Introduction
Alzheimer's disease (AD) is a common, age-associated
neurodegenerative disorder, which is characterized by a loss of
synapses and neurons, intracellular neurofibrillary tangles and the
formation of extracellular amyloid plaques (1). The classical hypothesis for the cause
of AD is the aberrant amyloid protein deposition of amyloid β
(Aβ)42 (2). The etiology of AD is
complex and is composed of genetic and environmental factors
(3,4). There is evidence indicating that
dysfunctions of neuro-immune networks also contribute to the
pathogenesis of AD (5,6).
Increasing evidence suggests that microglia are
important in the pathophysiology of AD, and Toll-like receptor
(TLR)2 is one of the pattern recognition receptors expressed in
microglia, which was originally identified based on their response
to invading microorganisms (7). In
mouse models of AD, microglia are activated and recruited to the
deposits of pathogenic Aβ, where they subsequently damage neurons
(8–10). Deficiencies in TLR2 and TLR4 in
cultured microglia are significantly reduced following Aβ-triggered
inflammatory activation (11,12).
TLR2 knockout in a mouse model of AD decreased the deposition of
cerebral Aβ (13). Additionally,
overexpression of TLR2 has been observed in patients with AD and
demonstrates that TLR2 is important in Aβ-triggered inflammatory
activation and Aβ phagocytosis (14).
MicroRNAs (miRs) are a group of negative regulators
of gene expression, which repress gene expression by directly
binding the 3′-untranslated region (UTR) of mRNA. miRs are
important in maintaining normal physiological conditions in the
human body, and abnormal expression of miR has been associated with
several human diseases, including psychiatric disorders and certain
types of malignant cancer (15–17).
The dysfunction of miR in neurodegenerative disorders is becoming
increasingly recognized. miR-146a has been demonstrated to be
important in AD, with aberrant expression of miR-146a having been
identified in transgenic mouse models of AD and in human AD brain
cells (18,19). Therefore, the present study aimed
to investigate whether single nucleotide polymorphisms (SNPs) or
mutations in the miR-146a coding region were associated with the
pathogenesis of AD.
Primary (pri)-miR-146a was sequenced in the genomic
DNA of 103 patients with AD. The effect of a polymorphism site on
the expression of mature miR-146a was examined and a novel miR-146a
target gene was confirmed. The biological function of this site on
miR-146a target genes expression and the immune response to Aβ42
was also investigated.
Materials and methods
DNA collection and genotyping
Venous blood samples from 103 patients with AD (age
range, 56–85 years; mean ± standard deviation, 68.32±8.12) and 206
healthy individuals (54–88 years; 69. 03 ±7.43) were obtained from
Shanghai Fengxian Central Hospital (Shanghai, China). All the
patients were from the Han population and their ancestries were
from the North China plain area (Beijing, Shandong and Hebei
province). The study was approved by the ethics committee of the
Department of Neurology, Shanghai Fengxian Central Hospital.
Written informed consent was obtained from the patient's
family.
Venous blood (5 ml) was collected from patient and
control individuals upon their first admission into the hospital.
To harvest cell-free serum, the blood was drawn into sterile tubes
(BD Biosciences, Franklin Lakes, NJ, USA) without anticoagulant and
left in a standing position for 20 min. The samples were
centrifuged at 1,500 x g for 10 min at 20°C and the supernatant
serum was removed and stored immediately at -80°C, until further
analysis.
The DNA from the blood samples was extracted using a
TIANamp Blood DNA kit (Tiangen, Beijing, China). The DNA specimens
were amplified by using standard PCR techniques. A total of 50 ng
DNA from each sample was used for the PCR reaction. The Pfu DNA
polymerase was purchased from Tiangen. The cycling conditions were
as follows: 95°C for 10 min; 95°C for 30 sec followed by 60°C for 1
min for 30 cycles. The primer sequences were as follows: Forward,
5′-GGTCTCCTCCAGATGTTTATAACTC-3′ and reverse
5′-GAACCCTGCTTAGCATAGAATTC-3′. The PCR products were sequenced in
forward direction with the ABI 3730xl sequencing platform (Applied
Biosystems, Foster City, CA, USA) by BGI-GBI Biotech Co., Ltd.
(Beijing, China). The sequencing results were analyzed using DNAMAN
version 7,0,2,176 (Lynnon Corporation, San Ramon, CA, USA) and
Chromas Lite version 2.22 (Technelysium Pty Ltd., South Brisbane,
Queensland, Australia) software.
Cell culture
The RAW264.7, A549 and HEK293T cells (China
Infrastructure of Cell Line Resources, Beijing, China) were
cultured in Dulbecco's modified Eagle's medium (Corning
Incorporated, Corning, NY, USA) containing 10% fetal bovine serum
(Hyclone, Logan, UT, USA), 100 U/ml penicillin and 10 mg/ml
streptomycin (Hyclone). All the cells were maintained at 37°C in an
atmosphere of 5% CO2.
miR-146a expression vectors
To construct the miR-146a expression vectors,
fragments (432 nt) corresponding to the pri-miR-146a and its
flanking regions (previously determined to have the two genotypes)
were amplified from the cDNA and cloned into the pcDNA3.1 vector
(Invitrogen Life Technologies, Carlsbad, CA, USA). The sequences of
the vectors were confirmed by direct sequencing and the only
difference was in the mutation site. The miR-146a expression
vectors were transfected into the cells using Lipofectamine 2000
(Invitrogen Life Technologies), according to the manufacturer's
instructions. An empty pcDNA3.1 vector was used as a control.
RT-qPCR
RT-qPCR analysis was used to determine the relative
expression of miR-146a-5p. The total RNA was extracted from cells
using TRIzol reagent (Invitrogen Life Technolgies) according to the
manufacturer's instructions. The expression of miR-146a-5p was
detected by TaqMan miRNA RT-Real Time PCR (Applied Biosystems).
Single-stranded cDNA was synthesized using a TaqMan MicroRNA
Reverse Transcription kit (Applied Biosystems) and then amplified
using TaqMan Universal PCR Master mix (Applied Biosystems) together
with miRNA-specific TaqMan MGB probes (Applied Biosystems)
targeting miR-146a-5p (Applied Biosystems). The U6 small nuclear
RNA (Applied Biosystems) was used for normalization. The
experiments were performed using the ABI 7300 PCR thermal cycler
(Applied Biosystems). The cycling conditions for the qPCR were as
follows: 95°C for 10 min, 95°C for 15 sec followed by 60°C 1 min
for 40 cycles. The samples in each group were measured in
triplicate and the experiment was repeated at least three times for
the detection of miR-146a-5p. For serum miR-146a detection, miR-16
(Applied Biosystems) was used for normalization. Total RNA was
extracted from a 200 µl serum sample from each participant
using TRIzol LS reagent (Invitrogen Life Technologies) according to
the manufacturer's instructions. The serum miR-146a and miR-16
level was detected using a specific primer and probe (Applied
Biosystems) as described above.
Dual luciferase assay
To generate a luciferase reporter vectors,
full-length TLR2 3′-UTR (828 bp) was cloned downstream of the
firefly luciferase coding region in the pmirGLO vector (Promega
Corporation, Madison, WI, USA). The sequences of primers for TLR2
3′-UTR amplification were as follows: Forward,
5′-CTCGAGGTTCCCATATTTAAGACCAG-3′ and reverse,
5′-TCTAGATTCTCATCCTGTAAAGTTTAA TAGG-3′. The PCR cycling conditions
were as follows: 95°C for 10 min, 95°C for 15 sec, 61°C for 30 sec,
then 72°C for 1 min. A total of 50 ng genomic DNA was used for each
PCR reaction. For the luciferase reporter assays, the HEK293T cells
were seeded into 48-well plates (Corning Incorporated) at a density
of 3×104 cells/well. miR-146a expression vector,
miR-146a mimic or miR-146a inhibitor (GenePharma Co., Ltd.,
Shanghai, China) were co-transfected with a luciferase reporter
vector using lipofectamine 2000 (Invitrogen Life Technologies).
After incubation for 4 h at 37°C, the medium was discarded and
replaced with fresh DMEM medium for a further 48 h culture. The
cells were harvested and assayed using a Dual-Luciferase assay
(Promega Corporation). Each treatment was performed in triplicate
in three independent experiments. The data are expressed as the
relative luciferase activity (firefly luciferase/Renilla
luciferase).
Western blotting
The cells were collected by centrifugation at 1,000
x g and then resuspended in lysis buffer for 10 min to ensure
complete lysis. The cells were centrifuged at 12,000 x g for 10
minutes at 4°C and the supernatant was harvested into a fresh tube.
The protein quantities were detected using a bicinchoninic protein
assay kit, according to the manufacturer's instructions. The
protein extracts were boiled in 50 µl 2X
SDS/β-mercaptoethanol sample buffer (Sigma-Aldrich) and 30
µg samples were loaded into each lane of 8% polyacrylamide
gels (Sigma-Aldrich). The proteins were separated by
electrophoresis and were subsequently blotted onto polyvinylidene
fluoride membranes (Amersham Pharmacia Biotech, St. Albans, UK) by
electrophoretic transfer. The membrane was incubated with rabbit
anti-TLR2 monoclonal antibody (Abcam, Cambridge, MA, USA; cat. no.
ab108998; 1:1,000) or mouse anti-β-actin monoclonal antibody (Santa
Cruz Biotechnology, Inc., Santa Cruz, CA, USA; cat. no. sc-58673;
1:1,000) for 1 h at 37°C. The specific protein-antibody complex was
detected using horseradish peroxidase-conjugated goat anti-rabbit
(Santa Cruz Biotechnology Inc.; cat. no. sc-2004; 1:5,000) or
rabbit anti-mouse IgG (Santa Cruz Biotechnology Inc.; cat. no.
sc-358920; 1:5,000). Detection of a chemiluminescence reaction was
performed using an enhanced chemiluminescence kit (Pierce
Biotechnology). The β-actin signal was used as a loading
control.
Aß42 challenge and ELISA detection of
tumor necrosis factor (TNF)-α
The RAW264.7 cells (2×105/well) were
plated into 48-well plates and treated with 10 µM aggregated
Aβ42 (AnaSpec, Fremont, CA, USA) for 24 h. The supernatants were
collected for the detection of TNF-α using a mouse TNF-α instant
ELISA kit (eBioscience, San Diego, CA, USA).
Statistical analysis
The data were analyzed using SPSS 16.0 statistical
software (SPSS, Inc., Chicago, IL, USA). Statistical significance
was determined using Student's t-test for two-group analyses and a
Mann-Whitney U test for the tissue levels of miR-146a. The results
of the genotype frequency and polymorphism distribution were
analyzed using the χ2 test. P<0.05 was considered to
indicate a statistically significant difference.
Results
Genotypes and risk of AD
Precursor (pre)-miR-146a is a 99 nt RNA segment and
four single nucleotide polymorphisms (SNPs) exist in this region,
according to the dbSNP database (build 137; http://www.ncbi.nlm.nih.gov/). To investigate whether
there is an association between the nucleotide variants of
pre-miR-146 and the pathogenesis of AD, the coding region of
pri-miR-146a in 103 patients with AD and in 206 healthy individuals
was scanned. Although no novel sequence alterations were detected,
the rare C allele of rs2910164 was found to be associated with AD
(OR=1.50, 95% CI=1.03–2.17; Table
I).
 | Table IGenotype frequencies of rs2910164 in
patients and controls and their association with Alzheimer's
disease. |
Table I
Genotype frequencies of rs2910164 in
patients and controls and their association with Alzheimer's
disease.
| Genotype | Patient (n=103) n
(%) | Control (n=206) n
(%) | Odds ratio (95%
CI) | P-value |
|---|
| C | 65 (31.550) | 97 (0.240) | 1.50 (1.030,
2.170) | 0.030 |
| G | 141 (68.450) | 315 (0.76) | 0.67 (0.460,
0.970) | |
| CC | 8 (0.078) | 4 (0.0190) | 4.25 (1.250,
14.47) | |
| CG | 49 (0.480) | 89 (0.430) | 1.19 (0.740,
1.920) | 0.004 |
| GG | 46 (0.450) | 113 (0.550) | 0.66 (0.410,
1.070) | |
C allele reduces the expression of
miR-146a in vitro and in vivo
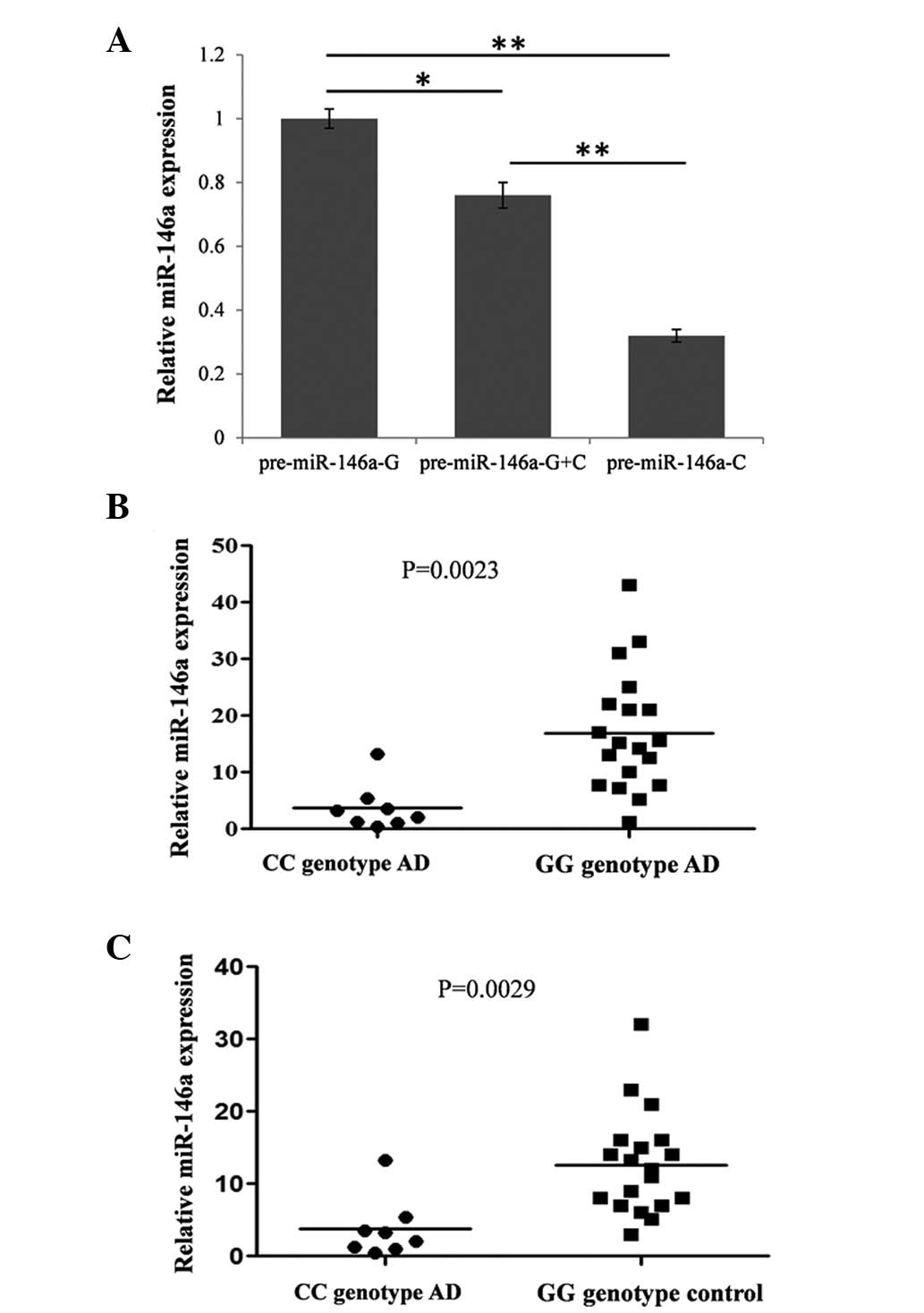
Previous studies have reported that rs2910164 within
the pre-miR-146a sequence reduces the levels of pri- and mature
miR-146a from the C allele, compared with the allele G (1.9- and
1.8-fold, respectively) (20). The
present study detected the levels of mature miR-146a in different
genotypic pri-miR-146a expression vectors in transiently
transfected HEK293T cells by RT-qPCR. As expected, the C allele
reduced the expression of mature miR-146a to 32% compared with
allele G (Fig. 1A).
To understand whether this SNP affected the
expression of miR-146a in vivo, the expression of miR-146a
was compared between the GG and CC genotypes in the serum of
patients with AD and healthy controls by RT-qPCR. The expression of
miR-16 was used as an internal control. The results demonstrated
that the expression of miR-146a in the serum of CC genotype
patients with AD was significantly downregulated compared with the
GG genotype patients with AD (Fig.
1B) and the GG genotype healthy individuals (Fig. 1C).
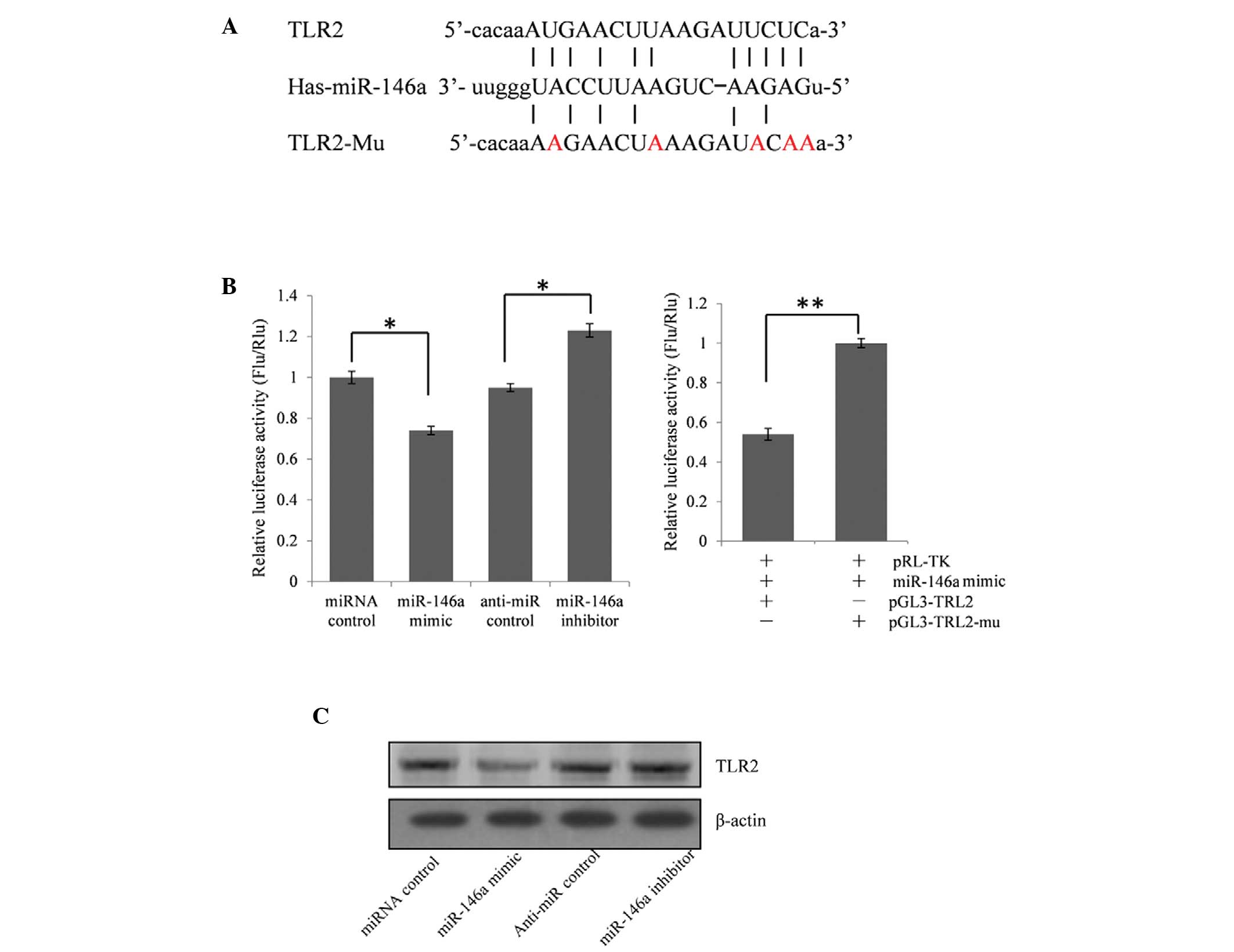
Expression of TLR2 is repressed by
miR-146a
The function of miRs are predominantly reflected in
the repression effect on their target genes. To investigate the
effect of a reduced expression of miR-146a on the pathogenesis of
AD, miR-146a target genes were predicted using the online
bioinformatics tool, miRanda (21). This identified that TLR2, the
upregulation of which is associated with triggering
neuro-inflammatory activation and the pathogenesis of AD, may be a
target gene of miR-146a (7).
To validate whether TLR2 is an miR-146a target gene,
the full-length 828 bp segment of TLR2 3′-UTR was cloned downstream
of the firefly luciferase reporter gene in the pGL3 control vector
(designated as pGL3-TLR2) for the dual luciferase assay. The
HEK293T cells were co-transfected with pGL3-TLR2 and a miR-146a
mimic or inhibitor (Fig. 2B). The
luciferase activity was reduced significantly by ~26.0% (P<0.05)
in the presence of miR-146a, compared with the control.
Furthermore, the luciferase activity was significantly upregulated
by ~29.5% (P<0.05) following treatment with the miR-146a
inhibitor, compared with the anti-miR control. These results
indicated that miR-146a targeted the 3′-UTR of TLR2, leading to the
change of firefly luciferase translation.
A seed sequence mutation clone was used to further
confirm the binding site for miR-146a (Fig. 2A). The vector contains putative
miR-146a binding regions in the 3′-UTR of TLR2, with five mutant
nucleotides (designated as pGL3-TLR2-Mu). This vector was used and
the wild-type TRL2 vector was used as a control. The histogram
(Fig. 2B) shows that the enzyme
activity was increased ~46.1% in cells co-transfected with the
miR-146a mimics and pGL3-TLR2-Mu compared with pGL3-TLR2
(P<0.01). These data indicated that miR-146a suppressed the
expression of TLR2 through binding to the seed sequence at the
3′-UTR of TLR2, and TLR2 may be a direct target of miR-146a.
miR-146a regulates the endogenous
expression of TLR2 in A549 cells
As TLR2 was identified as a target gene for
miR-146a, whether miR-146a regulated the endogenous expression of
TLR2 was examined. The A549 cells were transfected with either an
miR-146a mimic or an inhibitor to determine whether the
dysregulation of the expression of miR-146a affected the endogenous
expression of TLR2. Compared with the corresponding control, the
protein expression of TLR2 was significantly suppressed by the
miR-146a mimic and was upregulated by the miR-146a inhibitor
(Fig. 2C).
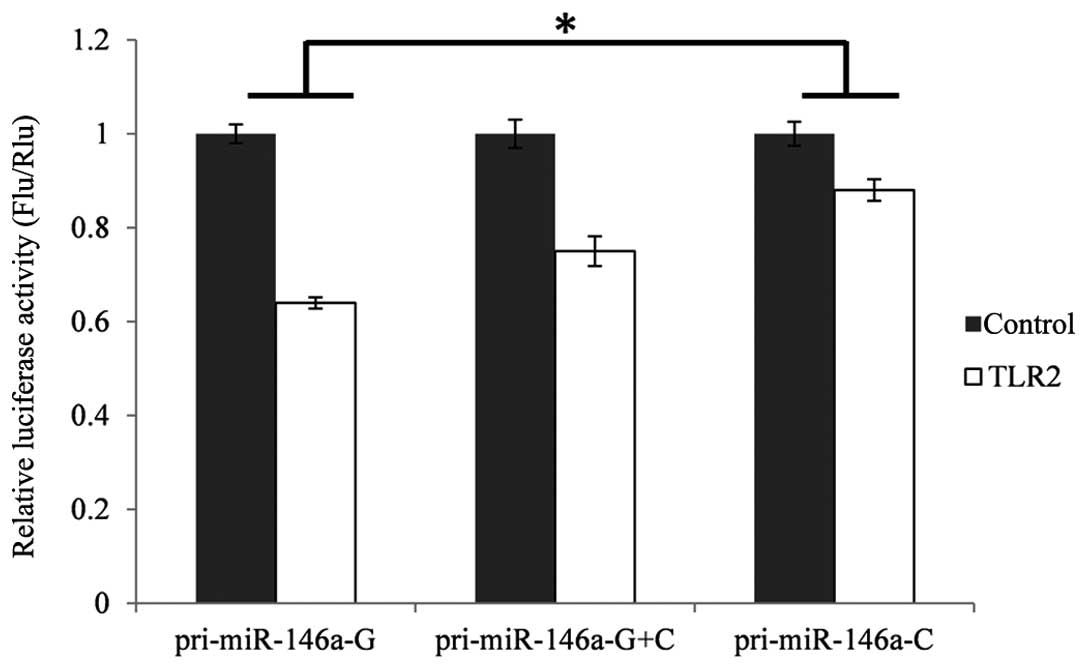
Impact of rare C allele on the expression
of TLR2
To investigate the functional consequences of
disturbed expression of miR-146a on its target genes, the TLR2
3′-UTR dual luciferase assay system was used. These reporter
constructs were transiently transfected into the HEK293T cells,
together with an expression plasmid containing the pri-miR-146a of
either genotype. The results were analyzed using multiple
comparison/post-hoc tests of analysis of variance (ANOVA) Levene's
test was used to assess the variance in homogeneity, which is a
pre-condition for parametric tests, including t-tests and ANOVA.
The results revealed that the variances were homogeneous in TLR2
(P=0.24). As shown in Fig. 3, the
activity of firefly luciferase was separately decreased by 23.9%
(P<0.05) in the cells co-transfected with TLR2 and
pcDNA3.1-miR146a-GG or pcDNA3.1-miR-146a-CC, compared with the
control.
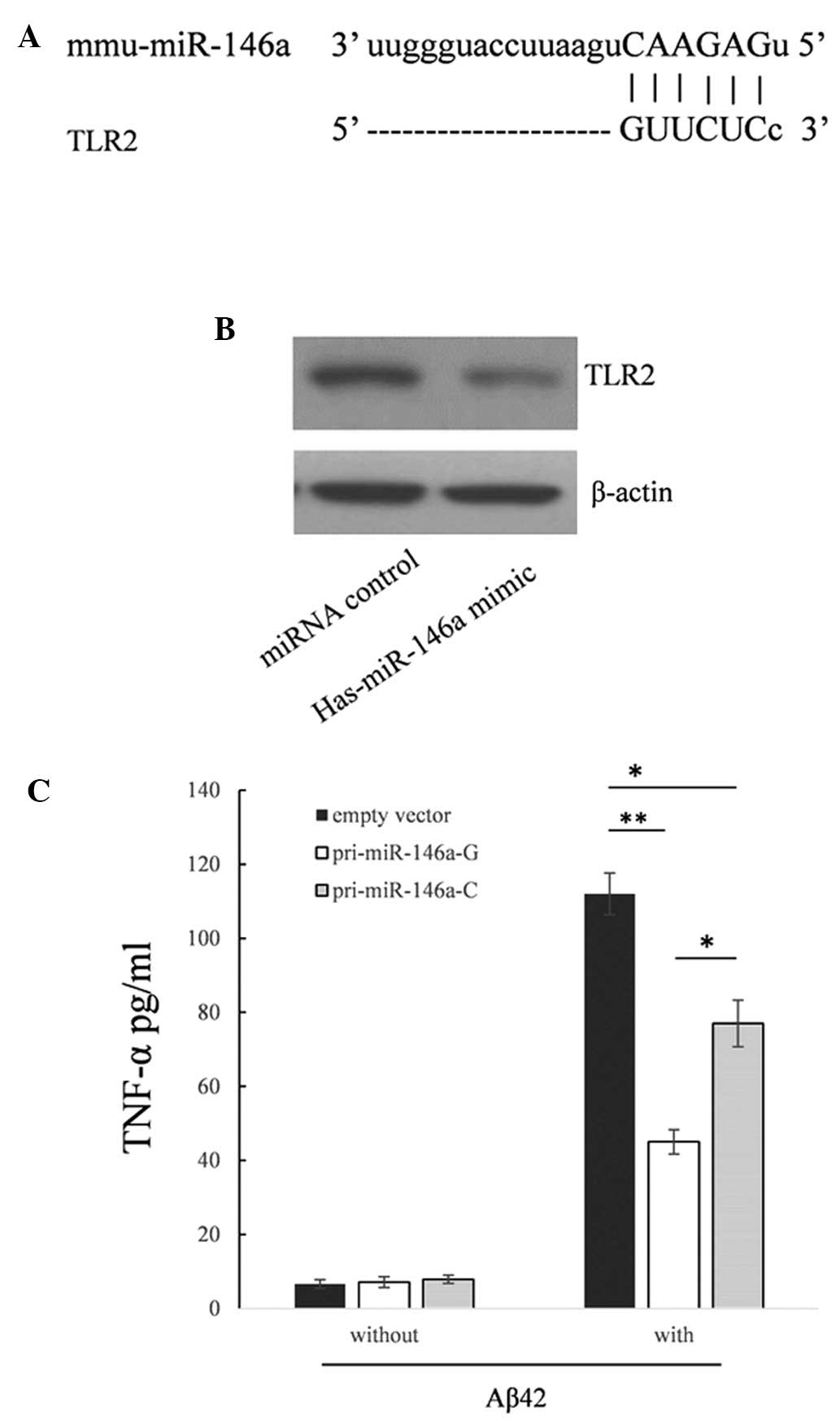
Rare C allele in pri-miR-146a upregulates
the production of TNF-α in RAW264.7 cells following stimulation
with Aß42
Since the expression of TLR2 was repressed in the
RAW264.7 cells, which are derived from mice (Fig. 4A and B), and the sequences of human
miR-146a and mouse miR-146a are identical, the present study
detected the biological function of the rare C allele in this cell
line. As shown in Fig. 4C, the
supernatant levels of TNF-α were reduced by 59.8% (P<0.01)
compared with the cells transfected with the empty vector. When
transfected with the pri-miR-146a-C vector, the supernatant levels
of TNF-α were raised by 71.1% (P<0.05) compared with the
pri-miR-146a-C.
Discussion
The innate immune response and inflammatory
signaling are critical for brain homeostasis, neuroprotection and
repair (22). If these are
overactivated, they produce excess oxygen free radicals,
pro-inflammatory cytokines and prostaglandins, subsequently
triggering an inflammatory cascade, resulting in neurodegeneration
(23).
miR-146a is a negative feedback regulator of the
innate immune system and may be important for controlling TLR and
cytokine signaling. High expression levels of miR-146a have been
identified in various inflammatory diseases, including rheumatoid
arthritis (24) and psoriasis
(25). Previous investigations
have observed that the expression of miR-146a is upregulated in the
brain of a mouse model of AD and in the brain tissue of patients
with AD (18,19,26).
These findings indicated that the overexpression of miR-146a may be
used to evaluate the presence and quantify the degree of
inflammation in a patient without infection, and may be used as a
diagnostic marker for AD. The present study identified the rare C
allele of rs2910164, which reduced the expression of miR-146a
associated with AD in the Chinese-Han population. This reduced the
repressive effect on the expression of its target genes, including
TLR2. TLR2 has been confirmed to interact with Aβ42 and is a
primary receptor for Aβ42. The roles of TLR2 in Aβ-triggered
inflammatory activation and Aβ phagocytosis have been partially
elucidated (7). The present study
demonstrated that downregulation of miR-146a may be involved in the
pathogenesis of AD. Reduced expression of miR-146a may weaken the
negative feedback regulation of the inflammatory reaction and
increase tissue damage by upregulating the expression of TLR2
during the pathogenesis of AD.
In conclusion, the present study established the
first, to the best of our knowledge, association between a
polymorphism site and the risk of developing AD in one Chinese-Han
population. These findings provide insight into understanding the
development of AD and offer a potential approach in the diagnosis
and treatment of AD.
References
|
1
|
Van den Hove DL, Kompotis K, Lardenoije R,
Kenis G, Mill J, Steinbusch HW, Lesch KP, Fitzsimons CP, De
Strooper B and Rutten BP: Epigenetically regulated microRNAs in
Alzheimer's disease. Neurobiol Aging. 35:731–745. 2014. View Article : Google Scholar
|
|
2
|
Schenk D, Barbour R, Dunn W, Gordon G,
Grajeda H, Guido T, Hu K, Huang J, Johnson-Wood K, Khan K, et al:
Immunization with amyloid-beta attenuates Alzheimer-disease-like
pathology in the PDAPP mouse. Nature. 400:173–177. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Machado A, Herrera AJ, de Pablos RM,
Espinosa-Oliva AM, Sarmiento M, Ayala A, Venero JL, Santiago M,
Villarán RF, Delgado-Cortés MJ, et al: Chronic stress as a risk
factor for Alzheimer's disease. Rev Neurosci. 25:785–804. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Karch CM, Cruchaga C and Goate AM:
Alzheimer's disease genetics: From the bench to the clinic. Neuron.
83:11–26. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Morales I, Farías G and Maccioni RB:
Neuroimmunomodulation in the pathogenesis of Alzheimer's disease.
Neuroimmunomodulation. 17:202–204. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hickman SE and El Khoury J: The
neuroimmune system in Alzheimer's disease: The glass is half full.
J Alzheimers Dis. 33(Suppl 1): S295–S302. 2013.
|
|
7
|
Liu S, Liu Y, Hao W, Wolf L, Kiliaan AJ,
Penke B, Rübe CE, Walter J, Heneka MT, Hartmann T, et al: TLR2 is a
primary receptor for Alzheimer's amyloid β peptide to trigger
neuroinflammatory activation. J Immunol. 188:1098–1107. 2012.
View Article : Google Scholar
|
|
8
|
Masters CL and Beyreuther K: Alzheimer's
centennial legacy: prospects for rational therapeutic intervention
targeting the Abeta amyloid pathway. Brain. 129:2823–2839. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Bolmont T, Haiss F, Eicke D, Radde R,
Mathis CA, Klunk WE, Kohsaka S, Jucker M and Calhoun ME: Dynamics
of the microglial/amyloid interaction indicate a role in plaque
main tenance. J Neurosci. 28:4283–4292. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Meyer-Luehmann M, Spires-Jones TL, Prada
C, Garcia-Alloza M, de Calignon A, Rozkalne A, Koenigsknecht-Talboo
J, Holtzman DM, Bacskai BJ and Hyman BT: Rapid appearance and local
toxicity of amyloid-beta plaques in a mouse model of Alzheimer's
disease. Nature. 451:720–724. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Jana M, Palencia CA and Pahan K: Fibrillar
amyloid-beta peptides activate microglia via TLR2: Implications for
Alzheimer's disease. J Immunol. 181:7254–7262. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Udan ML, Ajit D, Crouse NR and Nichols MR:
Toll-like receptors 2 and 4 mediate Abeta (1–42) activation of the
innate immune response in a human monocytic cell line. J Neurochem.
104:524–533. 2008.
|
|
13
|
Richard KL, Filali M, Préfontaine P and
Rivest S: Toll-like receptor 2 acts as a natural innate immune
receptor to clear amyloid beta 1–42 and delay the cognitive decline
in a mouse model of Alzheimer's disease. J Neurosci. 28:5784–5793.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhang W, Wang LZ, Yu JT, Chi ZF and Tan L:
Increased expressions of TLR2 and TLR4 on peripheral blood
mononuclear cells from patients with Alzheimer's disease. J Neurol
Sci. 315:67–71. 2012. View Article : Google Scholar
|
|
15
|
Maes OC, Chertkow HM, Wang E and Schipper
HM: MicroRNA: Implications for Alzheimer Disease and other Human
CNS Disorders. Curr Genomics. 10:154–168. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Xu J, Li Y, Wang F, Wang X and Cheng B:
Suppressed miR-424 expression via upregulation of target gene Chk1
contributes to the progression of cervical cancer. Oncogene.
32:976–987. 2013. View Article : Google Scholar
|
|
17
|
Farazi TA, Hoell JI, Morozov P and Tuschl
T: MicroRNAs in human cancer. Adv Exp Med Biol. 774:1–20. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lukiw WJ, Zhao Y and Cui JG: An
NF-kappaB-sensitive micro RNA-146a-mediated inflammatory circuit in
Alzheimer disease and in stressed human brain cells. J Biol Chem.
283:31315–31322. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li YY, Cui JG, Hill JM, Bhattacharjee S,
Zhao Y and Lukiw WJ: Increased expression of miRNA-146a in
Alzheimer's disease transgenic mouse models. Neurosci Lett.
487:94–98. 2011. View Article : Google Scholar
|
|
20
|
Jazdzewski K, Murray EL, Franssila K,
Jarzab B, Schoenberg DR and de la Chapelle A: Common SNP in
pre-miR-146a decreases mature miR expression and predisposes to
papillary thyroid carcinoma. Proc Natl Acad Sci USA. 105:7269–7274.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Betel D, Wilson M, Gabow A, Marks DS and
Sander C: The microRNA.org resource: Targets and expression.
Nucleic Acids Res. 36(Database): D149–D153. 2008. View Article : Google Scholar :
|
|
22
|
Ransohoff RM and Brown MA: Innate immunity
in the central nervous system. J Clin Invest. 122:1164–1171. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lynch MA: The impact of neuroimmune
changes on development of amyloid pathology; relevance to
Alzheimer's disease. Immunology. 141:292–301. 2014. View Article : Google Scholar :
|
|
24
|
Nakasa T, Miyaki S, Okubo A, Hashimoto M,
Nishida K, Ochi M and Asahara H: Expression of microRNA-146 in
rheumatoid arthritis synovial tissue. Arthritis Rheum.
58:1284–1292. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sonkoly E, Wei T, Janson PC, Sääf A,
Lundeberg L, Tengvall-Linder M, Norstedt G, Alenius H, Homey B,
Scheynius A, et al: MicroRNAs: Novel regulators involved in the
pathogenesis of psoriasis? PLoS One. 2:e6102007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Sethi P and Lukiw WJ: Micro-RNA abundance
and stability in human brain: Specific alterations in Alzheimer's
disease temporal lobe neocortex. Neurosci Lett. 459:100–104. 2009.
View Article : Google Scholar : PubMed/NCBI
|