Introduction
The lateral habenula (LHb) region of the brain, has
a critical role in controlling numerous behaviorally important
functions, including reward, emotional regulation, pain, stress,
sleep and cognition (1–3). The LHb is comprised of multiple
subnuclei and it is considered an integral neuronal component for
transmitting information between the limbic forebrain and the
midbrain regions. The LHb inhibits the activity of neurons that
contain monoaminergic neurotransmitters, including dopamine and
serotonin, via efferent projections that terminate in the
substantia nigra pars compacta, ventral tegmental area and raphe
nuclei (2–6). Dysfunction of the LHb is associated
with a number of neurological disorders, including sleep
disturbance, depression, drug addiction and schizophrenia (1,7–11).
A number of studies have demonstrated that neurons
in the LHb express high levels of estrogen receptor (ER)-α and
ER-β, which preferentially localizes to nuclei, axon terminals and
postsynaptic dendrites (12–15).
It has also been suggested that estrogen signaling in the LHb has
an essential role in regulating female reproductive behaviors
during pregnancy and post-parturition (14). Together, these findings support a
role for gonadal hormones in the LHb. Nevertheless, the sensitivity
of neurons in the LHb to hormonal regulation remains poorly
understood.
In the present study, the effects of estradiol
signaling on the LHb were investigated using a model of estradiol
replacement therapy in ovariectomized rats. The
electrophysiological responses of LHb neurons to estradiol, the
most biologically potent member of the estrogen family, were
evaluated.
Materials and methods
Animals
A total of 70 female Wistar rats aged 9–10 weeks,
were obtained from the Laboratory Animal Center of Jilin University
(Changchun, China; certification no. SCXK(Ji)2007-0003). Animals
were housed in standard temperature-, humidity- and
light-controlled conditions (temperature 22±1°C; humidity 40–50%;
lighting, 12 h light/dark cycle). All animals were housed in the
facility under these conditions for 1 week prior to experiments.
All experiments with animals were approved by the Animal Ethical
Committee of Jilin University and performed in accordance with
Decree No. 2 of the State and Technology Commission of China
(Approved by the State Council on October 31, 1988, and promulgated
by the State Science and Technology Commission on November 14,
1988).
Surgery and estradiol replacement
treatments
On day 0, bilateral ovariectomy of rats was
performed as previously described (16), with minor modifications.
Silastic® capsules (Dow Corning, Midland, MI, USA)
containing either 0, 90, 180 or 360 μg/ml of 17β-estradiol
(Sigma-Aldrich, St. Louis, MO, USA) in a solution with sesame oil
(Sigma-Aldrich) were implanted subcutaneously for the Oil, low
estradiol (LE), medial estradiol (ME) and high estradiol (HE)
groups, respectively. The Silastic® capsules were sealed
with Type A biogum (Dow Corning) and preincubated in sesame oil for
16 h. Following surgery, animals received daily intraperitoneal
injections of 40,000 units of penicillin for 3 days.
Evaluation of serum estradiol
content
Blood (1 ml) was collected from the tail vein at 3,
5, 7, 9, 11, 18, 26 and 33 days after surgery (17,18).
The samples were incubated for 1 h at room temperature and
centrifuged at 3,764 × g for 20 min. The serum was separated and
stored at 4°C for up to 24 h prior to serum estradiol
quantification by chemiluminescence using a UniCel DxI 800
Immunoassay System (Beckman Coulter, Brea, CA, USA).
Electrophysiology
Rats were anesthetized by intraperitoneal injection
of 10% chloral hydrate. The rats were then sacrificed by
decapitation and then LHb tissues were extracted for sample
preparation. Tissue sections containing the LHb regions were
dissected and 350 μm coronal slices were prepared. Slices
were immediately incubated in oxygenated artificial cerebrospinal
fluid (ACSF) (19) containing 135
mM NaCl, 3.5 mM KCl, 26 mM NaHCO3, 1.25 mM
NaH2PO4, 10 mM glucose, 2.5 mM
CaCl2 and 1.2 mM MgSO4 for 25 min at 34°C and
incubated at room temperature (20–25°C) for 1 h prior to patch
clamp analysis. For the duration of the patch clamp analysis
experiments, samples were perfused with oxygenated ACSF (2–3
ml/min) at 20–25°C and cells were continuously monitored using a
BX51WI Olympus microscope (Olympus, Tokyo, Japan). Patch
microelectrodes were guided stereotactically and pulled to a tip
resistance of 3–5 MΩ on a PP-83 micropipette puller (Narishige,
Tokyo, Japan) and filled with internal solution containing 125 mM
K-gluconate, 20 mM KCl, 10 mM HEPES, 5 mM EGTA, 4 mM Mg-ATP, 0.4 mM
NaGTP and 0.1 mM CaCl2. Estradiol (1 μM)
dissolved in dimethyl sulfoxide (DMSO) were administered into the
superfusion fluid within 5 min.
Signals were amplified using an EPC10 patch clamp
amplifier and digitized at 10 kHZ (HEKA Elektronik Dr. Schulze
GmbH, Pfalz, Germany) and data were collected using PatchMaster
v2x32 (HEKA Elektronik Dr. Schulze GmbH) software and analyzed with
FitMaster v2x65 (HEKA Elektronik Dr. Schulze GmbH).
Immunohistochemical analysis
Animals were anesthetized by intraperitoneal
injection of 20% urethane (6 ml/kg body weight). The hearts of the
animals were flushed with phosphate-buffered saline (PBS) and
perfused with a solution of 4% paraformaldehyde (PFA) in 0.1 M PBS
(pH 7.2–7.4). Brain tissue slices which contained LHb were fixed in
4% PFA for 24 h, dehydrated in a series of ethanol and embedded in
paraffin. Paraffin-embedded tissues were sectioned into 2 μm
thick coronal slices, c-Fos protein was detected using a rabbit
anti-rat primary antibody specific for c-Fos-specific antibody
(1:500 dilution; cat. no. ab102699; Abcam, Cambridge, England).
Immunostaining was detected using an UltraSensitive™ SP kit
according to the manufacture's instructions (KIT-9710; Fuzhou Maxim
Biotech Inc., Fujian, China). Briefly, samples were labeled with a
biotin-labeled secondary antibody (solution C, 50 μl) and
streptavidin-anti-biotin-peroxidase (solution D, 50 μl).
Immunostaining was visualized using 3,3′-diaminobenzidine
tetrahydrochloride kit (DAB-0031; Fuzhou Maxim Biotech Inc.).
Samples were counterstained with hematoxylin, washed with running
tap water, dehydrated in a series of ethanol, cleared in xylene and
mounted with neutral balsam.
Sections were subsequently counterstained with
hematoxylin prior to visualization on a BX53 microscope (Olympus,
Tokyo, Japan). The mean optical density of c-Fos staining in LHb
sections was determined by analysis of randomly selected fields and
quantification with Image-Pro Plus 6.0 software (Media Cybernetics,
Silver Spring, MD, USA).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was isolated from the tumor tissues using
TRIzol reagent according to the manufacturer's instructions
(Invitrogen Life Technologies, Carlsbad, CA, USA). First-strand
cDNA was synthesized by reverse transcription of 2 μg total
RNA using an RNA PCR kit (Takara Bio., Inc., Shiga, Japan). Primer
sequences used for RT-qPCR reactions were as follows: c-fos,
forward 5′-AGCATGGGCTCCCCTGTCA-3′ and reverse
5′-GAGACCAGAGTGGGCTGCA-3′; GAPDH, forward
5′-GGGTGATGCTGGTGCTGAGTATGT-3′ and reverse
5′-AAGAATGGGAGTTGCTGTTGAAGT-3′.
Reactions were run in an ABI PRISM 7900 HT
thermocycler (Applied Biosystems, Foster City, CA, USA) for 35
cycles. The relative mRNA expression of the target gene was
normalized to that of GAPDH. Data were analyzed from three
independent experiments (three animals).
Western blot analysis
The LHb tissue samples were lysed in RIPA solution
for 30 min on ice. Lysates were centrifuged at 12,000 × g for 10
min at 4°C and the supernatants were collected and stored at −80°C.
Protein concentrations for each sample were determined using a
Bradford assay according to the manufacturer's instructions
(Nanjing Jiancheng Biotechnology Institute, Nanjing, China).
Protein (40 μg) from each sample was separated in an 8 or
12% sodium dodecyl sulfate polyacrylamide gel and transferred onto
a polyvinylidene difluoride membrane (Millipore, Billerica, MA,
USA) at 4°C. Proteins were detected with a rabbit anti-rat
c-Fos-specific antibody (1:1,000 dilution; cat. no. ab102699;
Abcam, Cambridge, UK), horseradish peroxidase-conjugated goat
anti-rabbit IgG secondary antibodies (1:6,000 dilution; ZDR-5118;
Beijing Zhongshan Golden Bridge Biotechnology Co., Ltd., Beijing,
China) and visualized using enhanced chemiluminescence solution and
X-ray films. Band densities were determined by Scion version 4.02
imaging software (Scion Corporation, Frederick, MD, USA).
Statistical analysis
Data were analyzed by SPSS 11.0 software (SPSS,
Inc., Chicago, IL, USA) and are presented as the mean ± standard
error of the mean. Statistical significance was determined using
one-way analysis of variance and two-tailed t-test and displayed
using GraphPad Prism 6 software (GraphPad Software, Inc., San
Diego, CA, USA).
Results
Estradiol replacement therapy is
sustained for up to 3 weeks in ovariectomized rats
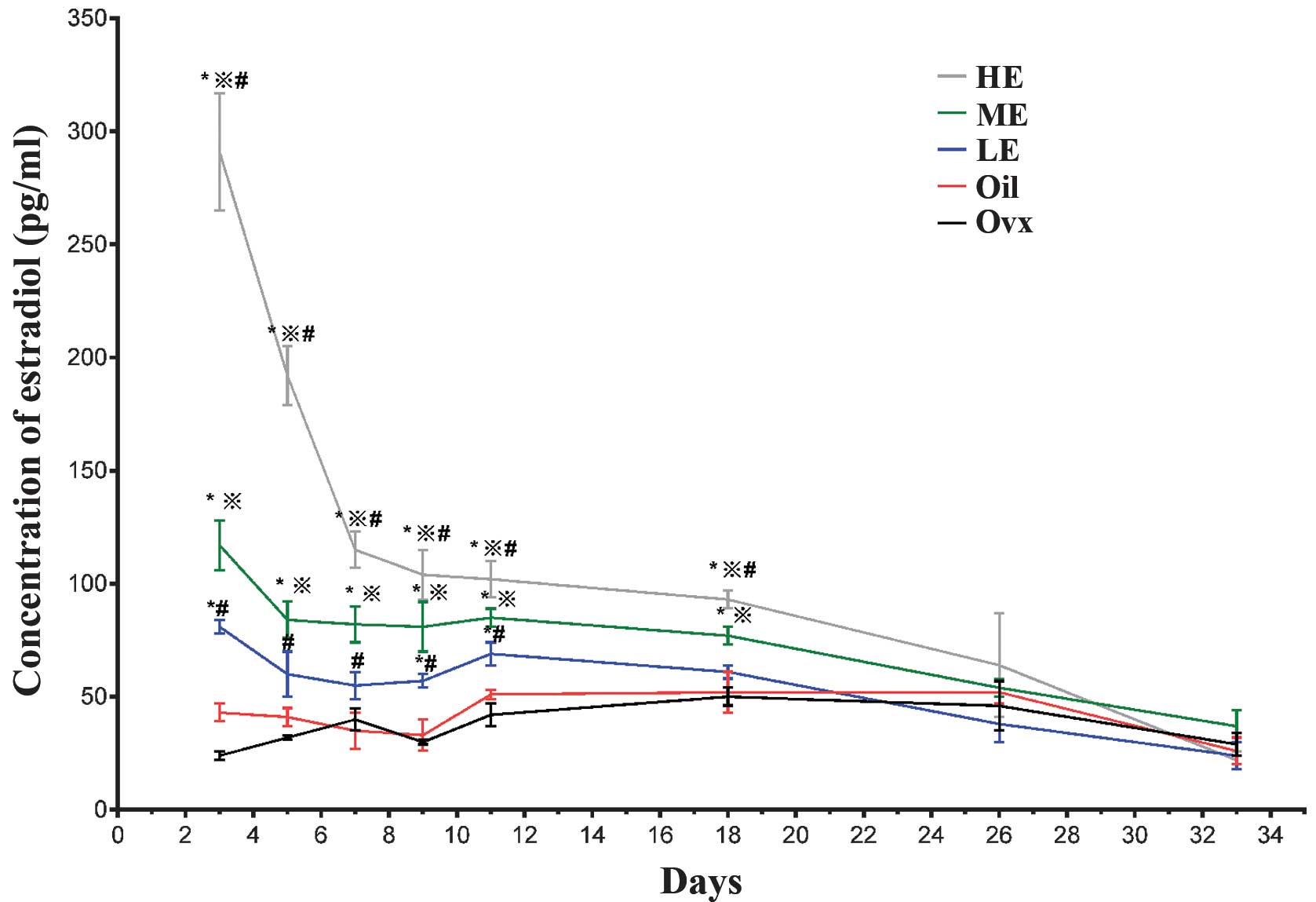
No significant differences were found in the serum
estradiol levels of untreated ovariectomized rats when compared
with those in the Oil group (P>0.05; Fig. 1), which demonstrated that the
sesame oil carrier solution had no biological effects in this
system. Serum estradiol levels in the medial estradiol (ME) and
high estradiol (HE) groups were significantly elevated at 3, 5, 7,
9, 11 and 18 days post-surgery compared with the Oil group
(P<0.05; Fig. 1). Animals in
the low estradiol (LE) dosage group exhibited increased serum
estradiol levels only at 9 and 11 days post-surgery (P<0.05;
Fig. 1). However, no statistically
significant differences were found in serum estradiol levels for
any group compared with the controls at 26 and 33 days post-surgery
(P>0.05; Fig. 1). These
findings demonstrated that hormone replacement therapy in
ovariectomized rats was effective for up to 3 weeks. Given that
animals in the ME group appeared to maintain serum estradiol levels
for the longest period, these animals were used to investigate the
effects of estrogen hormones on c-Fos expression in the LHb
neurons.
Estradiol suppresses spontaneous firing
activity in LHb neurons
The effects of estradiol (dissolved in DMSO) on the
functional activity of neurons in the LHb derived from normal
female rats were examined using patch clamp electrophysiology
techniques. Administration of a 0.01–0.05% DMSO solution to LHb
tissue samples demonstrated no effect on the spontaneous firing
rate of neurons in the LHb area. However, exposure of LHb tissue
samples to 1 μM estradiol, resulted in a 40.89% decrease in
the average firing rate of the neurons from 4.33±0.90 Hz prior to
treatment with estradiol to 2.65±0.63 Hz following administration
of estradiol (P<0.01; Fig.
2A–C). When the estradiol-treated samples were washed for 5 min
with ACSF, the firing rate was recovered to baseline levels
(Fig. 2A and B). Together these
results suggest that estradiol inhibits the spontaneous firing
activities of neurons in the LHb region and that this effect is
reversible when estrogen is attenuated.
Expression of c-Fos in the LHb of
ME-treated rats
The proto-oncogene c-Fos is a widely accepted marker
of neuron activation within the central nervous system. To
understand the effect of estradiol on LHb neurons, the expression
of c-Fos was examined at the mRNA and protein levels in ME-treated
and control animals. The mRNA expression of c-Fos was significantly
decreased in ME-treated animals compared with that of the control
animals 7 days post-surgery (P<0.05; Fig. 3A). These results were supported by
similar findings at the protein level that demonstrated reduced
quantities of c-Fos protein in ME-derived LHb tissues compared with
the equivalent control tissues (P<0.05; Fig. 3B and C).
Immunohistochemical analysis of tissue sections
stained for c-Fos revealed that the intensity of staining as well
as the number of c-Fos-positive neurons was decreased in the LHb
region of ME rats compared with Oil animals (P<0.05; Fig. 4A and B).
Discussion
The LHb region is known to regulate a wide range of
biological functions, including reward and aversion (1), and dysfunction in LHb neurons is
associated with numerous neurological disorders, including sleep
disturbance and depression (9).
Estradiol regulates numerous physiological functions in the central
nervous system, and changes in the levels of estrogen are
associated with the pathogenesis of psychiatric illness (20–22).
The present study investigated the association between estrogen
hormones and neuronal activity in the LHb region and demonstrated
that this hormone functions to inhibit neuron activity in the LHb
region.
The ovariectomized female rat has been widely used
as an ideal rodent model to investigate the effect of hormone
replacement on pain responses and analgesia (23,24).
To understand the effects of estradiol on neuronal activities in
the LHb area, estradiol replacement therapy was applied in
ovariectomized female rats to avoid the effects of endogenous
hormones on the outcome. It was found that subcutaneous
implantation of Silastic® capsules containing estradiol
provided sustained serum hormone levels for ~3 weeks after
implantation. In addition, no significant differences were found in
serum estradiol levels between animals in the Oil group and in the
ovariectomized group receiving no treatment, indicating that the
sesame carrier solution did not affect the results.
Patch clamp recording analysis demonstrated that
exogenous estradiol inhibited spontaneous firing activities in LHb
neurons, which suggested a role for this hormone in negative
regulation in the LHb. The proto-oncogene c-Fos is a marker of
activated neurons and is typically transiently induced in response
to multiple extracellular stimuli (25,26).
The present study demonstrated that estradiol replacement therapy
resulted in a significant decrease in c-Fos expression in LHb
tissues.
Our results suggest that estradiol may suppress
neuronal firing activity and c-Fos expression in the LHb, however,
additional investigations are required to clarify the precise
mechanisms underlying its action on LHb neurons.
In conclusion, the present study demonstrated that
estradiol hormone inhibits neuronal activity in the LHb region.
These results may provide a rationale for the development of
estradiol pharmacological interventions for neurological disorders,
via effects on the LHb neuronal activity.
Acknowledgments
This study was supported by the Bethune Medical
Scientific Research Support Plan (grant no. 2013205029) and Beihua
University doctoral research fund (Miss Chun-ying Li).
References
|
1
|
Zhao H, Zhang BL, Yang SJ and Rusak B: The
role of lateral habenula-dorsal raphe nucleus circuits in higher
brain functions and psychiatric illness. Behav Brain Res.
277:89–98. 2015. View Article : Google Scholar
|
|
2
|
Christoph GR, Leonzio RJ and Wilcox KS:
Stimulation of the lateral habenula inhibits dopamine-containing
neurons in the substantia nigra and ventral tegmental area of the
rat. J Neurosci. 6:613–619. 1986.PubMed/NCBI
|
|
3
|
Hikosaka O: The habenula: From stress
evasion to value-based decision-making. Nat Rev Neurosci.
11:503–513. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wang RY and Aghajanian GK: Physiological
evidence for habenula as major link between forebrain and midbrain
raphe. Science. 197:89–91. 1977. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Matsumoto M and Hikosaka O: Lateral
habenula as a source of negative reward signals in dopamine
neurons. Nature. 447:1111–1115. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bernard R and Veh RW: Individual neurons
in the rat lateral habenular complex project mostly to the
dopaminergic ventral tegmental area or to the serotonergic raphe
nuclei. J Comp Neurol. 520:2545–2558. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ellison G: Stimulant-induced psychosis,
the dopamine theory of schizophrenia and the habenula. Brain Res
Brain Res Rev. 19:223–239. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lecourtier L, Neijt HC and Kelly PH:
Habenula lesions cause impaired cognitive performance in rats:
Implications for schizophrenia. Eur J Neurosci. 19:2551–2560. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Aizawa H, Cui W, Tanaka K and Okamoto H:
Hyperactivation of the habenula as a link between depression and
sleep disturbance. Front Hum Neurosci. 7:8262013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sartorius A, Kiening KL, Kirsch P, et al:
Remission of major depression under deep brain stimulation of the
lateral habenula in a therapy-refractory patient. Biol Psychiatry.
67:e9–e11. 2010. View Article : Google Scholar
|
|
11
|
Li B, Piriz J, Mirrione M, et al: Synaptic
potentiation onto habenula neurons in the learned helplessness
model of depression. Nature. 470:535–539. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Shughrue PJ, Lane MV and Merchenthaler I:
Comparative distribution of estrogen receptor-alpha and -beta mRNA
in the rat central nervous system. J Comp Neurol. 388:507–525.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Laflamme N, Nappi RE, Drolet G, Labrie C
and Rivest S: Expression and neuropeptidergic characterization of
estrogen receptors (ERalpha and ERbeta) throughout the rat brain:
Anatomical evidence of distinct roles of each subtype. J Neurobiol.
36:357–378. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wagner CK, Silverman AJ and Morrell JI:
Evidence for estrogen receptor in cell nuclei and axon terminals
within the lateral habenula of the rat: Regulation during
pregnancy. J Comp Neurol. 392:330–342. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Milner TA, Thompson LI, Wang G, et al:
Distribution of estrogen receptor beta containing cells in the
brains of bacterial artificial chromosome transgenic mice. Brain
Res. 1351:74–96. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Deurveilher S, Rusak B and Semba K:
Estradiol and progesterone modulate spontaneous sleep patterns and
recovery from sleep deprivation in ovariectomized rats. Sleep.
32:865–877. 2009.PubMed/NCBI
|
|
17
|
Frank P, Schoenhard GL and Burton E: A
method for rapid and frequent blood collection from the rat tail
vein. J Pharmacol Methods. 26:233–238. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Staszyk C, Bohnet W, Gasse H and Hackbarth
H: Blood vessels of the rat tail: A histological re-examination
with respect to blood vessel puncture methods. Lab Anim.
37:121–125. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sun J, Chu Z and Moenter SM: Diurnal in
vivo and rapid in vitro effects of estradiol on voltage-gated
calcium channels in gonadotropin-releasing hormone neurons. J
Neurosci. 30:3912–3923. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gutiérrez-Lobos K, Scherer M, Anderer P
and Katschnig H: The influence of age on the female/male ratio of
treated incidence rates in depression. BMC Psychiatry. 2:32002.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li R and Shen Y: Estrogen and brain:
Synthesis, function and diseases. Front Biosci. 10:257–267. 2005.
View Article : Google Scholar
|
|
22
|
Soares CN: Mood disorders in midlife
women: Understanding the critical window and its clinical
implications. Menopause. 21:198–206. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Fillingim RB and Ness TJ: Sex-related
hormonal influences on pain and analgesic responses. Neurosci
Biobehav Rev. 24:485–501. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Mannino CA, South SM, Quinones-Jenab V and
Inturrisi CE: Estradiol replacement in ovariectomized rats is
antihyperalgesic in the formalin test. J Pain. 8:334–342. 2007.
View Article : Google Scholar
|
|
25
|
Harris JA: Using c-fos as a neural marker
of pain. Brain Res Bull. 45:1–8. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kovács KJ: Measurement of immediate-early
gene activation - c-fos and beyond. J Neuroendocrinol. 20:665–672.
2008. View Article : Google Scholar
|