37–2.03) | 0.74 |
| rs242948 | | | | | | | | | | | |
| Co-dominant | TT
TG
GG | 45 (72.6)
14 (22.6)
3 (4.8) | 23 (65.7)
12 (34.3)
0 | 0.23 | 1.00
1.66(0.65–423)
0 | 0.29
0.99 | 53(67.1)
24 (30.4)
2 (2.5) | 24 (72.7)
7(21.2)
2(6.1) | 0.44 | 1.00
0.56(0.21–1.53)
2.19(0.28–17.13) | 0.26
0.46 |
| Dominant | TG+GG | 17 (27.4) | 12 (34.3) | 0.48 |
1.35(0.55–3.32) | 0.52 | 26 (32.9) | 9 (27.3) | 0.56 |
0.68(0.27–1.72) | 0.42 |
Discussion
Infantile spasms are a devastating epileptic
syndrome in children and have unique features, including
characteristic epileptic spasms during infancy, specific
electrographic hypsarrhythmia and the arrest of psychomotor
development (23). However, the
exact pathophysiology of infantile spasms has remained elusive. In
the CRH-excess hypothesis, an excessively abnormal secretion of CRH
induced by different stressors in the developing brain is thought
to increase excitatory amino acid neurotransmission, thereby
causing spasms (24). ACTH being
the only effective drug for infantile spasms apart from vigabatrin
indicates that infantile spasms may be fundamentally different from
other epileptic syndromes (1). The
clinical response to ACTH can be explained by the suppression of
CRH production through negative feedback (10). Based on the effectiveness of ACTH
treatment and the HPA axis dysfunction hypothesis, Shi et al
(25) have proposed a pre-natal
stress exposure hypothesis, stating that diverse etiological
factors are the onset foundations of infantile spasms, whereas
adverse stress during the perinatal period is the onset condition
of infantile spasms.
In spite of the large amount of available studies on
infantile spasms, genetic studies on the association between
polymorphisms of genes and infantile spasms are limited. SNPs are
the most common genetic variations with a frequency >1% in
humans. Liu et al (26)
found that the haplotype TCCT in the MC2R promoter is strongly
associated with the responsiveness of ACTH therapy in patients with
infantile spasms. Further, experiments were conducted to evaluate
the function of TCCT in vitro, demonstrating that TCCT leads
to an increased expression of MC2R and a strong response to ACTH
(27). Ding et al (28) also reported that the response to
ACTH treatment in the CTA haplotype of GRIN1 in homozygous-carriers
was higher than that in heterozygous-carriers and non-carriers.
The CRHR1 gene, located at 17q12-q22, encodes a G
protein-coupled receptor that binds neuropeptides of the CRH
family, which are major regulators of the HPA pathway (29). The encoded protein is essential for
the activation of signal transduction pathways that regulate
diverse physiological processes, including stress, reproduction,
immune response and obesity (30).
Polymorphisms in the CRHR1 gene have been implicated in the
susceptibility for certain neuropsychiatric disorders, particularly
under conditions of stress, including major depression, alcoholism
and child abuse (31–33). Geng et al (31) found that a functional polymorphism
in the 3′UTR of the CRHR1 gene (rs28364032) and three haplotypes
containing it showed significant associations with anti-depressant
remission. Haplotype analyses of the CRHR1 gene in 2,533 unrelated
Caucasian individuals identified one haplotype in the proximal
block 1 and two haplotypes in the distal block 2 that showed
nominally significant genotype – traumatic stress interactive
effects on the likelihood of developing alcoholism (corrected
P<0.025) (32). Grabe et
al (33) reported an
association between childhood abuse and the TAT-haplotype of the
CRHR1 gene and adult depression, therefore connecting childhood
adversities and genetic susceptibility to neurological disorders.
The present study hypothesized that CRHR1 is involved in the
development of infantile spasms, and to the best of our knowledge,
no studies have been conducted regarding the association between
the polymorphisms of CRHR1 and infantile spasms to date.
In the present study, CRHR1 was selected as a
candidate gene to investigate its association with the risk of
infantile spasms and the effectiveness of ACTH treatment. Although
cryptogenic cases may rather be associated with hereditary
susceptibility, symptomatic and cryptogenic cases were all enrolled
in the present study, as not all symptomatic pathogens, including
perinatal asphyxia, congenital malformations and tuberous
sclerosis, will result in infantile spasms. Thus, genetic factors
may also contribute to symptomatic disorders, which has indeed been
confirmed by previous studies (34). Under the assumption of the dominant
model, the selected five SNPs, rs4458044, rs171440, rs17689966,
rs28364026 and rs242948, showed no association with the risk of
infantile spasms and the efficacy of ACTH treatment. In addition,
subsequent haplotype analysis suggested that none of them was
associated with the risk of infantile spasms.
In spite of the strong rationale of the present
study, the case-control results did not reveal any statistically
significant associations between polymorphisms in the CRHR1 gene
and infantile spasms. It is, however, possible that rare variants
in the CRHR1 gene may have a role in the onset of infantile spasms
and the effect of such polymorphisms may not have been detected in
the relatively small population of infantile spasm cases enrolled
in the present study. In order to detect the effect of rare
variations in the CRHR1 gene using a case control methodology, a
significantly larger number of infantile spasms and control
individuals would be required to be analyzed. Alternatively, it may
be possible to identify rare but important variations in the CRHR1
gene by directly sequencing regions of the CRHR1 gene in a smaller
sample of infantile spasms and control individuals.
In conclusion, the present study presented a
preliminary evaluation of the role of variations in the CRHR1 gene
in infantile spasms. It was the first attempt to study the impact
of polymorphisms in the CRHR1 gene in Chinese individuals with
infantile spasms. While the present study failed to detect any
significant associations between individual SNPs or haplotypes in
the CRHR1 gene with the infantile spasm phenotype, it is still
possible that variations in this gene may impact the development or
progression of this illness.
Acknowledgments
This study was supported by grants from the National
Natural Science Foundation of China (nos. 30770747, 30801249,
81071036 and 81211140048), the Beijing Municipal Natural Science
Foundation (nos. 7142150, 7081002 and 7042024) and the Capital
Characteristic Clinical Application Research Foundation (no.
Z121107001012056).
References
|
1
|
Hrachovy RA and Frost JD Jr: Infantile
spasms. Handb Clin Neurol. 111:611–618. 2013.PubMed/NCBI
|
|
2
|
Pavone P, Striano P, Falsaperla R, Pavone
L and Ruggieri M: Infantile spasms syndrome, West syndrome and
related phenotypes: What we know in 2013. Brain Dev. 36:739–751.
2014. View Article : Google Scholar
|
|
3
|
Trevathan E, Murphy CC and Yeargin-Allsopp
M: The descriptive epidemiology of infantile spasms among Atlanta
children. Epilepsia. 40:748–751. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Go CY, Mackay MT, Weiss SK, Stephens D,
Adams-Webber T, Ashwal S and Snead OC III; Child Neurology Society:
Evidence-based guideline update: Medical treatment of infantile
spasms Report of the Guideline Development Subcommittee of the
American Academy of Neurology and the Practice Committee of the
Child Neurology Society. Neurology. 78:1974–1980. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Osborne JP, Lux AL, Edwards SW, Hancock E,
Johnson AL, Kennedy CR, Newton RW, Verity CM and O’Callaghan FJ:
The underlying etiology of infantile spasms (West syndrome):
Information from the United Kingdom Infantile Spasms Study (UKISS)
on contemporary causes and their classification. Epilepsia.
51:2168–2174. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sogawa Y and Moshe SL: Catastrophic
epilepsies: Clinical controversies of infantile spasms. Neurol
Asia. 15:5–7. 2010.
|
|
7
|
Paciorkowski AR, Thio LL and Dobyns WB:
Genetic and biologic classification of infantile spasms. Pediatr
Neurol. 45:355–367. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Shang NX, Zou LP, Zhao JB, Zhang F and Li
H: Association between prenatal stress and infantile spasms: A
case-control study in China. Pediatr Neurol. 42:181–186. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang YJ, Zhang Y, Liang XH, Yang G and Zou
LP: Effects of adrenal dysfunction and high-dose
adrenocorticotropic hormone on NMDA-induced spasm seizures in young
Wistar rats. Epilepsy Res. 100:125–131. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Stafstrom CE: Infantile spasms: A critical
review of emerging animal models. Epilepsy Curr. 9:75–81. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Baram TZ and Schultz L:
Corticotropin-releasing hormone is a rapid and potent convulsant in
the infant rat. Brain Res Dev Brain Res. 61:97–101. 1991.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Pellock JM, Hrachovy R, Shinnar S, Baram
TZ, Bettis D, Dlugos DJ, Gaillard WD, Gibson PA, Holmes GL, Nordl
DR, et al: Infantile spasms: A U.S. consensus report. Epilepsia.
51:2175–2189. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Brunson KL, Avishai-Eliner S and Baram TZ:
ACTH treatment of infantile spasms: Mechanisms of its effects in
modulation of neuronal excitability. Int Rev Neurobiol. 49:185–197.
2002.PubMed/NCBI
|
|
14
|
Wypior G, Jeschke U, Kurpisz M and
Szekeres-Bartho J: Expression of CRH, CRH-related peptide and CRH
receptor in the ovary and potential CRH signalling pathways. J
Reprod Immunol. 90:67–73. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zoumakis E, Kalantaridou SN and
Makrigiannakis A: CRH-like peptides in human reproduction. Curr Med
Chem. 16:4230–4235. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hauger RL, Risbrough V, Brauns O and
Dautzenberg FM: Corticotropin releasing factor (CRF) receptor
signaling in the central nervous system: New molecular targets. CNS
Neurol Disord Drug Targets. 5:453–479. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bogdan R, Santesso DL, Fagerness J, Perlis
RH and Pizzagalli DA: Corticotropin-releasing hormone receptor type
1 (CRHR1) genetic variation and stress interact to influence reward
learning. J Neurosci. 31:13246–13254. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Liu Z, Liu W, Yao L, Yang C, Xiao L, Wan
Q, Gao K, Wang H, Zhu F, Wang G, et al: Negative life events and
corticotropin-releasing-hormone receptor1 gene in recurrent major
depressive disorder. Sci Rep. 3:15482013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Heim C, Bradley B, Mletzko TC, Deveau TC,
Musselman DL, Nemeroff CB, Ressler KJ and Binder EB: Effect of
childhood trauma on adult depression and neuroendocrine function:
Sex-specific moderation by CRH receptor 1 gene. Front Behav
Neurosci. 3:412009. View Article : Google Scholar
|
|
20
|
Chen AC, Manz N, Tang Y, Rangaswamy M,
Almasy L, Kuperman S, Nurnberger J Jr, O’Connor SJ, Edenberg HJ,
Schuckit MA, et al: Single-nucleotide polymorphisms in
corticotropin releasing hormone receptor 1 gene (CRHR1) are
associated with quantitative trait of event-related potential and
alcohol dependence. Alcohol Clin Exp Res. 34:988–996. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Engel J Jr; International League Against
Epilepsy (ILAE): A proposed diagnostic scheme for people with
epileptic seizures and with epilepsy: Report of the ILAE Task Force
on Classification and Terminology. Epilepsia. 42:796–803. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Gabriel SB, Schaffner SF, Nguyen H, Moore
JM, Roy J, Blumenstiel B, Higgins J, DeFelice M, Lochner A, Faggart
M, et al: The structure of haplotype blocks in the human genome.
Science. 296:2225–2229. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lux AL: Latest American and European
updates on infantile spasms. Curr Neurol Neurosci Rep. 13:3342013.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Brunson KL, Eghbal-Ahmadi M and Baram TZ:
How do the many etiologies of West syndrome lead to excitability
and seizures? The corticotropin releasing hormone excess
hypothesis. Brain Dev. 23:533–538. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Shi XY, Zou LP, Yang G and Ding YX:
Prenatal stress exposure hypothesis for infantile spasms. Med
Hypotheses. 78:735–737. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Liu ZL, He B, Fang F, Tang CY and Zou LP:
Genetic polymorphisms of MC2R gene associated with responsiveness
to adrenocorticotropic hormone therapy in infantile spasms. Chin
Med J (Engl). 121:1627–1632. 2008.
|
|
27
|
Ding YX, Zou LP, He B, Yue WH, Liu ZL and
Zhang D: ACTH receptor (MC2R) promoter variants associated with
infantile spasms modulate MC2R expression and responsiveness to
ACTH. Pharmacogenet Genomics. 20:71–76. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ding YX, Zhang Y, He B, Yue WH, Zhang D
and Zou LP: A possible association of responsiveness to
adrenocorticotropic hormone with specific GRIN1 haplotypes in
infantile spasms. Dev Med Child Neurol. 52:1028–1032. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wasserman D, Wasserman J and Sokolowski M:
Genetics of HPA-axis, depression and suicidality. Eur Psychiatry.
25:278–280. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Aguilera G, Nikodemova M, Wynn PC and Catt
KJ: Corticotropin releasing hormone receptors: Two decades later.
Peptides. 25:319–329. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Geng LY, Ye DQ, Shi YY, Xu Z, Pu MJ, Li
ZY, Li XL, Li Y and Zhang ZJ: Influence of genetic polymorphisms
involved in the hypothalamic-pituitary-adrenal axis and their
interactions with environmental factors on antidepressant response.
CNS Neurosci Ther. 20:237–243. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ray LA, Sehl M, Bujarski S, Hutchison K,
Blaine S and Enoch MA: The CRHR1 gene, trauma exposure, and
alcoholism risk: A test of G × E effects. Genes Brain Behav.
12:361–369. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Grabe HJ, Schwahn C, Appel K, Mahler J,
Schulz A, Spitzer C, Fenske K, Barnow S, Lucht M, Freyberger HJ, et
al: Childhood maltreatment, the corticotropin-releasing hormone
receptor gene and adult depression in the general population. Am J
Med Genet B Neuropsychiatr Genet. 153B:1483–1493. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Yang G, Zou LP, He B, Ding YX, Wang J, Shi
XY, Sun YH and Jia FY: NR3C1 gene polymorphism for genetic
susceptibility to infantile spasms in a Chinese population. Life
Sci. 91:37–43. 2012. View Article : Google Scholar : PubMed/NCBI
|