Introduction
Bone metabolism is predominantly regulated by two
types of functional cells: Osteoblasts and osteoclasts, which are
responsible for bone formation and resorption, respectively
(1). The formation and remodeling
of bone structures result from the combined action of osteoblasts
and osteoclasts. Bone resorption and formation are highly regulated
in order to maintain adequate bone mass. Osteoblasts also have a
role in the regulation of bone resorption via receptor activator of
nuclear factor-κB (RANK) ligand (RANKL) expression, in response to
bone resorptive stimuli (2). The
binding of RANKL to RANK, which is located at the cell surface of
osteoclastic precursors and mature osteoclasts, stimulates
osteoclastic differentiation and activation (2). Metabolic bone diseases such as
osteoporosis are caused by disordered bone remodeling. Numerous
humoral factors are crucial to the bone remodeling process,
including prostaglandins such as prosta glandin E2, and cytokines
such as interleukin-1 (3).
Osteoprotegerin (OPG) is a secreted protein
synthesized by osteoblasts, which inhibits osteoclastic
differentiation and activation (4). OPG and RANK belong to the tumor
necrosis factor receptor family. OPG acts as a decoy receptor by
binding RANKL, which prevents it from binding to RANK, resulting in
the suppression of bone resorption (4). RANKL knock out mice have been shown
to suffer from severe osteopetrosis (5). The RANK/RANKL/OPG axis is currently
recognized as a major regulatory system for functional osteoclast
forma tion (6).
Bone morphogenetic proteins (BMPs), including
transforming growth factor-β (TGF-β) and activin, are
multifunctional cytokines that belong to the TGF-β superfamily
(7). BMPs promote bone formation
via stimulation of osteoblastic proliferation and differentiation
(8). BMP intracellular signaling
occurs predominantly via the Smad (Smad 1/5/8)-dependent signaling
pathway (5). In addition, previous
studies have suggested that Smad-independent signaling, such as
mitogen-activated protein (MAP) kinase signaling, also participates
in BMP signaling (9,10). A previous study demonstrated that
BMP-4 stimulates osteo calcin synthesis in osteoblast-like MC3T3-E1
cells, and that osteocalcin synthesis is positively regulated by
p38 MAP kinase (11). In addition,
BMP-4 was shown to stimulate the release of vascular endothelial
growth factor (VEGF) via p38 MAP kinase in MC3T3-E1 cells (12). BMP also stimulates OPG production
in osteoblasts (13). However, the
precise mechanism underlying BMP-4-induced OPG synthesis in
osteoblasts remains to be elucidated.
It is widely accepted that polyphenolic compounds in
foods such as fruits and vegetables are beneficial to humans. Among
them, flavonoids exhibit antioxidative, anti-inflammatory and anti
carcinogenic effects (14,15). Resveratrol is a naturally occurring
polyphenolic flavonoid present in grapes and berries, which has
been shown to increase murine life span (16). In addition, the observed low
mortality rates due to coronary heart disease in France may be
associated with the consumption of wine, which is known to contain
abundant resveratrol (17). The
effects of resveratrol are mediated by the longevity gene SIRT1,
which improves the functioning of cells and organs by activating
the NAD+-dependent histone deacet ylase (16). NAD+ is a coenzyme of
oxidoreductase synthesized as a precursor of nicotinamide, which
has an important role in energy acquisition. Regarding the effects
of resveratrol on osteoblasts, a previous study reported that
resveratrol was able to stimulate osteoblastic differentiation
(18). However, the precise
mechanism underlying the effects of resveratrol on bone metabolism
remains to be elucidated.
The present study aimed to investigate the effects
of resveratrol on BMP-4-stimulated OPG synthesis in osteoblast-like
MC3T3-E1 cells.
Materials and methods
Materials
Resveratrol and SB203580 were purchased from EMD
Millipore (Billerica, MA, USA). BMP-4 and mouse OPG ELISA kits were
purchased from R&D Systems, Inc. (Minneapolis, MN, USA).
Phospho specific p38 MAP kinase (cat. no. 4511S), p38 MAP kinase
antibodies (cat. no. 9212) and anti-rabbit IgG, horseradish
peroxidase (HRP)-linked antibodies (cat. no. 7074) were obtained
from Cell Signaling Technology (Danvers, MA, USA). The Enhanced
Chemiluminescence (ECL) Western Blotting Detection system was
obtained from GE Healthcare Life Sciences (Chalfont, UK). α Minimum
Essential medium (α-MEM) was obtained from Sigma Aldrich (St.
Louis, MO, USA). Fetal bovine serum (FBS) was obtained from Gibco
Life Technologies (Grand Island, NY, USA). Polyvinylidene fluoride
membranes were obtained from Bio Rad Laboratories, Inc. (Berkeley,
CA, USA). Resveratrol and SB203580 were dissolved in dimethyl
sulfoxide. The maximum concentration of dimethyl sulfoxide was
0.1%, which did not affect the OPG assay or western blot
analysis.
Cell culture
Cloned osteoblast-like MC3T3 E1 cells were
generously provided by Dr M. Kumegawa (Meikai University, Sakado,
Japan). The cells were derived from newborn mouse calvaria
(19) and were maintained as
previously described (20).
Briefly, the cells were cultured in α-MEM supplemented with 10% FBS
at 37°C in a humidified atmosphere containing 5% CO2.
The cells were subsequently seeded at a density of 5×104
cells/dish onto 35 mm diameter dishes, or at a density of
2×105 cells/dish onto 90 mm diameter dishes in α-MEM
supplemented with 10% FBS. Following a period of five days, the
medium was exchanged for α-MEM supplemented with 0.3% FBS. The
cells were incubated for 48 h prior to further experimentation.
OPG assay
The cultured cells were treated with 10, 30, 50 or
70 µM resveratrol or 10, 20 or 30 µM SB203580 for 60
min, prior to being stimulated with either 70 ng/ml of BMP-4or
vehicle in 1 ml α-MEM supplemented with 0.3% FBS, and incubated for
48 h. The vehicle was a solvent, containing phosphate buffered
saline (PBS) supplemented with 0.1% bovine serum albumin, in which
BMP-4 was dissolved. The conditioned medium was subsequently
collected, and the OPG concentration was measured using the mouse
OPG ELISA kit, according to the manufacturer's instructions. The
absorbance was determined using an ELISA plate reader (Multiskan
JX; Thermo Labsystems, Inc., Franklin, MA, USA).
Western blot analysis
The cultured cells were treated with 10, 30 or 50
µM resveratrol for 60 min, prior to being stimu lated with
either 70 ng/ml BMP-4 or vehicle in 1 ml α-MEM supplemented with
0.3% FBS for 2 h. The cells were then washed twice with PBS prior
to being lysed. Adherent cells were scraped off the dish using a
plastic cell scraper, then the cell suspension was gently
transferred into a microcentrifuge tube in a lysis buffer
containing 62.5 mM Tris/HCl (pH 6.8), 2% SDS, 50 mM dithiothreitol,
and 10% glycerol, and then soni cated with 20 short burst of 1 sec.
SDS-PAGE was performed according to the Laemmli method (21) in 10% polyacrylamide gels. A western
blot analysis was performed as previously described (22). The membranes were blocked with 5%
fat free dry milk in Tris buffered saline with Tween (TBST; 20 mM
Tris/HCl, pH 7.6, 137 mM NaCl, 0.1% Tween 20) for 1 h prior to
incubation with the primary antibodies. The membranes were then
incubated with the primary antibodies targeting phospho specific
p38 MAP kinase and p38 MAP kinase, all at a dilution of 1:1,000 in
5% milk in TBST overnight at 4°C, using goat peroxidase labeled
antibodies and anti-rabbit IgG HRP linked secondary antibody (cat.
no. 7074) at a dilution of 1:1,000 in 5% milk in TBST for 1 h at
room temperature (Cell Signaling Technology). The peroxidase
activity on the polyvinylidene fluoride membrane (Bio-Rad
Laboratories, Inc.) was visualized on an X-ray film using the ECL
Western Blotting Detection system.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
The cultured cells were treated with either 50
µM resveratrol or vehicle for 60 min, prior to being
stimulated with either 70 ng/ml BMP-4or vehicle in α-MEM
supplemented with 0.3% FBS for 3 h. Total RNA was isolated and
reverse transcribed into cDNA using TRIzol® reagent
(Invitrogen Life Technologies, Carlsbad, CA, USA) and Omniscript
Reverse Transcriptase kit (Qiagen Inc., Valencia, CA, USA). RT-qPCR
was performed in capillaries using a Light Cycler 1.2 system and
the Fast Start DNA Master SYBR Green I kit (Roche Diagnostics,
Basel, Switzerland). The sense and antisense primers (primer set
ID: MA026526) for mouse OPG and GAPDH mRNA were purchased from
Takara Bio, Inc. (Otsu, Japan). The primer sequences were as
follows: Forward 5′-CAA TGG CTG GCT TGG TTT CATAG-3′ and reverse
5′-CTG AAC CAG ACA TGA CAG CTGGA-3′. The sense and antisense
primers for mouse GAPDH mRNA were synthesized based on the study of
Simpson et al (23) and
obtained from Sigma Aldrich. The primer sequences were as follows:
Forward 5′-AAC GAC CCC TTC ATT GAC-3′ and reverse 5′-TCC ACG ACA
TAC TCA GCAC-3′. The reaction mixtures were incubated at 95°C for
10 min, followed by 40 cycles at 60°C for 5 sec and 72°C for 7 sec.
The ampli fied products were determined by melting curve analysis
and 1% agarose gel electrophoresis. The mRNA expression levels of
OPG were normalized to those of GAPDH.
Densitometric analysis
A densitometric analysis was performed using ImageJ
1.32 (National Institutes of Health, Bethesda, MA, USA) image
analysis software program. The background subtracted signal
intensity of each phosphoryla tion signal was normalized to the
respective total protein signal and plotted as the fold increase,
as compared with non stimulated control cells.
Statistical analysis
Differences between the mean values for individual
groups were assessed with a one way analysis of variance, followed
by a Bonferroni analysis in order to carry out multiple comparisons
between pairs. Microsoft Excel 2010 (Redmond, WA, USA) was used for
statistical analysis. P<0.05 was considered to indicate a
statistically significant difference. All data are presented as the
mean ± standard error of the mean, and all experiments were
performed in triplicate from three independent cell
preparations.
Results
Effects of resveratrol on
BMP-4-stimulated OPG release in MC3T3-E1 cells
BMP-4has previously been shown to stimulate the
synthesis of osteocalcin and VEGF in osteoblast-like MC3T3 E1 cells
(11,12). In addition, BMP-2 induces the
synthesis of OPG in human osteoblasts (13). The present study investigated
whether BMP-4 stimulated OPG synthesis in MC3T3 E1 cells. The
results indicated that BMP-4 significantly increased the release of
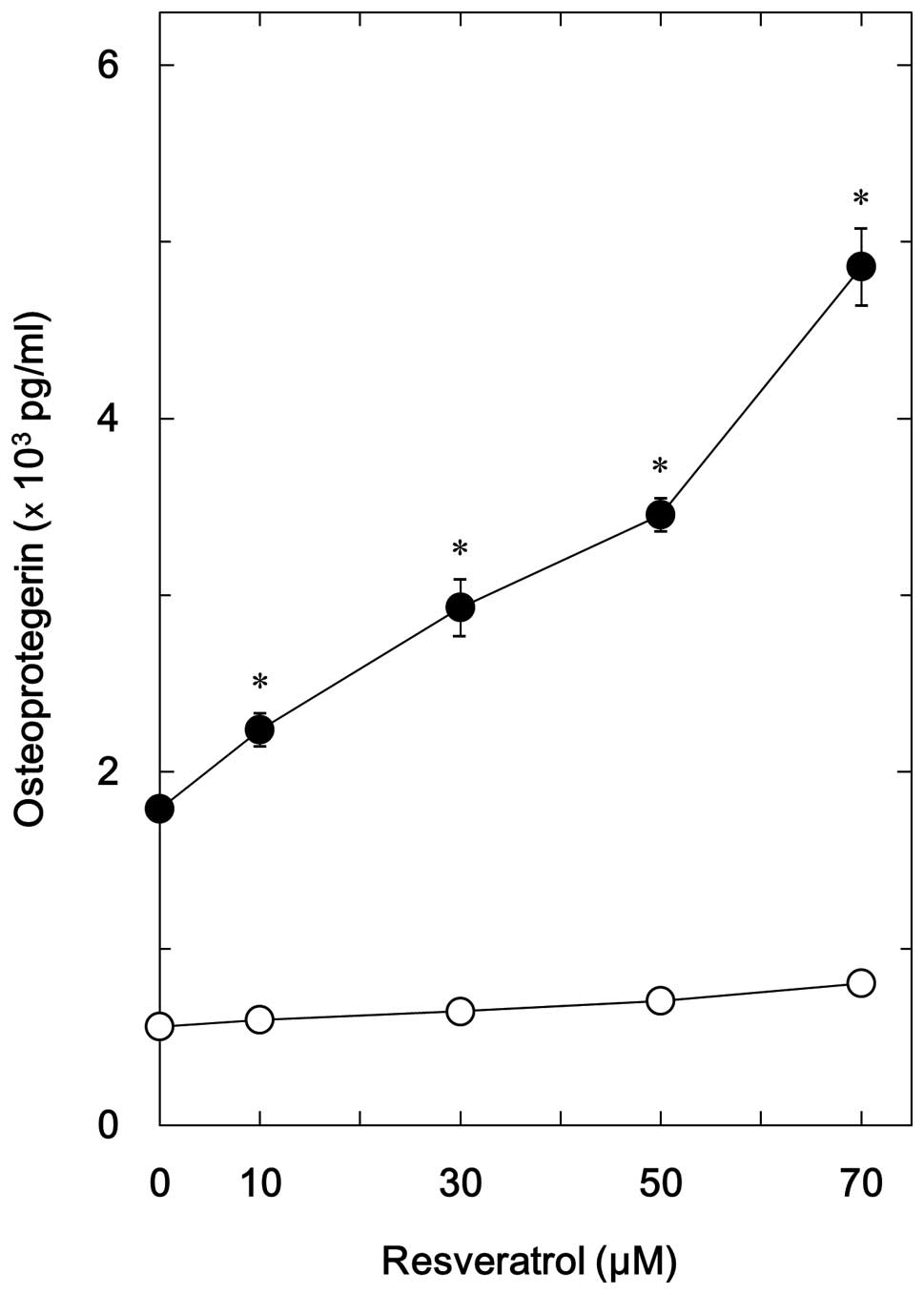
OPG (Fig. 1). The present study
also investigated the effects of resveratrol on BMP-4-stimulated
OPG release. The results suggested that although resveratrol alone
had little effect on the synthesis of OPG, it significantly
enhanced BMP-4-stimulated OPG release in a dose dependent manner
(10 70 µM) in MC3T3 E1 cells (Fig. 1). The maximum effect of resveratrol
was observed at 70 µM, which caused a ~230% increase in the
release of OPG.
Effects of resveratrol on the
BMP-4-induced mRNA expression of OPG in MC3T3-E1 cells
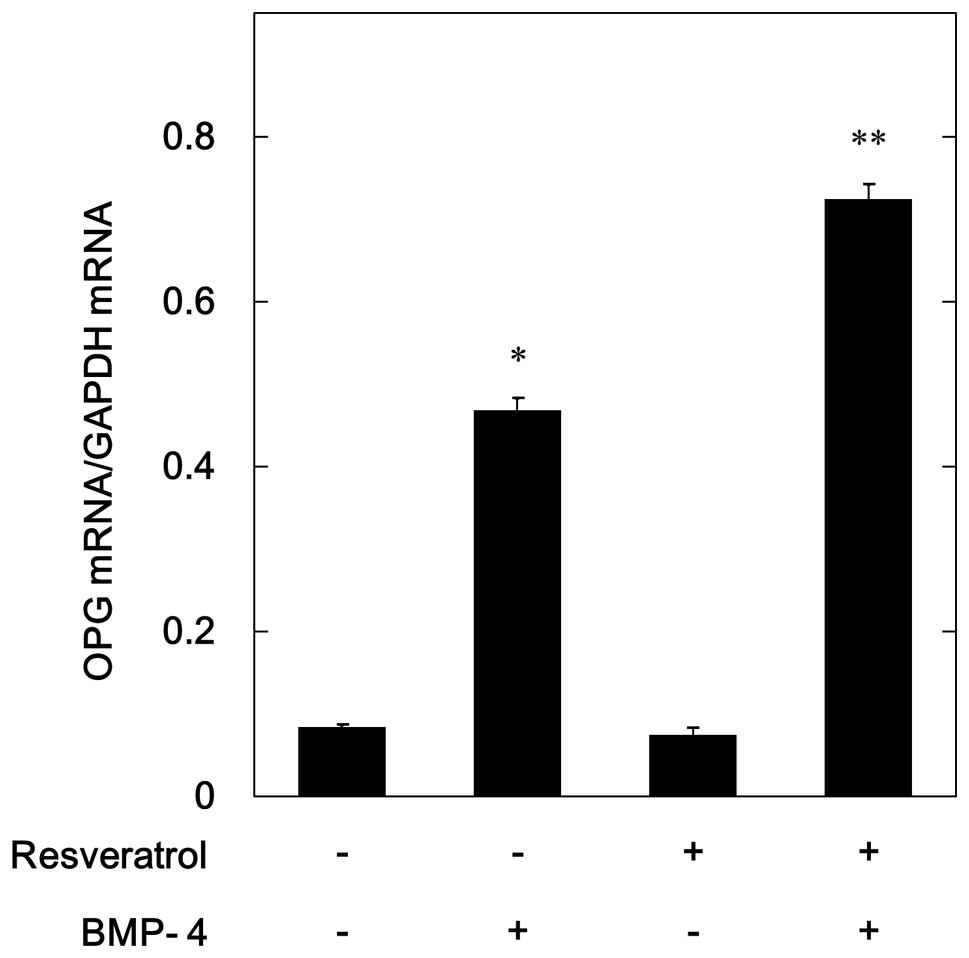
The results of the RT qPCR demonstrated that
BMP-4-induced the mRNA expression of OPG in osteoblast-like MC3T3
E1 cells (Fig. 2). In order to
investigate whether the amplifying effects of resveratrol on
BMP-4-stimulated OPG release were mediated through tran scription,
the effects of resveratrol on BMP-4-induced OPG mRNA expression
were also examined. Resveratrol alone failed to affect the mRNA
expression levels of OPG, but markedly enhanced BM-4-induced OPG
mRNA expression (Fig. 2). These
findings were concordant with the observed increase in
BMP-4-stimulated OPG release following resveratrol treatment.
Effects of SB203580 on BMP-4-stimulated
OPG release in MC3T3-E1 cells
Results from previous studies have suggested that
Smad independent signaling, such as MAP kinase signaling, mediates
the effects of BMPs (9,10). p38 MAP kinase has previously been
reported to function in BMP-4-stimulated osteocalcin synthesis as a
positive regulator in osteoblast-like MC3T3 E1 cells (11). p38 MAP kinase has also been shown
to upregulate BMP-4-stimulated VEGF synthesis in MC3T3 E1 cells
(12). In order to determine
whether p38 MAP kinase is involved in BMP-4-induced OPG synthesis
in MC3T3 E1 cells, the effects of SB203580, an inhibitor of p38 MAP
kinase (24), were examined on
BMP-4-stimulated OPG release. Treatment with SB203580 alone had
little effect on OPG release, but significantly reduced
BMP-4-stimulated OPG release in a dose dependent manner (10–30
µM) (Fig. 3).
Effects of resveratrol on BMP-4-induced
phosphorylation of p38 MAP kinase in MC3T3-E1 cells
In order to clarify whether the enhancing effects of
resveratrol on BMP-4-stimulated OPG synthesis were mediated by the
modulation of p38 MAP kinase activation in osteoblast-like MC3T3 E1
cells, the effects of resveratrol on BMP-4-induced phosphorylation
of p38 MAP kinase were examined. The results indicated that
resveratrol significantly enhanced BMP-4-induced phosphorylation of
p38 MAP kinase in a dose dependent manner (10 50 µM)
(Fig. 4). Conversely, a previous
study demonstrated that resveratrol did not affect BMP-4-induced
phosphorylation of Smad 1/5/8 in MC3T3 E1 cells (25).
Effects of SB203580 on resveratrol
enhancement of BMP-4-stimulated OPG release in MC3T3-E1 cells
The present study examined the effects of SB203580
on resveratrol enhancement of BMP-4-stimulated OPG release in
MC3T3-E1 cells. The results indicated that SB203580 signifi cantly
suppressed the resveratrol induced amplification of
BMP-4-stimulated OPG release (Table
I). SB203580 caused a ~70% decrease in the effects of
resveratrol enhanced BMP-4.
 | Table IEffects of SB203580 on the resveratrol
enhancement of BMP-4-stimulated OPG release in MC3T3-E1
osteoblastic like cells. |
Table I
Effects of SB203580 on the resveratrol
enhancement of BMP-4-stimulated OPG release in MC3T3-E1
osteoblastic like cells.
| SB203580 | Resveratrol | BMP-4 | OPG (pg/ml) |
|---|
| − | − | − | 576±17 |
| − | − | + | 2,268±46a |
| − | + | − | 632±14 |
| − | + | + | 4,610±26b |
| + | − | − | 443±31 |
| + | − | + | 948±16b |
| + | + | − | 457±20 |
| + | + | + | 1,522±41c |
Discussion
The results of the present study demonstrated that
resveratrol significantly enhanced BMP-4-stimulated OPG release in
osteoblast-like MC3T3-E1 cells. In addition, resveratrol was able
to amplify BMP-4-induced mRNA expression levels of OPG in MC3T3-E1
cells. Therefore, it is likely that the enhancing effects of
resveratrol on BMP-4-stimulated OPG release are mediated through
transcriptional events. To the best of our knowledge, this is the
first study to report the resveratrol enhancement of
BMP-4-stimulated OPG synthesis in osteoblasts.
Based on these results, the mechanism underlying
resve ratrol enhancement of BMP-4-induced OPG synthesis was
investigated in the osteoblast-like cells. Previous studies have
demonstrated that the effects of BMPs are exerted through the
intracellular signaling of both Smad-dependent, and Smad
independent pathways such as MAP kinase signaling (7,9,10).
In Smad dependent signaling, resveratrol previously failed to
affect BMP-4-induced phosphorylation of Smad 1/5/8 in
osteoblast-like MC3T3 E1 cells (25). These results suggested that it is
unlikely that the resveratrol enhance ment of BMP-4-induced OPG
synthesis is mediated through the Smad dependent pathway in
osteoblast-like MC3T3-E1 cells. Previous studies have demonstrated
that p38 MAP kinase acts as a positive regulator in the
BMP-4-stimulated synthesis of both osteocalcin and VEGF in
osteoblast-like MC3T3 E1 cells (11,12).
In addition, a previous study examined the effects of SB203580 on
BMP-4-stimulated OPG synthesis in MC3T3 E1 cells, and demonstrated
that SB203580 significantly reduced BMP-4-stimulated OPG release
(24). The results of the present
study suggested that p38 MAP kinase was also involved in
BMP-4-stimulated OPG synthesis in osteoblast-like MC3T3-E1 cells.
In addition, resveratrol was shown to markedly enhance
BMP-4-induced phosphorylation of p38 MAP kinase in MC3T3-E1 cells.
Furthermore, SB203580 significantly suppressed the resve ratrol
enhancement of BMP-4-stimulated OPG release. These results
indicated that the resveratrol amplification of BMP-4-stimulated
OPG synthesis may be mediated through the upregulation of p38 MAP
kinase activity in osteoblast-like MC3T3-E1 cells.
Resveratrol is a naturally occurring polyphenol that
is abundantly present in grapes and berries, and exhibits numerous
beneficial effects on human health, such as anti oxidation, anti
aging, and reduction of stress (26,27).
BMP-4 increases bone formation and is an important regulator of
fracture healing (8). OPG is a
decoy receptor of RANKL, which blocks RANK/RANKL interaction
thereby preventing osteoclast differentiation and activation
(4). The results of the present
study suggested that the resveratrol enhancement of BMP-4-induced
OPG synthesis led to an increase in bone formation, and a
suppression of bone resorption. Therefore, resveratrol may prove
useful in the treatment of skeletal conditions via modulation of
osteoblast function. Further investigation is required in order to
clarify the mechanism underlying resveratrol amplification of OPG
synthesis in osteoblasts.
In conclusion, the results of the present study
suggested that resveratrol was able to upregulate BMP-4-stimulated
OPG synthesis via amplification of p38 MAP kinase activity in
osteoblasts.
Acknowledgments
The authors of the present study are grateful for
the tech nical assistance of Mrs. Yumiko Kurokawa (Department of
Pharmacology, Gifu University Graduate School of Medicine). The
present study was supported by grants from the Ministry of
Education, Science, Sports and Culture of Japan (grant no.
19591042), the Foundation for Growth Science (grant no. 23–9), and
the Ministry of Health, Labour and Welfare of Japan (grant no.
25–4).
References
|
1
|
Karsenty G and Wagner EF: Reaching a
genetic and molecular understanding of skeletal development. Dev
Cell. 2:389–406. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Boyce BF, Rosenberg E, de Papp AE and
Duong le T: The osteoclast, bone remodelling and treatment of
metabolic bone disease. Eur J Clin Invest. 42:1332–1341. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Parfitt AM: Targeted and nontargeted bone
remodeling: Relationship to basic multicellular unit origination
and progression. Bone. 30:5–7. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Simonet WS, Lacey DL, Dunstan CR, Kelley
M, Chang MS, Lüthy R, Nguyen HQ, Wooden S, Bennett L and Boone T:
Osteoprotegerin: A novel secreted protein involved in the regu
lation of bone density. Cell. 89:309–319. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Mizuno A, Kanno T, Hoshi M, Shibata O,
Yano K, Fujise N, Kinosaki M, Yamaguchi K, Tsuda E, Murakami A, et
al: Transgenic mice overexpressing soluble osteoclast differen
tiation factor (sODF) exhibit severe osteoporosis. J Bone Miner
Metab. 20:337–344. 2002. View Article : Google Scholar
|
|
6
|
Kwan Tat S, Padrines M, Théoleyre S,
Heymann D and Fortun Y: IL 6, RANKL, TNF alpha/IL 1: Interrelations
in bone resorption pathophysiology. Cytokine Growth Factor Rev.
15:49–60. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Miyazono K, Maeda S and Imamura T: BMP
receptor signaling: Transcriptional targets, regulation of signals,
and signaling cross talk. Cytokine Growth Factor Rev. 16:251–263.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Krause C, de Gorter DJJ, Karperien M and
ten Dijke P: Signal trans duction cascades controlling osteoblast
differentiation. Primer on the Metabolic Bone Diseases and
Disorders of Mineral Metabolism. Rosen CJ: 7th. ASBMR; Washington:
2008, View Article : Google Scholar
|
|
9
|
Moustakas A and Heldin CH: Non Smad TGF
beta signals. J Cell Sci. 118:3573–3584. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Cai J, Pardali E, Sánchez Duffhues G and
ten Dijke P: BMP signaling in vascular diseases. FEBS Lett.
586:1993–2002. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kozawa O, Hatakeyama D and Uematsu T:
Divergent regulation by p44/p42 MAP kinase and p38 MAP kinase of
bone morpho genetic protein 4 stimulated osteocalcin synthesis in
osteoblasts. J Cell Biochem. 84:583–589. 2002. View Article : Google Scholar
|
|
12
|
Tokuda H, Hatakeyama D, Shibata T,
Akamatsu S, Oiso Y and Kozawa O: p38 MAP kinase regulates
BMP-4-stimulated VEGF synthesis via p70 S6 kinase in osteoblasts.
Am J Physiol Endocrinol Metab. 284:E1202–E1209. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hofbauer LC, Dunstan CR, Spelsberg TC,
Riggs BL and Khosla S: Osteoprotegerin production by human
osteoblast lineage cells is stimulated by vitamin D, bone morpho
genetic protein 2, and cytokines. Biochem Biophys Res Commun.
250:776–781. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Jankun J, Selman SH, Swiercz R and
Skrzypczak Jankun E: Why drinking green tea could prevent cancer.
Nature. 387:5611997. View
Article : Google Scholar : PubMed/NCBI
|
|
15
|
Harborne JB and Williams CA: Advances in
flavonoid research since 1992. Phytochemistry. 55:481–504. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Baur JA, Pearson KJ, Price NL, Jamieson
HA, Lerin C, Kalra A, Prabhu VV, Allard JS, Lopez Lluch G, Lewis K,
et al: Resveratrol improves health and survival of mice on a high
calorie diet. Nature. 444:337–342. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Renaud S and de Lorgeril M: Wine, alcohol,
platelets, and the French paradox for coronary heart disease.
Lancet. 339:1523–1526. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Mizutani K, Ikeda K, Kawai Y and Yamori Y:
Resveratrol stimulates the proliferation and differentiation of
osteoblastic MC3T3 E1 cells. Biochem Biophys Res Commun.
253:859–863. 1998. View Article : Google Scholar
|
|
19
|
Sudo H, Kodama HA, Amagai Y, Yamamoto S
and Kasai S: In vitro differentiation and calcification in a new
clonal osteogenic cell line derived from newborn mouse calvaria. J
Cell Biol. 96:191–198. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kozawa O, Tokuda H, Miwa M, Kotoyori J and
Oiso Y: Cross talk regulation between cyclic AMP production and
phosphoinositide hydrolysis induced by prostaglandin E2 in
osteoblast-like cells. Exp Cell Res. 198:130–134. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Laemmli UK: Cleavage of structural
proteins during the assembly of the head of bacteriophage T4.
Nature. 227:680–685. 1970. View
Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kato K, Ito H, Hasegawa K, Inaguma Y,
Kozawa O and Asano T: Modulation of the stress induced synthesis of
hsp27 and alpha B crystallin by cyclic AMP in C6 rat glioma cells.
J Neurochem. 66:946–950. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Simpson DA, Feeney S, Boyle C and Stitt
AW: Retinal VEGF mRNA measured by SYBR green I fluorescence: A
versatile approach to quantitative PCR. Mol Vis. 6:178–183.
2000.PubMed/NCBI
|
|
24
|
Cuenda A, Rouse J, Doza YN, Meier R, Cohen
P, Gallagher TF, Young PR and Lee JC: SB 203580 is a specific
inhibitor of a MAP kinase homologue which is stimulated by cellular
stresses and interleukin 1. FEBS Lett. 364:229–233. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kondo A, Otsuka T, Kuroyanagi G, Yamamoto
N, Matsushima Nishiwaki R, Mizutani J, Kozawa O and Tokuda H:
Resveratrol inhibits BMP-4-stimulated VEGF synthesis in osteo
blasts: Suppression of S6 kinase. Int J Mol Med. 33:1013–1018.
2014.PubMed/NCBI
|
|
26
|
Blander G and Guarente L: The Sir2 family
of protein deacetylases. Annu Rev Biochem. 73:417–435. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Koo SH and Montminy M: In vino veritas: a
tale of two sirt1s? Cell. 127:1091–1093. 2006. View Article : Google Scholar : PubMed/NCBI
|