Introduction
Colorectal cancer (CRC) is one of the most common
malignant tumor types, and the incidence of CRC is increasing by 2%
per year worldwide. Although the prognosis for CRC has greatly
improved with the continuous development of diagnostic techniques,
the mortality rate remains high (1). The major causes of CRC-associated
mortality are recurrence and metastasis. Loss of intercellular
adhesions has a vital role during the process of tumor invasion and
metastasis. E-cadherin is the major molecule responsible for
maintaining intercellular adherens junctions, and loss of
E-cadherin expression is closely associated with de-differentiation
and metastasis in a variety of tumor types (2,3).
E-cadherin can maintain the stability of intercellular adherens
junctions by binding with multiple catenins to form the cadherin
catenin complex.
δ-Catenin is one member of the catenin family and
consists of 10 Armadillo (Arm) repeats. Initially, δ-catenin was
considered to only be expressed in cerebral neurons, where it has
been shown to bind with presenilin to exert a function in the
development of Alzheimer's disease (4–6).
However, previous studies have reported a close correlation between
the expression of δ-catenin and cancer. In a comparative microarray
study of benign prostatic hyperplasia and prostate cancer, Burger
et al (7) verified that
δ-catenin was markedly upregulated at the transcriptional level in
prostate adenocarcinoma. Subsequently, Lu et al (8) demonstrated that δ-catenin was
significantly overexpressed and associated with the Gleason score
in prostate cancer. Furthermore, Zhang et al (9) reported that δ-catenin is
overexpressed in lung cancer tissues and can promote a malignant
phenotype in non-small cell lung cancer cells via enhancing the
activity of the transcription factor Kaiso (10). These studies indicated that
δ-catenin has a role in the initiation and progression of cancer.
However, it has remained elusive whether δ-catenin is overexpressed
in CRC, or whether the expression of δ-catenin is correlated with
the clinicopathological features of CRC.
In the present study, δ-catenin protein expression
was determined in 110 cases of CRC using immunohistochemistry, and
the correlation between δ-catenin expression and the
clinico-pathological features of CRC was investigated. In addition,
the expression of δ-catenin in primary tumor foci and lymph node
metastases was compared in 40 matched tissues from CRC patients
with lymph node metastases. The prognostic value of δ-catenin in
the 70 cases of CRC for which complete follow-up data were
available was determined. Finally, δ-catenin protein and mRNA
expression were compared in 30 paired CRC and adjacent normal
tissues, and the correlation between the expression of δ-catenin
mRNA and the clinicopathological features of CRC was
investigated.
Materials and methods
Tissue samples
Formalin-fixed, paraffin-embedded (FFPE) blocks from
110 cases of CRC and 30 normal colorectal tissue specimens were
obtained from the Department of Pathology, Shengjing Hospital of
China Medical University (Shenyang, China). None of the patients
had received radiotherapy, chemotherapy or immunotherapy prior to
tumor excision. In total, 67 of the patients were male and 43 were
female (1.56:1 male-to-female ratio). The patients' age at the time
of surgery ranged from 31 to 87, with an average age of 61
years.
Complete follow-up data were available for 70 of the
cases of primary CRC, which were surgically excised between May
2004 and July 2005. In addition, lymph node metastases were present
in 40 of the 110 cases. To evaluate the tumor, nodes and metastasis
(TNM) stage, at least 12 lymph nodes were obtained during surgical
resection. All tumors were classified according to the TNM staging
system, as revised by the International Union Against Cancer (UICC)
in 2002 (11). All specimens were
re-evaluated for diagnosis according to the World Health
Organization (WHO) criteria (12)
for the classification of colorectal cancer. All 110 of the CRC
specimens were adenocarcinomas, with 14 stage-I cases, 39 stage-II
cases, 42 stage-III cases and 15 stage-IV cases; furthermore, 36
cases were highly differentiated, 55 cases were moderately
differentiated and 19 cases were poorly differentiated.
In addition, paired tumor and non-tumor tissues
(>5 cm from the primary tumor edge) were obtained from 30 cases
of CRC, immediately frozen in liquid nitrogen and stored at −70°C
for RNA and protein analysis. The present study was conducted in
accordance with the regulations of and was approved by the
Institutional Review Board of China Medical University. Informed
consent was obtained prior to surgery from all enrolled
patients.
Immunohistochemical staining and
evaluation
FFPE tissue blocks were cut into 4-µm
sections and mounted on poly-L-lysine-coated slides (SLI-2002;
MaiXin, Fuzhou, China). The sections were de-paraffinized in
xylene, re-hydrated using a graded ethanol series, incubated with
3% hydrogen peroxide in methanol for 20 min, and then washed in
phosphate buffered saline (PBS) for 5 min (All from MaiXin).
Antigen retrieval was conducted using enzymatic antigen retrieval
solution for 15 min at room temperature, and then the sections were
washed in PBS for 5 min and blocked using normal blocking serum
from the Ultrasensitive™ S-P kit (KIT-9720; MaiXin) for 30 min. The
sections were incubated with anti-δ-catenin primary antibody
(1:400; Abcam, Cambridge, UK) at 4°C overnight. Following washing
with PBS, the sections were stained using diaminobenzidine
tetrahydrochloride (MaiXin) as a chromogen, lightly counter-stained
with hematoxylin (MaiXin), dehydrated and mounted. For the negative
controls, the primary antibody was replaced with PBS.
The scoring criteria for δ-catenin were identical to
those described by Lu et al (8). All sections were assessed by three
observers blinded to the study. Cases with discrepancies were
jointly re-evaluated by the investigators, and a consensus was
obtained. The sections were evaluated at low magnification (×100)
with the Olympus IX51 microscope (Olympus America, Inc., Melville,
NY, USA) to identify areas in which δ-catenin was evenly stained. A
total of 400 tumor cells were counted and the percentage of
positively-stained cells was calculated. The proportion of cells
exhibiting δ-catenin expression was categorized as follows: 0,
<1%; 1, 1–25%; 3, 51–75%; and 4, >75%. The relative staining
intensity was categorized as follows: 0 (no staining); 1 (weak); 2
(intermediate) and 3 (strong). The proportion and intensity scores
were multiplied to obtain a total score; scores <2 were
considered negative, while scores ≥2 were considered positive.
Reverse transcription quantitative
polymerase chain reaction (RT-qPCR)
RT-qPCR was performed following the MIQE guidelines
(Minimum Information for Publication of Quantitative Real-Time PCR
Experiments) published by Bustin et al (13). Total RNA was isolated from the
tissues using TRIzol (Invitrogen Life Technologies, Carlsbad, CA,
USA) according to the manufacturer's instructions. 1% agarose gel
electrophoresis was performed for 30 min to detect the degradation
of RNA samples. The concentration and purity of extracted RNA were
determined using the GenQuant RNA/DNA calculator (GeneQuant
1300/100; Amersham-Pharmacia Biotech, Cambridge, UK).
Deoxyribonuclease (DNase) treatment was performed using DNase I
(Invitrogen Life Technologies). 1 µg RNA was reverse
transcribed using the high-capacity cDNA RT kit (Applied
Biosystems, Thermo Fisher Scientific, Waltham, MA, USA) following
the manufacturer's instructions.
Real-time PCR was performed using the ABI Prism
7900HT Fast System (Applied Biosystems) with SYBR Premix Ex Taq II
(Takara, Dalian, China). Amplifications were performed in a total
volume of 10 µl using the primer sequences from Takara
listed in Table I, with an initial
dena-turation at 95°C for 30 sec followed by 40 cycles of 95°C for
5 sec and 60°C for 30 sec. GAPDH was used as an internal control.
The presence of single PCR amplicons in each reaction was confirmed
using melting curve analysis. The data were analyzed using the
2−ΔCq method with the SDS 2.4 software package (Applied
Biosystems) following the method of Schmittgen and Livak (14).
 | Table IPrimers and amplification range used
in reverse transcription quantitative polymerase chain
reaction. |
Table I
Primers and amplification range used
in reverse transcription quantitative polymerase chain
reaction.
| Gene name | Primer sequence
sequence (5′→3′) | Amplification range
(bp) |
|---|
| δ-catenin | Forward:
TTCATCACAGGTGCTGCGTAA | 2266–2358 |
| Reverse:
CCATCACACTCTCTCATCCTTCTG | (NM_001332.2) |
| GAPDH | Forward:
GGTGAAGGTCGGAGTCAACG | 111–232 |
| Reverse:
CCATGTAGTTGAGGTCAATGAAG | (NM_002046.3) |
Western blot analysis
Total protein was extracted from the colorectal
tissues using lysis buffer (150 mM NaCl, 1% NP-40, 0.1% SDS, 2
mg/ml aprotinin and 1 mM phenylmethanesulfonylfluoride; Beyotime
Insititute of Biotechnology, Shanghai, China) for 30 min at 4°C,
the lysates were centrifuged at 14,000 ×g for 30 min at 4°C and the
supernatants were collected. Aliquots containing 50 mg protein were
separated by 8% SDS-PAGE (Beyotime Institute of Biotechnology) and
transferred to PVDF membranes (Merck Millipore, Darmstadt, Germany)
at 100 V for 2.5 h at 4°C. The protein concentration was determined
using the bicinchoninic acid assay (Beyotime Institute of
Biotechnology). The membranes were blocked in 5% skimmed milk for 2
h, and the proteins were detected by incubation with mouse
monoclonal antibodies against δ-catenin (ab54578; 1:500; Abcam,
Cambridge, UK) or GAPDH (E021010-01; 1:1,000; EarthOx, San
Francisico, CA, USA) overnight at 4°C, followed by anti-mouse
immunoglobulin G conjugated to horse radish peroxidase at a
dilution of 1:2,000 for 2 h at room temperature. The bands were
visualized using the EC3 Imaging System (UVP LLC, Upland, CA, USA)
and the optical densities were normalized to GAPDH using ImageJ
software, version 1.44 (NIH, Bethesda, MD, USA).
Statistical analysis
All in vitro experiments were performed at
least three times and values are expressed as the mean ± standard
deviation. Pearson's χ2 test was used to analyze the
correlation between δ-catenin expression and clinicopatho-logical
features of CRC. The overall survival probabilities were calculated
using the Kaplan-Meier method and compared using the log-rank test.
To determine the significant factors associated with overall
survival, a multivariate Cox proportional hazard model was created.
All statistical analyses were performed using SPSS 13.0 for Windows
(SPSS Inc., Chicago, IL, USA). P<0.05 was considered to indicate
a statistically significant difference between values.
Results
δ-Catenin is overexpressed and associated
with poor prognosis in CRC
Negative or weak δ-catenin immunoreactivity was
observed in the cytoplasm of normal colorectal mucosal epithelial
cells (Fig. 1A). Based on the
scoring criteria, all of the normal colorectal mucosa sections were
scored as negative. Varying degrees of δ-catenin-positive staining
were observed in CRC, mainly in the cytoplasm of the tumor cells
(Fig. 1B and C). The rate of
positive δ-catenin expression in CRC (75/110; 68.18%) was
significantly higher than that in normal colorectal tissues (11/30,
36.7%; P<0.001).
There was no significant association between
positive δ-catenin expression and the age, gender, lesion location
or tumor size in CRC patients (P>0.05); however, positive
δ-catenin expression was closely associated with the degree of
differentiation, TNM stage and lymph node metastasis in CRC
(Table II). The rates of positive
δ-catenin expression in poorly differentiated CRC (18/19; 94.74%),
moderately differentiated CRC (36/55; 65.45%) and
well-differentiated CRC (21/36; 58.33%) were significantly
different (P=0.012 according to Fisher's probabilities test). The
rate of positive δ-catenin expression in stage-I–II CRC (56.6%;
30/53) was significantly lower than that in stage-III–IV CRC
(78.95%; 45/57; P<0.05). The rate of positive δ-catenin
expression in the tumors of patients with lymph node metastasis
(79.25%; 42/53) was significantly higher than that in patients
without lymph node metastasis (57.89%; 33/57; P= 0.016). In the 40
cases of CRC with matched lymph node metastases specimens, the
expression rate of δ-catenin in lymph node metastases (92.5%;
37/40; Fig. 1D) was significantly
higher than that in the corresponding primary tumor foci (75%;
30/40; P=0.002; Fig. 1C).
| Table IIClinical and histological features of
110 patients with colorectal cancer. |
Table II
Clinical and histological features of
110 patients with colorectal cancer.
| Variable | All patients (n) | δ-catenin
| P-value |
|---|
| Positive (n) | Negative (n) |
|---|
| Total | 110 | 75 | 35 | |
| Age (years) | | | | 0.738 |
| <61 | 54 | 36 | 18 | |
| ≥61 | 56 | 39 | 17 | |
| Gender | | | | 0.775 |
| Male | 67 | 45 | 22 | |
| Female | 43 | 30 | 13 | |
| Lesion location | | | | 0.686 |
| Rectum | 69 | 48 | 21 | |
| Colon | 41 | 27 | 14 | |
| Size (cm) | | | | 0.122 |
| <5 | 67 | 42 | 25 | |
| ≥5 | 43 | 33 | 10 | |
| TNM stage | | | | 0.012 |
| I–II | 53 | 30 | 23 | |
| III–IV | 57 | 45 | 12 | |
| Grade | | | | 0.012a |
| Well | 36 | 21 | 15 | |
| Moderate | 55 | 36 | 19 | |
| Poor | 19 | 18 | 1 | |
| Lymph node
metastasis | | | | 0.016 |
| Yes | 53 | 42 | 11 | |
| No | 57 | 33 | 24 | |
For the 70 CRC patients with complete follow-up
data, the five-year survival rate was 64.29% (45/70). Kaplan-Meier
survival analysis demonstrated that the mean survival time of
patients with positive δ-catenin expression was significantly
shorter than that of patients with negative δ-catenin expression
(84.02±4.63 vs. 58.53±5.22 months; Log-Rank test P=0.005; Fig. 2), indicating that positive
expression of δ-catenin may be indicative of a poor prognosis in
CRC patients. The Cox model multivariate analysis confirmed that
the TNM stage (P= 0.009) and positive expression of δ-catenin
(P=0.031) were independent risk factors, which affected the
prognosis of patients with CRC (Table III).
| Table IIICox regression model for the
prediction of survival of 70 patients with colorectal cancer. |
Table III
Cox regression model for the
prediction of survival of 70 patients with colorectal cancer.
| Factor | β | SE | P-value | Exp (β) | 95% CI for Exp
(β) |
|---|
| Age | 0.012 | 0.018 | 0.528 | 1.012 | 0.976–1.049 |
| Gender | −0.809 | 0.461 | 0.080 | 0.445 | 0.180–1.100 |
| Lesion
location | −0.511 | 0.441 | 0.247 | 0.600 | 0.253–1.425 |
| Tumor size | −0.408 | 0.473 | 0.388 | 0.665 | 0.263–1.680 |
| Grade | 0.149 | 0.307 | 0.626 | 1.161 | 0.636–2.119 |
| TNM stage | 1.598 | 0.608 | 0.009 | 4.945 | 1.502–16.281 |
| Lymph node
metastasis | −0.948 | 0.669 | 0.156 | 0.387 | 0.104–1.437 |
| δ-catenin
expression | 1.113 | 0.515 | 0.031 | 3.042 | 1.109–8.343 |
Expression of δ-catenin mRNA and protein
are significantly upregulated in CRC
The expression of δ-catenin mRNA was assessed by
RT-qPCR in 30 paired colorectal cancer specimens and the adjacent
normal colorectal tissues. The relative δ-catenin mRNA expression
levels were seven-fold higher in CRC tissues (22.61±2.57) than
those in the matched normal colorectal tissues (3.25±0.88; t=7.137,
P<0.0001; Fig. 3).
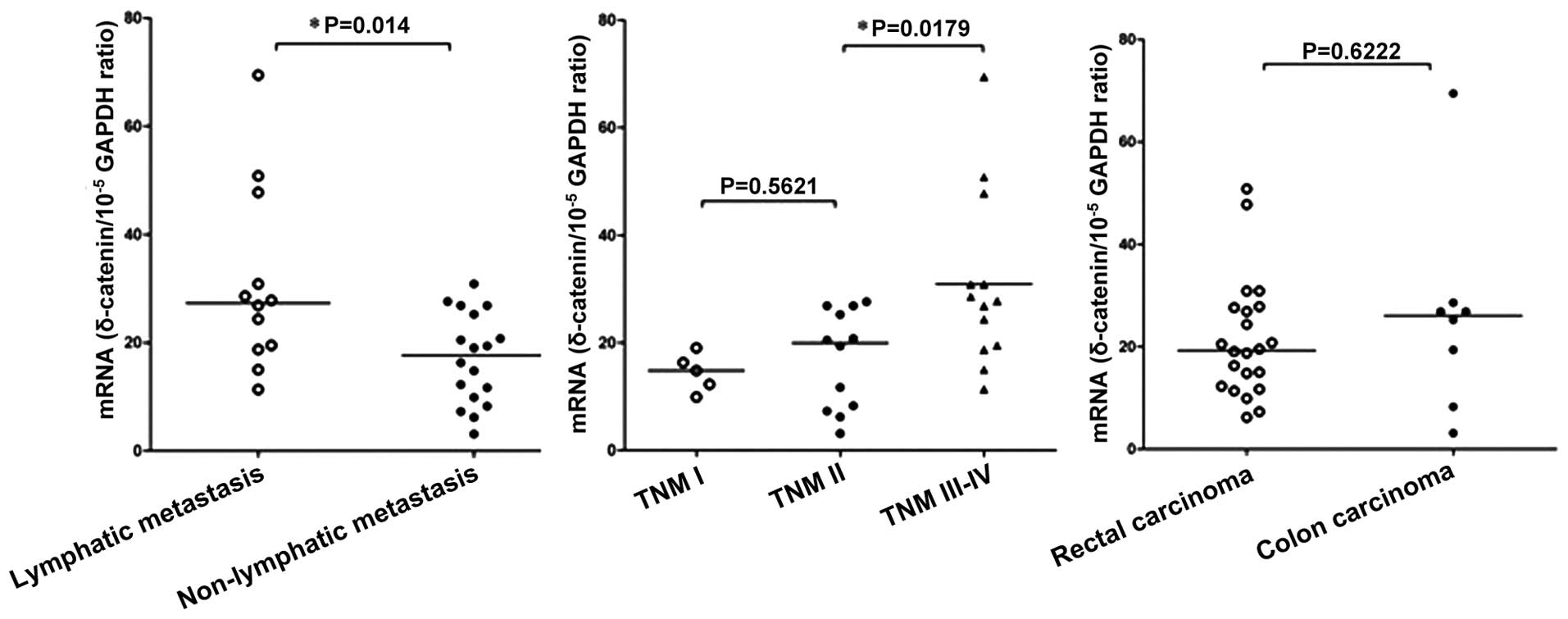
The correlation between δ-catenin mRNA expression
and the clinicopathological features of CRC was examined in 30
patients (Fig. 4). A significant
correlation was observed between the expression of δ-catenin mRNA
and lymph node metastasis, as the δ-catenin mRNA expression levels
were 1.8-fold higher in patients with lymph node metastasis
(30.93±4.89) than those in patients without metastasis
(17.06±1.971; P=0.014). Furthermore, a positive correlation was
observed between TNM stage and δ-catenin mRNA expression. The
Kruskal-Wallis test revealed that the δ-catenin mRNA expression
levels were significantly different in CRC tissues at different TNM
stages (P=0.0116), with the most significant difference observed
between stage III–IV and stage I–II (P=0.0179). There was no
correlation between the δ-catenin mRNA expression levels and tumor
location, consistent with the immunohistochemistry results.
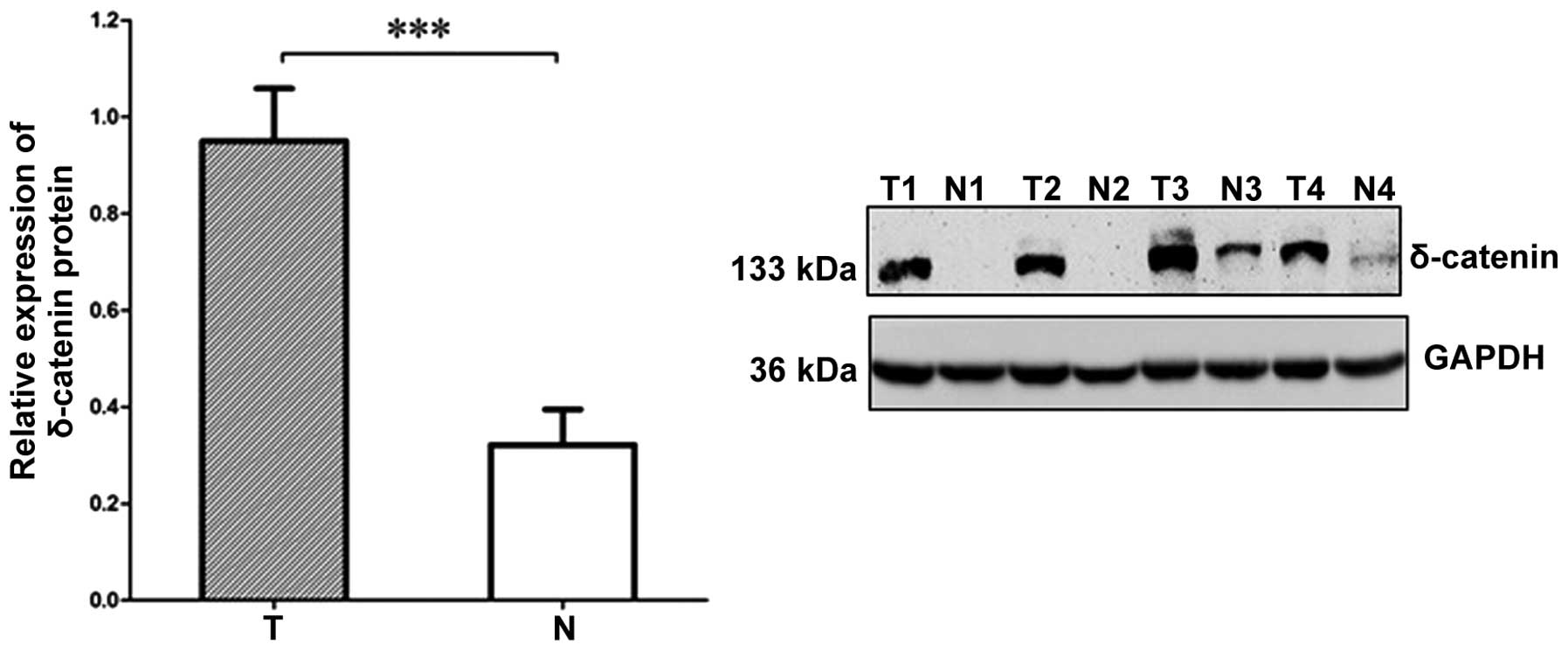
Finally, the δ-catenin protein expression levels
were determined in 30 cases of CRC and the paired adjacent normal
colorectal tissues using western blot analysis (Fig. 5). The average δ-catenin protein
expression in the normal colorectal tissues (0.32±0.07) was
significantly lower than that in the matched CRC tissues
(0.95±0.11; P<0.05).
Discussion
Initially, δ-catenin mRNA was considered to be
mainly expressed in the brain. δ-Catenin can bind presenilin
(5,6) and is considered to be a major
adherens junction-associated protein (4,15–18).
Subsequent studies revealed that δ-catenin mRNA is also weakly
expressed in pancreatic tissues (5). Paffenholz et al (19) provided the first evidence of
mammalian δ-catenin protein expression in the external limiting
membrane of the retina. δ-Catenin mRNA was also shown to be
expressed at low levels in a number of tumor cell lines, including
PC12 and human neuroblastoma cells; however, these cell types have
the capacity for neuronal differentiation (20). δ-catenin has been shown to be
upregulated in >80% (55/65) of prostate adenocarcinoma samples,
and expression of δ-catenin was demonstrated to be positively
correlated with the Gleason score in prostate adenocarcinoma
(8). In addition, δ-catenin is
expressed at high levels in >60% of patients with non-small cell
lung cancer (9), and δ-catenin has
been shown to promote a malignant phenotype in non-small cell lung
cancer cell by affecting the activity of the transcriptional
repressor Kaiso (10).
Analysis of human expressed sequence tags
demonstrated that δ-catenin mRNA may be expressed in multiple tumor
types, including esophageal, ovarian and breast cancer (21); however, the expression and clinical
significance of δ-catenin in numerous tumor types has remained
elusive. To determine the role of δ-catenin in CRC, the present
study determined the expression of δ-catenin in 110 cases of CRC
and 15 adjacent normal colorectal tissues using
immunohistochemistry. δ-Catenin was observed to be absent or weakly
expressed in the cytoplasm of normal colorectal epithelial cells,
which were classified as δ-catenin-negative according to the
scoring criteria. Overexpression of δ-catenin was observed in ~70%
of the CRC tissues, similar to the results of studies on non-small
cell lung cancer (9,10). In addition, semi-quantitative
western blot analysis confirmed that δ-catenin protein was
overex-pressed in CRC tissues.
The immunohistochemical analysis in the present
study demonstrated that δ-catenin was mainly expressed in the
cytoplasm of CRC cells, with only small amounts of δ-catenin
observed at the cell junctions, in agreement with the localization
of δ-catenin in prostate cancer and lung cancer as reported by Lu
et al (8) and Zhang et
al (9). It is known that
δ-catenin can bind to the juxtamembrane domain of E-cadherin to
exert a function in the formation and stability of adherens
junctions (15); however, the
purpose of the abundance of δ-catenin in the cytoplasm of tumor
cells is currently elusive. It is possible that overexpression of
δ-catenin may lead to supersaturation of E-cadherin and abnormal
accumulation of δ-catenin in the cytoplasm. Alternatively,
E-cadherin may lose its function as an adhesion molecule in CRC,
and as a consequence, be rarely expressed at the cell membrane.
Further study regarding the metabolism of δ-catenin and its
association with E-cadherin is required to explain the cytoplasmic
expression pattern of δ-catenin in CRC.
Zhang et al (9) and Dai et al (10) demonstrated that δ-catenin is not an
independent prognostic factor in non-small cell lung cancer;
however, an association between δ-catenin and poor prognosis was
observed. In the present study, over-expression of δ-catenin in CRC
was associated with poor differentiation, high TNM stage and lymph
node metastasis. The rate of positive δ-catenin expression in lymph
node metastases (92.5%; 37/40) was significantly higher than that
in the matched primary CRC tumor foci (75%; 30/40). Survival
analysis revealed that the mean survival time of CRC patients with
positive δ-catenin expression was markedly shorter than that of
patients with negative δ-catenin expression, and multivariate
analysis confirmed that δ-catenin was an independent risk factor
which affected the survival of CRC patients (P=0.031). These
results demonstrated that positive expression of δ-catenin is
associated with a poorer prognosis in CRC, and indicate that
δ-catenin may have an important role in the occurrence and
development of colorectal cancer. δ-Catenin should be considered as
a potential prognostic factor for predicting the clinical outcome
of patients with colorectal cancer.
It has been reported that increased expression of
δ-catenin in tumor tissues may be linked to overexpression of the
transcription factors E2F1 and Pax6 (22). However, it has also been suggested
that the transcription of δ-catenin is not altered in tumor
tissues, and that the increased protein expression levels are due
to increased translational efficiency (23). In the present study, RT-qPCR
analysis demonstrated that the expression of δ-catenin mRNA was
significantly increased in CRC tissues compared to that in the
matched normal tissues. In addition, the δ-catenin mRNA expression
levels were positively correlated with the pathological stage and
lymph node metastasis. These results indicated that overexpression
of δ-catenin protein in CRC is due to increased transcription of
δ-catenin mRNA; however, further study is required to identify the
specific mechanisms responsible for this process.
In conclusion, the presents study showed that
δ-catenin protein is overexpressed and mainly localizes to the
cytoplasm in CRC. Positive expression of δ-catenin was associated
with poor differentiation, higher TNM stage, lymph node metastasis
and poor prognosis in CRC. Expression of δ-catenin mRNA was
upregulated in CRC compared to the corresponding adjacent normal
tissues, and the δ-catenin mRNA expression levels were positively
correlated with the tumor stage and lymph node metastasis in CRC.
Hence, δ-catenin may represent a potentially clinically useful
independent prognostic factor in CRC.
Acknowledgments
This work was supported by grants from the Liaoning
Province Natural Science Foundation of China (no. 2013021098 to
Hong Zhang) and the National Natural Science Foundation of China
(no. 81401881 to Shun-Dong Dai and no. 81372338 to Shu-Li Liu).
References
|
1
|
Boedefeld WM II, Bland KI and Heslin MJ:
Recent insights into angiogenesis, apoptosis, invasion and
metastasis in colorectal carcinoma. Ann Surg Oncol. 10:839–851.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Gumbiner BM: Regulation of
cadherin-mediated adhesion in morphogenesis. Nat Rev Mol Cell Biol.
6:622–634. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Makrilia N, Kollias A, Manolopoulos L and
Syrigos K: Cell adhesion molecules: Role and clinical significance
in cancer. Cancer Invest. 27:1023–1037. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Paffenholz R and Franke WW: Identification
and localization of a neurally expressed member of the
plakoglobin/armadillo multigene family. Differentiation.
61:293–304. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhou J, Liyanage U, Medina M, Ho C,
Simmons AD, Lovett M and Kosik KS: Presenilin 1 interaction in the
brain with a novel member of the Armadillo family. Neuroreport.
8:2085–2090. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Tanahashi H and Tabira T: Isolation of
human delta-catenin and its binding specificity with presenilin 1.
Neuroreport. 10:563–568. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Burger MJ, Tebay MA, Keith PA, Samaratunga
HM, Clements J, Lavin MF and Gardiner RA: Expression analysis of
delta-catenin and prostate-specific membrane antigen: Their
potential as diagnostic markers for prostate cancer. Int J Cancer.
100:228–237. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lu Q, Dobbs LJ, Gregory CW, Lanford GW,
Revelo MP, Shappell S and Chen YH: Increased expression of
delta-catenin/neural plakophilin-related armadillo protein is
associated with the down-regulation and redistribution of
E-cadherin and p120ctn in human prostate cancer. Hum Pathol.
36:1037–1048. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang JY, Wang Y, Zhang D, Yang ZQ, Dong
XJ, Jiang GY, Zhang PX, Dai SD, Dong QZ, Han Y, et al:
delta-Catenin promotes malignant phenotype of non-small cell lung
cancer by non-competitive binding to E-cadherin with p120ctn in
cytoplasm. J Pathol. 222:76–88. 2010.PubMed/NCBI
|
|
10
|
Dai SD, Wang Y, Zhang JY, Zhang D, Zhang
PX, Jiang GY, Han Y, Zhang S, Cui QZ and Wang EH: Upregulation of
δ-catenin is associated with poor prognosis and enhances
transcriptional activity through Kaiso in non-small-cell lung
cancer. Cancer Sci. 102:95–103. 2011. View Article : Google Scholar
|
|
11
|
Sobin LH and Wittekind C: Colon and
rectum. TNM classification of malignant tumors. 6th edition. New
York: Wiley, John & Sons; pp. 72–76. 2002
|
|
12
|
Bosman FT, Carneiro F, Hruban RH and
Theise ND: WHO Classification of Tumors of the Digestive System.
4th edition. IARC press; Lyon: 2010
|
|
13
|
Bustin SA, Benes V, Garson JA, Hellemans
J, Huggett J, Kubista M, Mueller R, Nolan T, Pfaffl MW, Shipley GL,
et al: The MIQE guidelines: minimum information for publication of
quantitative real-time PCR experiments. Clin Chem. 55:611–622.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Schmittgen TD and Livak KJ: Analyzing
real-time PCR data by the comparative C (T) method. Nat Protoc.
3:1101–1108. 2008. View Article : Google Scholar
|
|
15
|
Lu Q, Paredes M, Medina M, Zhou J, Cavallo
R, Peifer M, Orecchio L and Kosik KS: delta-catenin, an adhesive
junction-associated protein which promotes cell scattering. J Cell
Biol. 144:519–532. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Jones SB, Lanford GW, Chen YH, Morabito M,
Kim K and Lu Q: Glutamate-induced delta-catenin redistribution and
dissociation from postsynaptic receptor complexes. Neuroscience.
115:1009–1021. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Martinez MC, Ochiishi T, Majewski M and
Kosik KS: Dual regulation of neuronal morphogenesis by a
delta-catenin-cortactin complex and Rho. J Cell Biol. 162:99–111.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kim K, Sirota A, Chen YH, Jones SB, Dudek
R, Lanford GW, Thakore C and Lu Q: Dendrite-like process formation
and cytoskeletal remodeling regulated by delta-catenin expression.
Exp Cell Res. 275:171–184. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Paffenholz R, Kuhn C, Grund C, Stehr S and
Franke WW: The arm-repeat protein NPRAP (neurojungin) is a
constituent of the plaques of the outer limiting zone in the
retina, defining a novel type of adhering junction. Exp Cell Res.
250:452–464. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lu Q, Mukhopadhyay NK, Griffin JD, Paredes
M, Medina M and Kosik KS: Brain armadillo protein delta-catenin
interacts with Abl tyrosine kinase and modulates cellular
morphogenesis in response to growth factors. J Neurosci Res.
67:618–624. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lu Q, Abdul A, Chen YH, et al: δ-Catenin
has the potential to promote the proliferation/survival and
invasiveness of human cancer cells. Mol Biol Cell. 14:3412003a.
|
|
22
|
Kim K, Oh M, Ki H, Wang T, Bareiss S, Fini
ME, Li D and Lu Q: Identification of E2F1 as a positive
transcriptional regulator for delta-catenin. Biochem Biophys Res
Commun. 369:414–420. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang T, Chen YH, Hong H, Zeng Y, Zhang J,
Lu JP, Jeansonne B and Lu Q: Increased nucleotide polymorphic
changes in the 5′-untranslated region of delta-catenin (CTNND2)
gene in prostate cancer. Oncogene. 28:555–564. 2009. View Article : Google Scholar :
|