Introduction
Colorectal cancer (CRC) is a major cause of
morbidity and mortality. It is the third most common type of
malignant cancer worldwide, and the fourth leading cause of
cancer-associated mortality (1).
The incidence of CRC is ranked only third to lung and breast cancer
(2). Furthermore, it has been
estimated that >143,460 new CRC cases were diagnosed in 2012,
and the number of patients with CRC worldwide are expected to reach
9,000,000 by 2020 (3,4).
Despite progress in surgical and nonsurgical
treatment, the prognosis for CRC remains poor; this is partially
due to CRC usually being diagnosed at an advanced stage, when
curative therapy is not possible. Therefore, earlier detection,
diagnosis and prevention are key factors in controlling and
treating CRC (5). It is well known
that CRC develops as the result of a series of genetic and
epigenetic alterations, which lead to the transformation of normal
colorectal epithelium into colorectal adenocarcinoma (6). Identifying these key genetic
alterations is critical for improving current understanding of CRC
and for improving treatment strategies.
Hematopoietic cell-specific protein 1
(HS-1)-associated protein X-1 (HAX-1) was first identified as a 35
kDa protein, which interacts with HS-1, an Src kinase substrate.
HAX-1 is composed of a putative transmembrane domain, a putative
PEST sequence and an acid box (7).
HAX-1 has emerged as an important factor in the intrinsic
mitochondria-dependent pathway of cell death, which is
characterized by the activation and permeabilization of
mitochondria, resulting in the release of cytochrome c and
other pro-apoptotic molecules into the cytosol (8). Previous studies have demonstrated
that HAX-1 is involved in the regulation of mitochondrial membrane
potential during apoptosis (9,10).
In addition, HAX-1 protein interacts with a number of cellular and
viral proteins (11–13). However, the function of HAX-1
remains to be fully elucidated, particularly in CRC. Several
studies have demonstrated that HAX-1 may be important in apoptosis
and proliferation (14,15).
In our previous study, yeast two-hybrid screening
demonstrated that HAX-1 had a key role in the Salvador-warts-hippo
signaling pathway, which is an important supplement for the classic
apoptosis pathway (16,17). The present study aimed to analyze
the potential prognostic significance of HAX-1 in CRC by detecting
its expression levels in human colorectal tumor tissues, and to
assess the role of HAX-1 in apoptosis and proliferation using
gene-overexpression and silencing methods.
Materials and methods
Patients
The present study analyzed the expression levels of
HAX-1 in tumor tissues from 60 patients with CRC, who were
diagnosed and underwent elective surgery at the Gastrointestinal
Surgery Center, Tongji Hospital (Wuhan, China) between February
2010 and November 2011. A total of 40 patients (66.7%) were
>60-years old at diagnosis. The tissue samples (~2×2 cm) were
collected in surgery, and were cut into three sections, one for
protein extraction, one for RNA extraction and one for
immunohistochemistry. The clinical stages of the CRC tissue samples
were graded according to Dukes' Staging System (10). A total of 10 patients were
classified as stage A, 26 patients as stage B, 14 patients as stage
C and 10 patients as stage D. The current study was approved by the
Ethics Committee of Tongji Hospital, Huazhong University of Science
and Technology (Wuhan, China).
Animals
Athymic female nude mice (4–6 weeks old, 15–20 g)
were obtained from the Shanghai Laboratory Animal Center (Shanghai,
China). Mice were fed under specific pathogen free conditions in a
temperature- and humidity-controlled environment. All animal
experiments were in accordance with the Institutional Animal
Research Guidelines approved by the Ethics Committee. For
tumor-growth studies, a total of 5×105 SW48-shcon or
SW480 KD1 cells were injected subcutaneously into each mouse (5
mice/group).
Colorectal tumor tissue and cell
lines
Samples of fresh tumor tissue were obtained from 60
patients with CRC and available for investigation. The tumor
samples were obtained at the time of diagnosis in all cases.
Institutional review board-approved, informed consent was obtained
from the patients. All specimens were coded by the admission of the
patient prior to analysis. The SW480 human CRC cell line and HEK293
human embryonic kidney cell line (used as a lentivirus packaging
cell line) were obtained from Peking Tissue Type Culture Collection
(Beijing, China). The cells were cultured in Dulbecco's modified
Eagle's media (DMEM; GE Healthcare Life Sciences, Logan, UT, USA)
supplemented with 10% fetal bovine serum (FBS; GE Healthcare Life
Sciences) and 1% penicillin/streptomycin (Gibco Life Technologies,
Carlsbad, CA, USA) at 37°C in a 5% CO2 incubator.
(S)-(+)-CPT, G418 were purchased from Sigma-Aldrich (St. Louis, MO,
USA). CPT and G418 were dissolved in dimethyl sulfoxide
(Sigma-Aldrich). Cancer cells were treated with CPT (0.12
µM) for 24 h and cells transfected with shRNA were screened
using G418 with concentrations of 300–800 µg/ml.
Vectors and primers
Of the three empty vectors used in the present
study, pEGFP-N1 (4.7 kb; cat. no. 6085-1) was purchased from
Addgene (Cambridge, MA, USA); pFLAG-CMV™-4 (6.3 kb; cat. no. E7158)
was purchased from Sigma-Aldrich; and pSilencer™ 2.1-U6 neo Vector
(4521 bp; cat. no. 113P06) was purchased from Invitrogen Life
Technologies (Carlsbad, CA, USA).
The sequences of the specific primers used for the
quantitative polymerase chain reaction (qPCR) were as follows:
HAX-1, sense 5′-ATGAGCCTCTTTGATCTCTTCC-3′ and antisense,
5′-CTACCGGGACCGGAACCAAC-3′ and β-actin, sense
5′-ACGTGGACATCCGCAAAGAC-3′ and antisense
5′-CTGCTGTCACCTTCACCGTTC-3′.
The HAX-1 primer sequences designed for plasmid
construction were as follows: HAX-1-up-HindIII,
5′-CCAAGCTTACCATGAGCCTCTTTGATCTCTTCC-3′; and
HAX-1-down-BamHI, 5′-CGGGATCCCTACCGGGACC GGAACCAAC-3′. The
length of fragments, amplified using qPCR with the HAX-1 and
β-actin primers, were 370 and 850 bp, respectively. The primers
were synthesized by Sangon Biotech Co., Ltd. (Shanghai, China).
Sequences for short hairpin (sh)HAX-1 and
small interfering (si)HAX-1
shHAX-1 and the longer lasting and more stable
siHAX-1 were designed and synthesized by Guangzhou RiboBio Co.,
Ltd. (Guangzhou, China), as follows: shHAX-1, sense 5′-GAT CCG CGG
ACA GAG ACT ACA GTA ATC AAG AGT TAC TGT AGT CTC TG TCC GTT TTT TGT
CGA CGG AAA-3′ and antisense 5′-GCG CCT GTC TCT GAT GTC ATT AGT TCT
CTC AAT GAC ATC AGA GAC AGG CAA AAA ACA GCT GCC TTT TCG A-3′; and
shRNA-con, sense 5′-GAT CCG GTC CAT GCA TGC CGT ATC AAG AGT ACG GCA
TGC ATG GAC TTT TTT GTC GAC GGA AA-3′ and antisense 5′-GCG TCC ATG
CAT GCC GTA AGT TCT CTC TAC GGC ATG CAT GGA CAA AAA ACA GCT GCC TTT
TCG A-3′; siHAX-1 dsRNA, sense 5′-CGG ACA GAG ACU ACA GUA A dTdT-3′
and antisense 5′-dTdT GCC UGU CUC UGA UGU CAU U-3′; scramble dsRNA,
sense 5′-TCC TGT GGC ATC CAC GAA ACT-3′ and antisense 5′-GAA GCA
TTT GCG GTG GAC GAT-3′. Restriction enzymes (BamHI and HindI; Santa
Cruz Biotechnology, Inc., Dallas, TX, USA) were used to cut at the
BamHI and HindIII sites, to insert the sequences into
the pSilencer™ 2.1-U6 neo Vector.
Transfection
Transfection was performed using
Lipofectamine® 2000 (Invitrogen Life Technologies) in
Opti-MEM (Gibco Life Technologies), according to the manufacturer's
instructions. The seed density of the cells was ~30% and the final
concentration of siRNA was 100 nM. The final concentrations of
shRNA and the overexpression vector were 1 µg/ml. The
temperature of incubation was 37°C The media was replaced after 5
h.
RNA extraction and reverse
transcription-qPCR (RT-qPCR)
Subsequent to homogenization on ice with the
TGrinder Electric Tissue Grinder [Tiangen Biotech (Beijing) Co.,
Ltd., Beijing, China], RNA was extracted from tumor tissues and the
adjacent nontumor tissues and purified using RNA extraction reagent
(TRIzol; Invitrogen Life Technologies), according to the
manufacturer's instructions. All the RNA samples used in the
present study were purified, and had optical density 260/280 ratios
of 1.8–2.0 (Nanodrop Spectrophotometer; Thermo Fisher Scientific,
Waltham, MA, USA). RT was performed using a Quantitect cDNA
synthesis system (Qiagen, Inc., Valencia, CA, USA) in a final
volume of 20 µl. Actin was used as the internal control. The
qPCR amplification reactions were conducted with QuantiFast SYBR
Green PCRMaster Mix (Qiagen, Inc.) on the ABI 7300 Real-time PCR
system (Applied Biosystems Life Technologies, Foster City, CA,
USA). The reaction conductions were as follows: Enzyme activation
at 95°C for 5 min, followed by 25 cycles at 94°C for 1 min, 58°C
for 1 min, extension at 72°C for 1 min, and a final extension step
at 72°C for 10 min. Following amplification, a melting curve was
created to confirm the specificity of the reaction. The relative
mRNA expression was then calculated by the 2−ΔΔCt
method.
Western blot analysis
For cell lysates, cells were rinsed with ice-cold
phosphate-buffered saline (PBS; Google Biotechnology Company,
China), harvested, centrifuged at 4°C, and cell pellets were lysed
by incubation at 4°C for 30 min in 500 µl lysis buffer
(Beyotime Institute of Biotechnology, Shanghai, China). For tissue
lysates, tissues were homogenized with the TGrinder Electric Tissue
Grinder with 500 µl lysis buffer. The concentration of
protein was performed using Pierce™ BCA Protein Assay kit (Thermo
Fisher Scientific) and the protein quantity for each lysate was
normalized to β-actin. The cell lysates were separated by SDS-PAGE
(12%; Wuhan Google Biotechnology, Ltd., Wuhan, China) at room
temperature, and transferred to polyvinylidene fluoride membranes
(Merck Millipore, Darmstadt, Germany) on ice. The blots were probed
at 4°C overnight with the following primary antibodies: Polyclonal
rabbit anti-human HAX-1 (1:200; Santa Cruz Biotechnology, Inc.;
sc-28268), polyclonal rabbit anti-human phosphorylated (p)-akt
(S473; sc-33437) and total-akt (1:200; Santa Cruz Biotechnology,
Inc.; sc-8312) and monoclonal mouse anti-β-actin (1:5,000;
Sigma-Aldrich; A5316). The primary antibody was detected by
chemiluminescence using horseradish peroxidase-conjugated
anti-mouse (OE185739) or anti-rabbit (NL181270) immunoglobulin G
secondary antibodies (Biofly Biotechnology Co., Wuhan, China).
Immunohistochemical staining
Following deparaffinization, then rehydratation in
xylene and graded (70, 98 and 100%) ethanol (Medical Company of
Hubei Province, Wuhan, China), the tissue samples were incubated
overnight with rabbit anti-HAX-1 (1:50, Santa Cruz Biotechnology,
Inc.) diluted in 1% FBS. A GTVision™ III kit (Gene Tech Company
Ltd., Shanghai, China) was used, according to the manufacturer's
instructions. The sections (4–10 µm thick) were then washed
in water, counterstained with hematoxylin (Google Biotechnology
Company, China), dehydrated in graded alcohols and xylene, and
mounted onto slides using Permount (Wuhan Google Biotechnology,
Ltd.). PBS was used as a blank control in place of the primary
antibody. The intensity of staining was classified as 0, negative;
1, weak; 2, moderate; and 3, strong. The expression levels of HAX-1
were analyzed by classifying the immunoreactivity scores. The
staining intensity was graded on a scale of 0–3 (0, no staining; 1,
weak immunoreactivity; 2, moderate immunoreactivity; 3, strong
immunoreactivity). The percentage of cells that exhibited positive
HAX-1 staining within the normal/cancerous region of a section was
scored as follows: 1, 0–25% of cells positive; 2, 26–50% positive;
3, 51–75% positive; 4, 76–100% positive for HAX-1. The staining
intensity score and the percentage immunoreactivity score were then
multiplied to obtain a composite score. The values of the composite
score ranged from 0–12, where a score of 0–3 was defined as low
expression, 4–6 was moderately high expression and >6 was high
expression.
Annexin V-fluorescein isothiocyanate
(FITC)/propidium iodide (PI) and annexin V-phycoerythrin
(PE)/7-aminoactinomycin D (7-AAD) detection
The SW480 cells (40–50%) were resuspended in 500
µl 1X binding buffer for 12 h at 37°C in 5% CO2;
subsequently 5 µl Annexin V and 5 µl PI/7-AAD (all
from BD Biosciences, Franklin Lakes, NJ, USA) were added to the
binding buffer. The cells were then gently suspended and incubated
for 15–20 min at room temperature in the dark, prior to being
analyzed using flow cytometry (FACSVantage; BD Biosciences)
(18).
Bromodeoxyuridine (BrdU) assay
A total of 10 µM BrdU (Sigma-Aldrich) was
added to the cells 1–2 h prior to harvesting. The cells were then
fixed in 70–80% cold ethanol and maintained at −20°C overnight. The
fixed cells were resuspended, following being washed in 1 ml 4 M
HCl (Google Biotechnology Company, China) at room temperature for
30–50 min in 0.1 M Brox solution (Sigma-Aldrich). Monoclonal
anti-BrdU antibody (1:200; BioLegend, San Diego, CA, USA) was then
added, and the cells were incubated in the dark at 4°C for ≥1 h.
The cells were then rinsed and incubated with a secondary
FITC-conjugated antibody (Dako, Glostrup, Denmark) for 30 min in
the dark at 4°C. After being incubated with 500 µl solution
containing 50 µg/ml PI and 50 µg/ml RNase at room
temperature for 20–30 min in the dark, the cells were analyzed by
flow cytometry.
Carboxyfluorescein succinimidyl ester
(CFSE) proliferation assay
The harvested cells were washed twice in cold PBS
and then suspended in 10 ml labeling reaction dilution
(Sigma-Aldrich). Following the addition of 200 µl 0.5 mM
CFSE (Beyotime Institute of Biotechnology, Haimen, China),
according to the manufacturer's instructions, the cells were
incubated at 37°C for 10 min. The reaction was terminated by adding
10 ml 10% FBS complete DMEM media, and the cells were washed twice.
Subsequently, the cells were cultured at 37°C for 0, 16, 24, 36 and
48 h, and analyzed using flow cytometry. The proliferation index
was calculated using ModFit LT for Mac V2.0 (BD Biosciences).
Statistical analysis
The t-test and c-test (or Fisher's exact test) were
conducted using SPSS software, version 19.0 (IBM SPSS, Armonk, NY,
USA) software. P<0.05 was considered to indicate a statistically
significant difference.
Results
Expression of HAX-1 and correlation with
the prognosis of CRC
The expression levels of HAX-1 in the individual
tumor and noncancerous tissue sections from the 60 patients with
CRC were detected using immunohistochemistry. As shown in Fig. 1, HAX-1 was ubiquitously expressed
in the colorectal tissues, as reported previously (19,20).
Notably, the most prominent expression of HAX-1 was detected in the
mucosal membrane of the colon, and was predominantly located in the
cytoplasm (Fig. 1A). The mRNA and
protein expression levels of HAX-1 were detected using RT-qPCR and
western blotting, respectively. The mRNA and protein expression
levels of HAX-1 were significantly higher in the CRC tissues, than
in the benign tissues (Fig. 1B and
C).
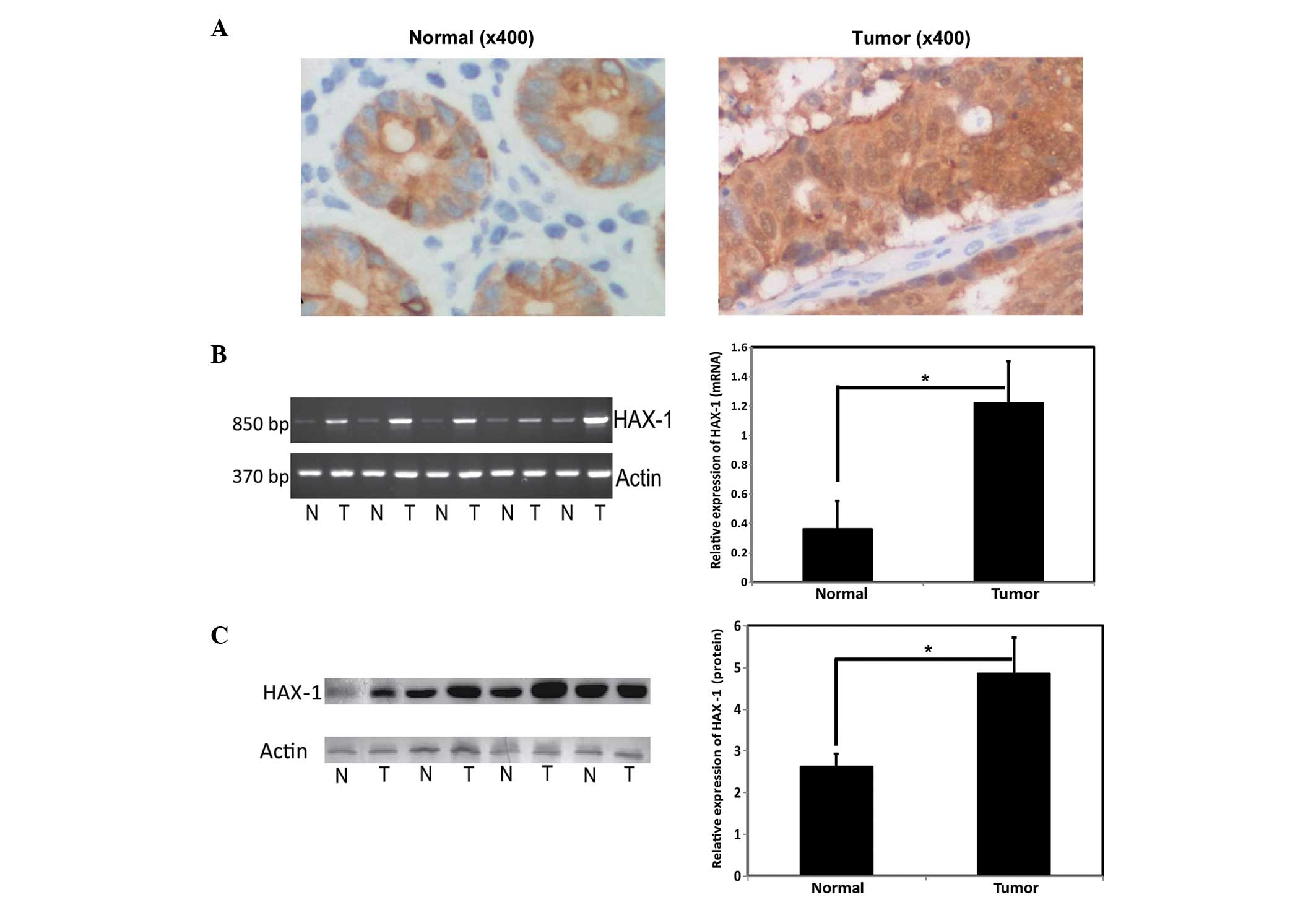 | Figure 1Expression of HAX-1 in CRC tissue. (A)
Representative immunohistochemical images of CRC tissue
(magnification, x400), Left, normal tissue; right, tumor tissue).
The expression levels of HAX-1 in CRC tissues were observed to be
significantly higher than those in benign/adjacent noncancerous
tissues. (B) mRNA expression levels of HAX-1 in colorectal cancer
tissues (Left, reverse transcription-quantitative polymerase chain
reaction; right, normalized with actin.) *P<0.05.
Values are presented as the mean ± standard deviation. (C) Protein
expression levels of HAX-1 in CRC tissues (Left, western blotting;
right, normalized with acctin). *P<0.05. Values are
presented as the mean ± standard deviation. HAX-1, hematopoietic
cell specific protein 1-associated protein X-1; N, normal tissue;
T, tumor tissue. CRC, colorectal cancer. |
Correlations between the expression levels of HAX-1
and the clinical data obtained from the patients are summarized in
Table I. The clinical factors
evaluated included age, gender, differentiation, lymph node
metastasis, muscular layer invasion, pathology type and Dukes'
stage. In addition, the patients were stratified into high and
moderate high groups according to the expression of HAX-1. Of the
60 primary CRC tissue samples, 50 exhibited high expression levels
of HAX-1. The high rate of expression was significantly increased
in the patients with CRC with lymph node metastasis, compared with
those without lymph node metastasis (91.67, vs. 75.00%,
respectively). Furthermore, patients with muscular layer invasion
exhibited higher expression levels of HAX-1, compared with those
without muscular layer invasion (90.2, vs. 44.44%, respectively). A
total of 22 of the 24 patients (91.67%) with Dukes C+D stages also
exhibited high expression levels of HAX-1. The expression levels of
HAX-1 were also increased in 24 of the patients with Dukes A+B;
however, the rate of increased expression was only 75.00%. No
significant correlations were observed between the expression of
HAX-1 and other prognostic factors, including, age, differentiation
and pathology type.
 | Table IAssociation between the gene
expression levels of HAX-1 and prognostic factors in 60 patients
with colorectal cancer. |
Table I
Association between the gene
expression levels of HAX-1 and prognostic factors in 60 patients
with colorectal cancer.
| Clinical factor | Number of patients
(n) | Expression of HAX-1
(n)
| High expression
rate (%) | P-value |
|---|
| High | Moderate high |
|---|
| Age (years) | | | | | >0.05 |
| <60 | 20 | 17 | 3 | 85.00 | |
| >60 | 40 | 33 | 7 | 82.50 | |
| Gender | | | | | >0.05 |
| Male | 41 | 35 | 6 | 85.37 | |
| Female | 19 | 15 | 4 | 78.95 | |
|
Differentiation | | | | | >0.05 |
| Low | 9 | 9 | 0 | 100 | |
| High/Moderate | 51 | 41 | 10 | 80.39 | |
| Lymph node
metastasis | | | | | 0.025 |
| No | 36 | 27 | 9 | 75.00 | |
| Yes | 24 | 22 | 2 | 91.67 | |
| Muscular layer
invasion | | | | | 0.001 |
| No | 9 | 4 | 5 | 44.44 | |
| Yes | 51 | 46 | 5 | 90.20 | |
| Pathology type | | | | | >0.05 |
|
Adenocarcinoma | 53 | 44 | 9 | 83.02 | |
| Mucinous
adenocarcinoma | 7 | 5 | 2 | 71.43 | |
| Dukes stage | | | | | 0.025 |
| A+B | 36 | 27 | 9 | 75.00 | |
| C+D | 24 | 22 | 2 | 91.67 | |
Effects of the expression of HAX-1 on
cell apoptosis
To identify the effects of HAX-1 on apoptosis,
HAX-1-overexpressing SW480 cells were constructed by transfecting
the cells with an adenovirus vector containing the EGFP expressing
reporter (Ad-HAX-1-EGFP). Western blot analysis was performed 48 h
post-transfection, which revealed that HAX-1 was overexpressed in
the SW480 cells transfected with the Ad-HAX-1-EGFP plasmid
(Fig. 2A, upper panel). A
significant increase in the resistance of the HAX-1-transfected
SW480 cells to camptothecin (CPT) was observed 8 h following
administration, whereas the SW480 cells without HAX-1 (Ad-EGFP) did
not exhibit resistance to CPT (Fig. 2B
and C). In addition, apoptosis was reduced in the SW480 cells
overexpressing-HAX-1.
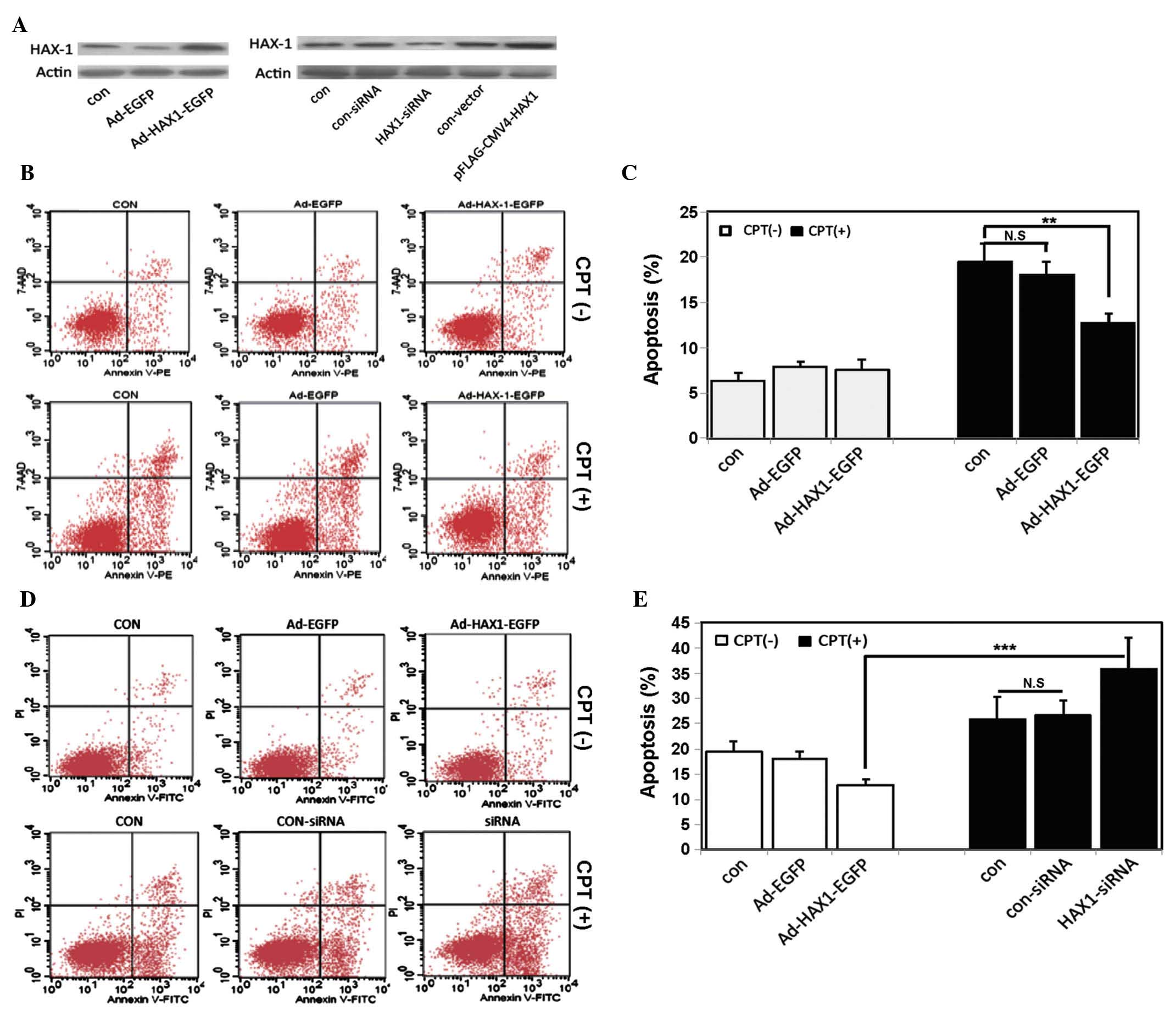 | Figure 2Effects of the expression of HAX-1 on
apoptosis. (A) SW480 human colorectal cancer cells were transfected
with an HAX-1 overexpression vector (upper panel) or an HAX-1-siRNA
vector (lower panel) for 48 h. The protein expression levels of
HAX-1 were detected using western blotting. (B) Representative
annexin V/7-AAD dot-plot distributions of SW480 cells treated
without (upper panel) or with (lower panel) CPT in the different
groups: Con, SW480 cells without infection; Ad-EGFP, SW480 cells
infected with empty vector; Ad-HAX-1-EGFP, SW480 cells infected
with Ad-HAX-1-EGFP vector. (C) Quantification of apoptosis.
Experiments were repeated at least three times. Values represent
the mean ± standard deviation (**P<0.01). (D)
Representative annexin V/PI dot-plot distributions of transfected
SW480 cells treated without (upper panel) or with (lower panel) CPT
in the different groups: Con, SW480 cells without transfection;
con-siRNA, SW480 cells transfected with control siRNA; siRNA, SW480
cells transfected with HAX-1-siRNA. (E) Quantification of
apoptosis. Experiments were repeated at least three times. Values
represent the mean ± standard deviation (***P<0.001).
HAX-1, hematopoietic cell-specific protein 1-associated protein
X-1; siRNA, small interfering RNA; CPT, camptothecin; siRNA, small
interfering RNA; N.S, not significant. |
To further confirm the effects of HAX-1 on
apoptosis, the expression of HAX-1 was silenced using an HAX-1
siRNA plasmid. As shown in Fig. 2,
the mRNA and protein expression levels of HAX-1 were significantly
decreased in the SW480 cells following transfection with HAX-1
siRNA (Fig. 2A, lower panel).
Following treatment with CPT for 8 h, apoptosis was increased in
the SW480 cells transfected with the HAX-1 siRNA plasmid (Fig. 2D and E). These results were
consistent with those of a previous study, which demonstrated that
HAX-1 had a functional role in inhibiting apoptosis in CRC
(7,21).
Effects of HAX-1 expression on cell
proliferation
CRC cells present with malignant proliferative
properties. To investigate the effects of HAX-1 on CRC cell
proliferation, a Brdu incorporation assay for DNA synthesis was
performed. The overexpression of HAX-1 promoted DNA synthesis, as
indicated by the increased number of Brdu-postive cells. However,
this promotion was repressed when the expression of HAX-1 was
silenced by siRNA, compared with the control SW480 cells (Figs. 3A and B). Similar results were
observed in the CFSE assay (Fig.
3C).
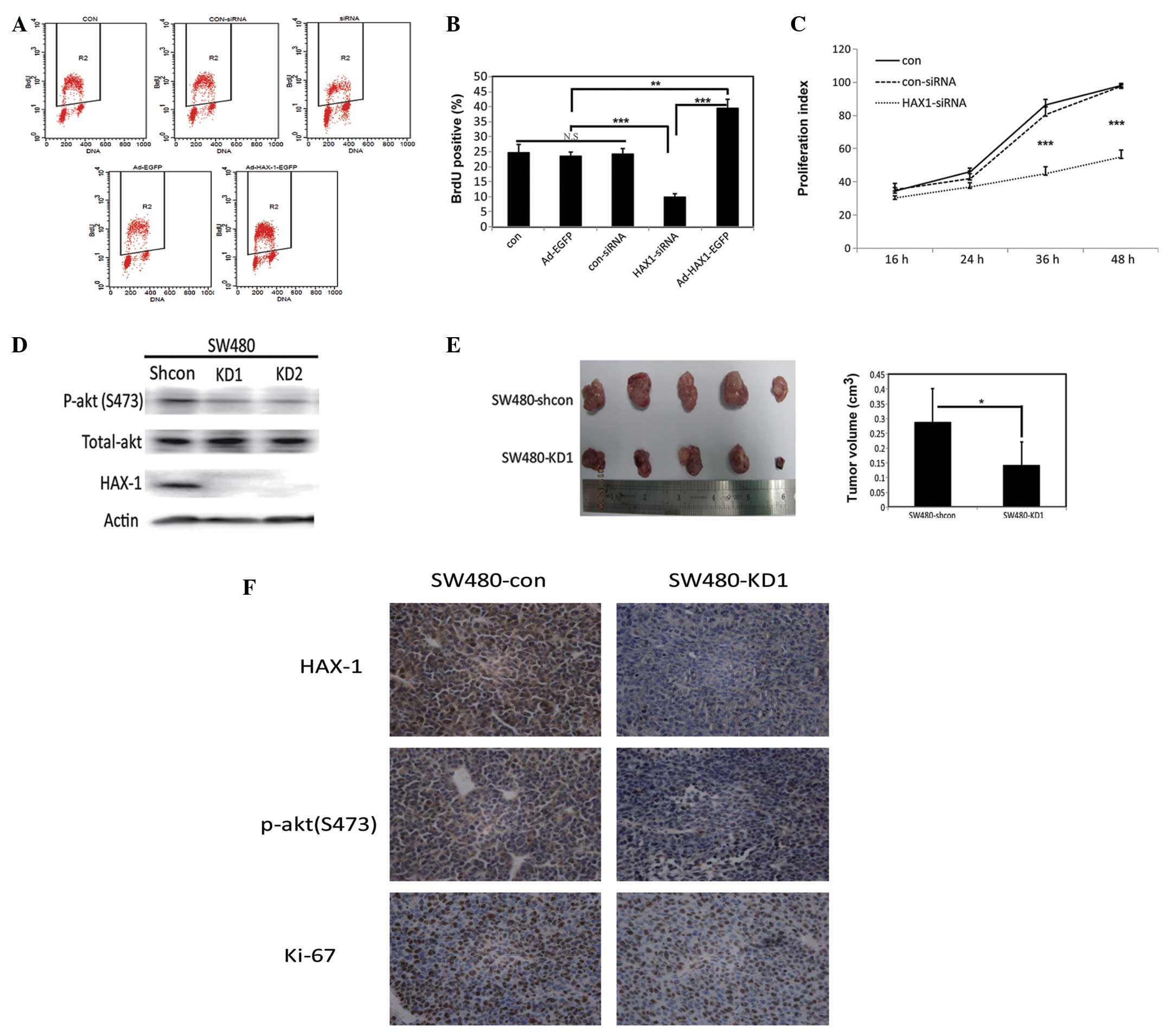 | Figure 3Effects of the expression of HAX-1 on
proliferation. (A) Representative dot-plot distributions of BrdU
assay using flow cytometry in the SW480 human colorectal cancer
cells transfected with Ad-EGFP, Ad-HAX-1-EGFP, con-siRNA or
HAX-1-siRNA for 48 h. (Con, SW480 cells without infection; Ad-EGFP,
SW480 cells infected with empty vector; Ad-HAX-1-EGFP, SW480 cells
infected with Ad-HAX-1-EGFP vector; con-siRNA, SW480 cells
transfected with control siRNA; siRNA, SW480 cells transfected with
HAX-1-siRNA). The R2, gate represents the percentage of BrdU
positive cells. (B) Quantification of Brdu positive cells.
Experiments were repeated at least three times. Values represent
the mean ± standard deviation (**P<0.01;
***P<0.001). (C) Proliferation index curve of SW480
cells transfected with con-siRNA and HAX-1-siRNA for 48 h. The
proliferation index was calculated based on fluorescence intensity,
detected using a CFSE assay. (Con, SW480 cells without
transfection; con-siRNA, SW480 cells transfected with scramble
siRNA; HAX-1-siRNA, SW480 cells transfected with HAX-1-siRNA). (D)
Western blot analysis of the expression levels of p-akt (S473),
total-akt and HAX-1 in SW480 cells transfected with shRNA-con
(Shcon), shRNA1 (KD1) or shRNA2 (KD2) vectors for 48 h. (E) Left
panel shows xenografts of BALB/c mice (n=5) transplanted with
5×105 HAX-1-knockdown SW480 cells (SW480-KD1), compared
with the control (SW480-shcon). The right panel shows
quantification of tumor volumes Values are presented as the mean ±
standard deviation (*P<0.05). (F) Representative
immunohistochemical images of HAX-1, p-akt (S473) and Ki-67 in the
mice xenografts. HAX-1, hematopoietic cell-specific protein
1-associated protein X-1; shRNA, short hairpin RNA; siRNA, small
interfering RNA; BrdU, bromodeoxyuridine; siRNA, small interfering
RNA; shRNA, short hairpin; N.S, not significant. |
The proliferation of SW480 cells was markedly
decreased when the cells were transiently transfected with
anti-HAX-1 siRNA for 48 h. To further understand the effects of
HAX-1 on cell proliferation, shRNA-HAX-1 was constructed. Stable
HAX-1-knockdown SW480 cell lines were generated through
transfection of the cells with potent shRNA targeting HAX-1
(SW480-KD), and the cells were subsequently screened using G418.
The knockdown efficiencies were then measured using western
blotting (Fig. 3D). Two stable
clones, SW480-KD1 and SW480-KD2, were obtained. A total of
5×105 SW480-KD1 cells were then subcutaneously implanted
into nude mice, and primary tumor growth was monitored. SW480-shcon
cells were injected into the control mice at the same time. After 2
weeks, the tumors were dissected and sizes of the tumor masses were
measured. As expected, the size of the tumor masses from the
HAX-1-knockdown mice were significantly smaller compared with those
fron the mice in the control group (Fig. 3E). Immunohistochemical analysis of
the xenografts from the mice injected with the SW480-KD1 cells
revealed a decrease in the expression of ki-67, indicating that
proliferation was markedly inhibited (Fig. 3F). As shown in Fig. 3D and F, the protein expression
levels of p-akt (S473) were decreased in the xenografts from the
mice injected with the SW480-KD1 cells. These results suggested
that HAX-1 may be involved in the AKT signaling pathway.
Discussion
HAX-1 is a multifunctional protein, which has been
recognized as a regulator of apoptosis and cell survival (7,8,13,22).
The expression of HAX-1 is increased in numerous malignancies,
including lung cancer, breast cancer, CRC, lymphoma, melanoma and
leukemia (23–26). In the present study, the expression
of HAX-1 in the CRC tissues was determined using
immunohistochemistry, RT-qPCR and western blotting. The results of
the present study demonstrated that the expression levels of HAX-1
were significantly upregulated in the tumor specimens, compared
with the adjacent noncancerous tissues, which was concordant with
the findings of previous studies (25,27).
Previous reports have demonstrated that patients with esophageal
squamous cell carcinoma overexpressing HAX-1 are more likely to
exhibit lymph node metastasis and have a poor prognosis (27). It has also been demonstrated that
upregulated HAX-1 is associated with advanced clinicopathological
characteristics and prognosis in CRC (25). The present study demonstrated that
the expression of HAX-1 was significantly associated with lymph
node metastasis, muscular layer invasion and clinical stage in CRC,
which suggested that HAX-1 may be important in the progression and
metastasis of CRC.
Apoptosis is involved in the regulation of cell
survival and homeostasis, and abnormal apoptotic processes can
promote carcinogenesis and chemotherapy resistance (28,29).
Previous studies have reported on HAX-1-mediated chemotherapy
resistance in T-cell leukemia and melanoma (26,30).
In the present study, ectopic overexpression of HAX-1 protected
SW480 cells from CPT, an agent that induces apoptosis via
activation of the intrinsic mitochondrial pathway (31). Conversely, the protection is
decreased when HAX-1 is suppressed by siRNA. These results support
the hypothesis that high expression levels of HAX-1 protect CRC
cells from genotoxic agents.
The results of the present study indicated that
HAX-1 was involved in the proliferation of CRC. Cellular
proliferation was monitored using BrdU and CFSE assays, which
demonstrated that upregulating the expression of HAX-1 promoted DNA
synthesis. However, this proliferation was inhibited by
HAX-1-siRNA. To confirm this finding, stable HAX-1-knockdown
(SW480-KD) and control (SW480-shcon) cells were implanted into nude
mice. The results revealed that downregulating the expression of
HAX-1 in the SW480 cells suppressed tumor growth in vivo in
the mice xenograft model. These results suggested that HAX-1 may be
a potent therapeutic molecular target for inhibiting the growth of
human cancer.
In conclusion, the present study demonstrated that
the expression levels of HAX-1 were significantly increased in
colorectal tissues, and may be important in the apoptosis and
proliferation of CRC cells. Further investigations are required to
elucidate the mechanism underlying the effects of HAX-1 on cell
apoptosis and proliferation.
Acknowledgments
The authors of the present study would like to thank
all of the patients, their families, investigators and students in
the investigations using clinical specimens. The present study was
supported by grants from the National Natural Science Foundation of
China (grant nos. 81272278, 81101943, 81171927, 81201638 and
81302309).
References
|
1
|
Siegel R, Desantis C and Jemal A:
Colorectal cancer statistics, 2014. CA Cancer J Clin. 64:104–117.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel R, Ma J, Zou Z and Jemal A: Cancer
statistics, 2014. CA Cancer J Clin. 64:9–29. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Eilstein D, Hedelin G and Schaffer P:
Incidence of colorectal cancer in Bas-Rhin, trend and prediction in
2009. Bull Cancer. 87:595–599. 2000.In French. PubMed/NCBI
|
|
4
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2012. CA Cancer J Clin. 62:10–29. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Adam R, Haller DG, Poston G, et al: Toward
optimized front-line therapeutic strategies in patients with
metastatic colorectal cancer-an expert review from the
International Congress on Anti-Cancer Treatment (ICACT) 2009.
Annals of Oncology. 21:1579–1584. 2010. View Article : Google Scholar
|
|
6
|
Hatakeyama S, Watanabe M, Fujii Y and
Nakayama KI: Targeted destruction of c-Myc by an engineered
ubiquitin ligase suppresses cell transformation and tumor
formation. Cancer Res. 65:7874–7879. 2005.PubMed/NCBI
|
|
7
|
Suzuki Y, Demoliere C, Kitamura D,
Takeshita H, Deuschle U and Watanabe T: HAX-1, a novel
intracellular protein, localized on mitochondria, directly
associates with HS1, a substrate of Src family tyrosine kinases. J
Immunol. 158:2736–2744. 1997.PubMed/NCBI
|
|
8
|
Chao JR, Parganas E, Boyd K, Hong CY,
Opferman JT and Ihle JN: Hax1-mediated processing of HtrA2 by Parl
allows survival of lymphocytes and neurons. Nature. 452:98–102.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wyllie AH, Kerr JF, Macaskill IA and
Currie AR: Adrenocortical cell deletion: the role of ACTH. J
Pathol. 111:85–94. 1973. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Radhika V, Onesime D, Ha JH and
Dhanasekaran N: Galpha13 stimulates cell migration through
cortactin-interacting protein Hax-1. J Biol Chem. 279:49406–49413.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yedavalli VS, Shih HM, Chiang YP, et al:
Human immunodeficiency virus type 1 Vpr interacts with
antiapoptotic mitochondrial protein HAX-1. J Virol. 79:13735–13746.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Vafiadaki E, Sanoudou D, Arvanitis DA,
Catino DH, Kranias EG and Kontrogianni-Konstantopoulos A:
Phospholamban interacts with HAX-1, a mitochondrial protein with
anti-apoptotic function. J Mol Biol. 367:65–79. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Vafiadaki E, Arvanitis DA, Pagakis SN, et
al: The anti-apoptotic protein HAX-1 interacts with SERCA2 and
regulates its protein levels to promote cell survival. Mol Biol
Cell. 20:306–318. 2009. View Article : Google Scholar :
|
|
14
|
Sun SJ, Feng L, Zhao GQ and Dong ZM: HAX-1
promotes the chemoresistance, invasion, and tumorigenicity of
esophageal squamous carcinoma cells. Dig Dis Sci. 57:1838–1846.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Banerjee A, Saito K, Meyer K, et al:
Hepatitis C virus core protein and cellular protein HAX-1 promote
5-fluorouracil-mediated hepatocyte growth inhibition. J Virol.
83:9663–9671. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Luo X, Li Z, Li X, et al: hSav1 interacts
with HAX1 and attenuates its anti-apoptotic effects in MCF-7 breast
cancer cells. Int J Mol Med. 28:349–355. 2011.PubMed/NCBI
|
|
17
|
Li X, Luo X, Li Z, et al: Screening of
binding proteins that interact with human Salvador 1 in a human
fetal liver cDNA library by the yeast two-hybrid system. Mol Biol
Rep. 39:8225–8230. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Jiang J, Liu L, Li X, Tao D, Hu J and Qin
J: Defining the restriction point in normal asynchronous human
peripheral blood lymphocytes. Cytometry A. 83:944–951.
2013.PubMed/NCBI
|
|
19
|
Lees DM, Hart IR and Marshall JF:
Existence of multiple isoforms of HS1-associated protein X-1 in
murine and human tissues. J Mol Biol. 379:645–655. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Carlsson G, van't Hooft I, Melin M, et al:
Central nervous system involvement in severe congenital
neutropenia: neurological and neuropsychological abnormalities
associated with specific HAX1 mutations. J Intern Med. 264:388–400.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Han Y, Chen YS, Liu Z, et al:
Overexpression of HAX-1 protects cardiac myocytes from apoptosis
through caspase-9 inhibition. Circ Res. 99:415–423. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Fadeel B and Grzybowska E: HAX-1: a
multifunctional protein with emerging roles in human disease.
Biochim Biophys Acta. 1790:1139–1148. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Rhodes DR, Yu J, Shanker K, et al:
ONCOMINE: a cancer microarray database and integrated data-mining
platform. Neoplasia. 6:1–6. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Trebinska A, Rembiszewska A, Ciosek K, et
al: HAX-1 overexpression, splicing and cellular localization in
tumors. BMC Cancer. 10:762010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wei XJ, Li SY, Yu B, Chen G, Du JF and Cai
HY: Expression of HAX-1 in human colorectal cancer and its clinical
significance. Tumour Biol. 35:1411–1415. 2014. View Article : Google Scholar
|
|
26
|
Ashhab Y, Alian A, Polliack A, Panet A and
Ben Yehuda D: Two splicing variants of a new inhibitor of apoptosis
gene with different biological properties and tissue distribution
pattern. FEBS Lett. 495:56–60. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Li M, Tang Y, Zang W, et al: Analysis of
HAX-1 gene expression in esophageal squamous cell carcinoma. Diagn
Pathol. 8:472013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kaufmann SH and Gores GJ: Apoptosis in
cancer: cause and cure. Bioessays. 22:1007–1017. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Schmitt CA: Senescence, apoptosis and
therapy - cutting the lifelines of cancer. Nat Rev Cancer.
3:286–295. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
30
|
Nachmias B, Ashhab Y, Bucholtz V, et al:
Caspase-mediated cleavage converts Livin from an antiapoptotic to a
proapoptotic factor: implications for drug-resistant melanoma.
Cancer Res. 63:6340–6349. 2003.PubMed/NCBI
|
|
31
|
Deng W, Wang DA, Gosmanova E, Johnson LR
and Tigyi G: LPA protects intestinal epithelial cells from
apoptosis by inhibiting the mitochondrial pathway. Am J Physiol
Gastrointest Liver Physiol. 284:G821–G829. 2003. View Article : Google Scholar : PubMed/NCBI
|














