Introduction
Vascular smooth muscle cells (VSMCs) are an
important constituent of vessel walls, and their dysfunction may
lead to pathological processes and cause vascular disease. VSMCs
undergo cell biological changes in response to a variety of
cytokines and growth factors (1).
Adiponectin is an important adipocyte-derived
hormone (2,3), which is well-known for its
involvement in lipid and glucose metabolism and insulin sensitivity
(4–7). Numerous studies have suggested that
adiponectin has potent anti-inflammatory (8), athero-protective (9), antihypertensive (10) and antidiabetic (11) effects. A previous study
demonstrated that adiponectin is also expressed and secreted by
VSMCs (12), and adiponectin
inhibits neointimal formation through suppression of the
proliferation and migration of VSMCs (13–15).
However, the mechanisms underlying the effect of adiponectin on
VSMC proliferation and apoptosis require further investigation.
The present study aimed to investigate whether
mitofusin-2 (MFN2) which mediates the effect of adiponectin on VSMC
proliferation and apoptosis, which may provide a novel basis for
the therapy of vascular disease.
Materials and methods
Cell culture
The Aortic Smooth Mus cells (VSMCs; cat. no.
CC-2571) were purchased from Clonetics (San Diego, CA, USA). The
VSMCs were cultured in Dulbecco's modified Eagle's medium
(Gibco-BRL, Rockville, MD, USA) with 20% fetal bovine serum
(Gibco-BRL) in a humidified atmosphere at 37°C of 5%
CO2. The culture medium was replaced every 3 days.
Treatment of VSMCs with adiponectin
Adiponectin was purchased from Cayman Chemical
Company (Ann Arbor, MI, USA) and dissolved in phosphate-buffered
saline (1 mg/ml). To obtain different experimental concentrations
(5, 10, 20 and 40 µg/ml), serial dilutions were prepared in
the culture medium. VSMCs (5×105 cells/ml) were seeded
into 6-well plates and when the VSMCs reached 80% confluence,
adiponectin was added to treat the cells for 24 h at 37°C.
Transfection
The cells were seeded into 6-well plates at a
density of 3×105 cells/well and allowed to grow for 24
h. MFN2 small interfering (si)RNA, forward
5′-CCAUGAGGCCUUUCUCCTT-3′ and reverse 5′-GGAGAAAGGCCUCAUGGTT-3′; or
control siRNA, forward 5′-UUCUCCGAACGUGUCACGUTT-3′ and reverse
5′-ACGUGACACGUUCGGAGAATT-3′. were synthesized at Sangon Biotech
Inc. (Shanghai, China) and transfected into the cells at a final
concentration of 2 µM using lipofectamine 2000 transfection
reagent (Invitrogen Life Technologies, Carlsbad, CA, USA),
according to the manufacturer's instructions.
Flow cytometry assay
For cell apoptosis analysis using flow cytometry,
the cells were harvested and fixed in 70% ethanol (Xinyu Inc.,
Shanghai, China) on ice. Following centrifugation at 1,000 × g for
5 min at room temperature, the cells were stained with annexin V
and propidium iodide (Kaiji Biological Inc., Nanjing, China),
according to the manufacturer's instructions, and analyzed using
flow cytometry (FC 500 MPL system; Beckman Coulter, Inc., Miami,
FL, USA).
MTT assay
The cells were seeded into 96-well plates at a
density of 4×103 cells/well. Following treatment with
adiponectin for 24, 48 and 72 h, 10 µl MTT solution
(Sigma-Aldrich, St. Louis, MO, USA) was added to each well, and the
incubation was continued for 4 h at 37°C. The formazan was
solubilized in dimethyl sulfoxide (Sigma-Aldrich) and the viable
cells were determined by measuring the absorbance at 570 nm using a
microplate reader (Multiskan Ascent 354; Thermo Labsystems,
Waltham, MA, USA).
Western blot analysis
The proteins from cultured cells were extracted
using cell lysis buffer, containing 50 mM Tris (Amresco LLC, Solon,
OH, USA), 150 mM NaCl (Tianjin Dingshengxin Chemical Industry Co.,
Ltd., Tianjin, China), 1 mM EDTA (Zhiyuan, Inc., Tianjin, China),
1% Triton X-100 (Amresco LLC), 1 mM sodium orthovanadate (Beyotime
Institute of Biotechnology, Shanghai, China), 1 mM PMSF (Amresco
LLC) and 2 mM DTT (pH 7.4; Amresco LLC). Protein concentrations
were quantified using a bicinchoninic acid assay method, using
reagents purchased from Pierce Biotechnology, Inc. (Rockford, IL,
USA). Equal quantities of the cell lysates were separated on 12%
SDS-PAGE (Invitrogen Life Technologies, Carlsbad, CA, USA), and
then transferred on to polyvinylidene difluoride membranes
(Millipore, Billerica, MA, USA) via electroblotting. The membranes
were blocked with Tris-buffered saline containing 5% bovine serum
albumin (Sigma-Aldrich) at 37°C for 1 h. The blot was then probed
with mouse monoclonal antibody against MFN2 (sc-100560; 1:500;
Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA), mouse
monoclonal antibody against Ras (sc-166691; 1:800; Santa Cruz
Biotechnology, Inc.), mouse monoclonal antibody against c-Raf
(#12552; 1:400; Cell Signaling Technology, Inc., Beverly, MA, USA),
rabbit monoclonal antibody against extracellular signal regulated
kinase (Erk)1/2 (#4695; 1:400; Cell Signaling Technology, Inc.),
rabbit monoclonal antibody against phosphorylated (p)-c-Raf
(Ser338; #9427; 1:400; Cell Signaling Technology, Inc.) and rabbit
monoclonal antibody against p-Erk1/2 (Thr202/Tyr204; #4377; 1:400;
Cell Signaling Technology, Inc.). Subsequently, the membranes were
incubated with horseradish peroxidase (HRP)-conjugated secondary
antibody [goat anti-rabbit IgG/HRP (sc-2004; 1:10,000) and goat
anti-mouse IgG/HRP (sc-2031; 1:10,000); Santa Cruz Biotechnology,
Inc.)] at 37°C for 1 h. The signals were detected using an enhanced
chemiluminescence western blotting kit (Pierce Biotechnology,
Inc).
Statistical analysis
All data are expressed as the mean ± standard
deviation of at least three independent experiments. Comparisons of
the parameters between the two groups were performed using
Student's t-test. SPSS software, version 19 (IBM SPSS, Armonk, NY,
USA) was used for statistical analysis. P<0.05 was considered to
indicate a statistically significant difference.
Results
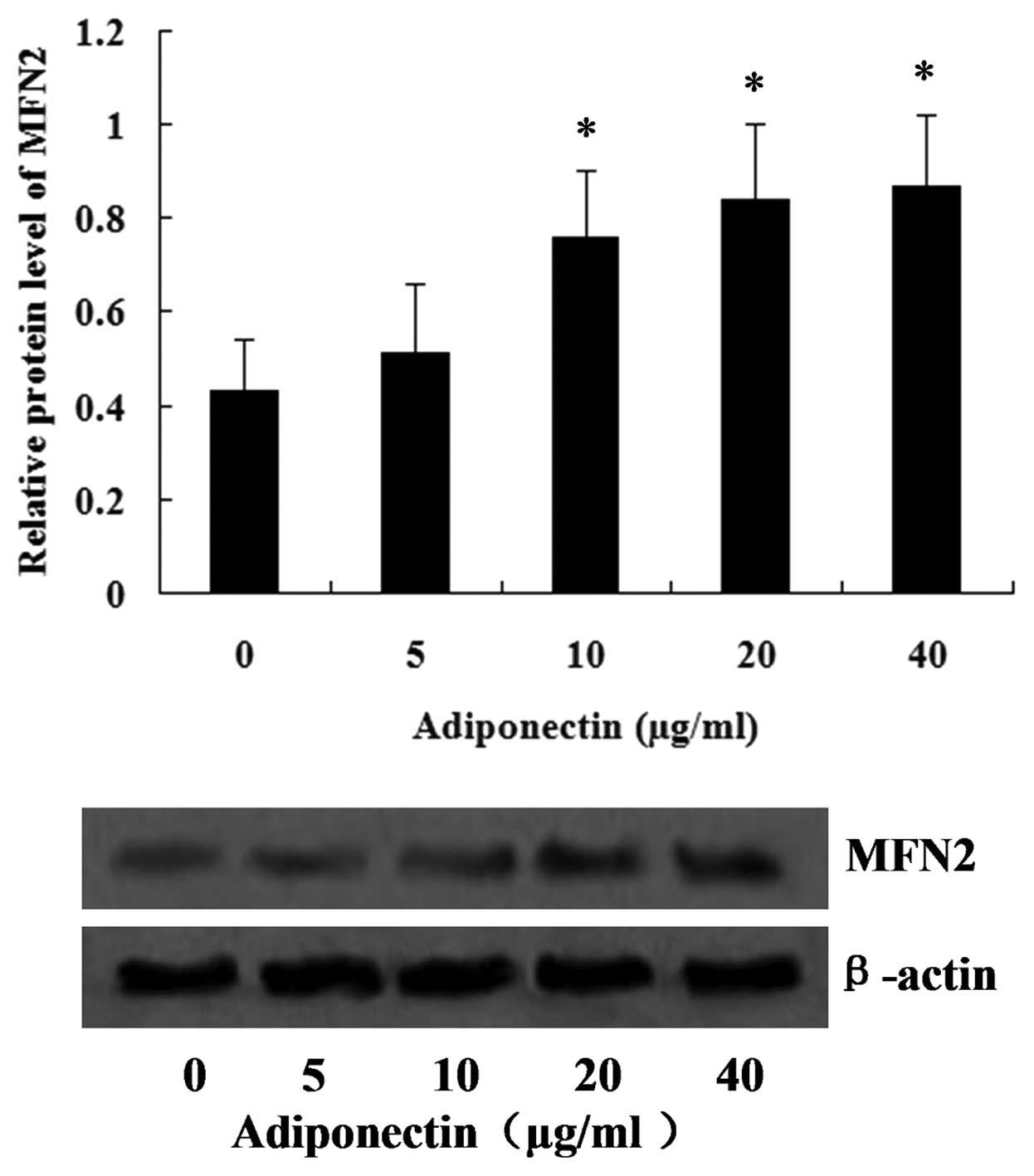
Adiponectin induces the expression of
MFN2 in VSMCs
To investigate the effect of adiponectin on the
expression of MFN2, the VSMCs were treated with adiponectin at
concentrations between 5 and 40 µg/ml, and, after 24 h, the
expression of MFN2 in the VSMCs was assessed using western blot
analysis. The results are shown in Fig. 1, and revealed that the expression
of MFN2 increased with increasing concentrations of adiponectin.
Concentrations between 10 and 40 µg/ml adiponectin
significantly increased the expression of MFN2, with the peak value
observed at 40 µg/ml (P<0.05).
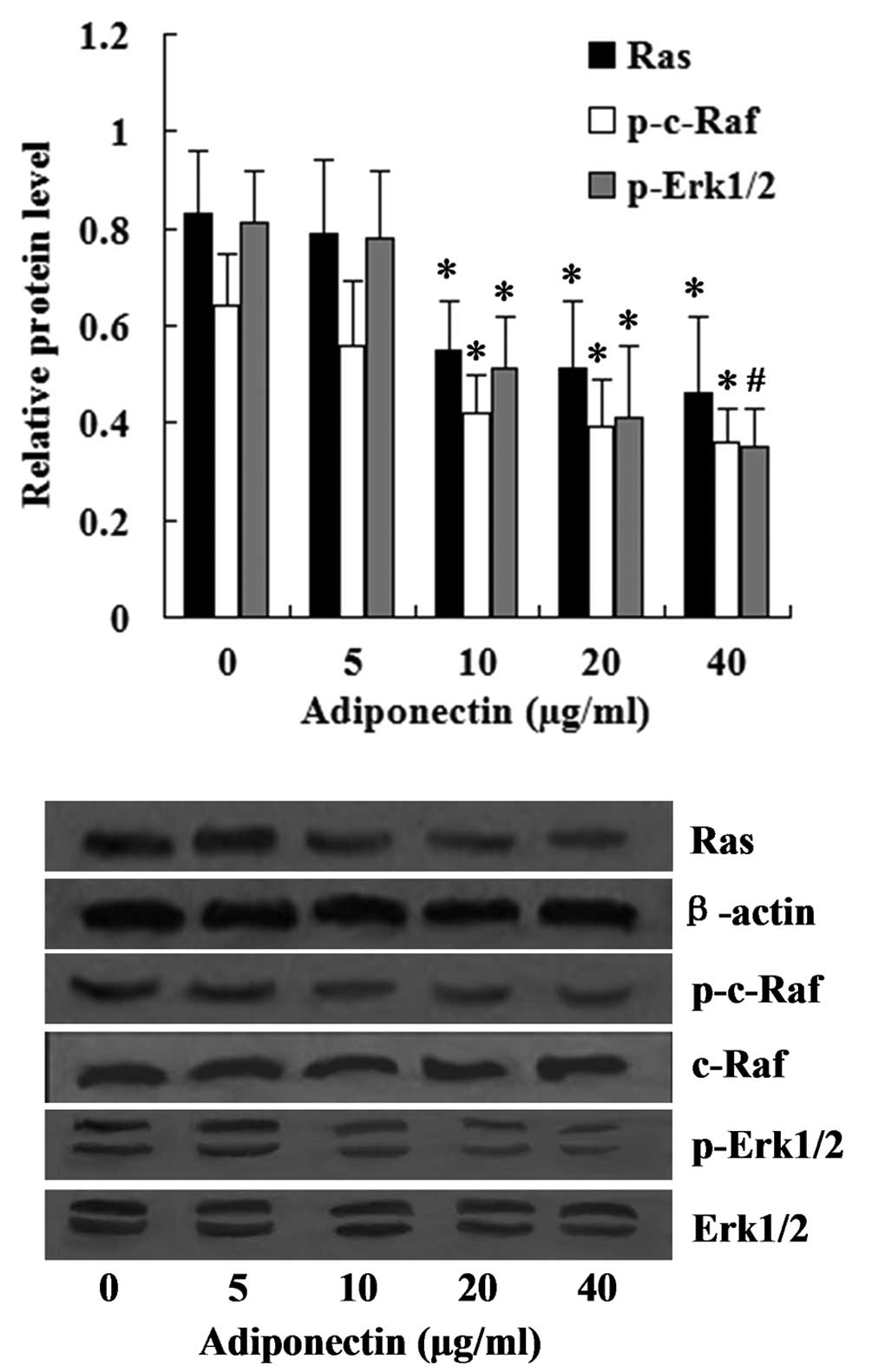
Inhibition of the Ras-Raf-Erk1/2
signaling pathway following treatment with adiponectin
The Ras-Raf-Erk1/2 pathway is one of the most
important downstream signaling cascades of MFN2. The effect of
adiponectin on the expression levels of Ras, p-c-Raf and p-Erk1/2
was investigated in the present study. The relative expression
level of Ras was normalized to that of β-actin, while the relative
expression levels of the phosphorylated forms of c-Raf and Erk1/2
were normalized to the levels of total c-Raf and Erk1/2. The
western blot analysis revealed that the relative expression levels
of Ras, p-c-Raf, p-Erk1/2 in the VSMCs were significantly inhibited
by adiponectin at concentrations between 10 and 40 µg/ml
(P<0.05) (Fig. 2).
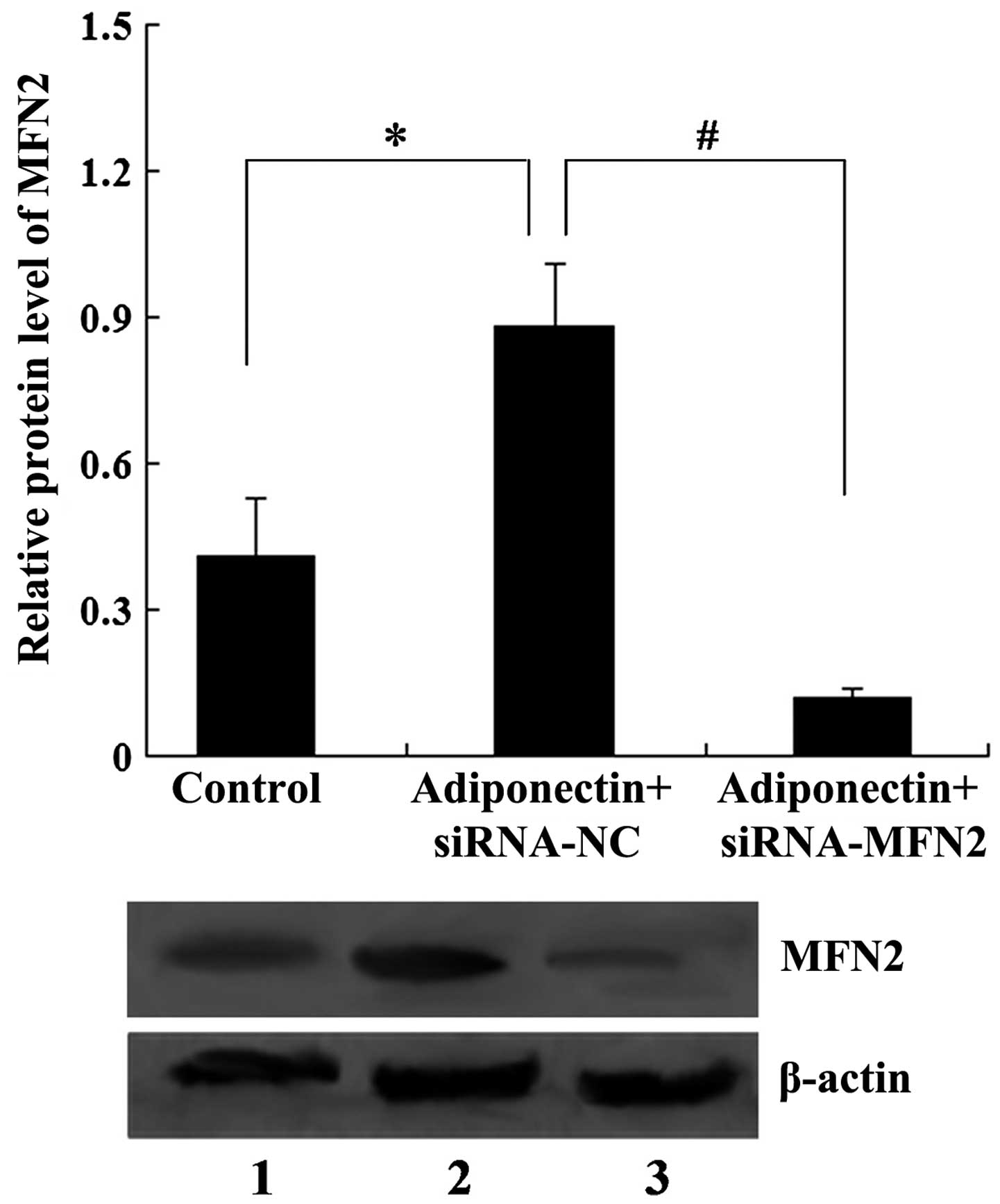
MFN2 knockdown attenuates the inhibitory
effect of adiponectin on VSMC proliferation
To examine whether MFN2 was involved in the effect
of adiponectin on VSMC proliferation, siRNA-MFN2 was transfected
into VSMCs for 24 h to knock down the expression of MFN2, following
which the VSMCs were treated with 40 µg/ml adiponectin for
another 24 h. The results revealed that 40 µg/ml adiponectin
upregulated the expression of MFN2 in the VSMCs (P<0.05).
However, the expression ofMFN2 was suppressed by transfection with
siRNA-MFN2, with the expression of MFN2 significantly decreased in
the adiponectin + siRNA-MFN2 group, compared with the adiponectin +
siRNA-control group (P<0.01; Fig.
3).
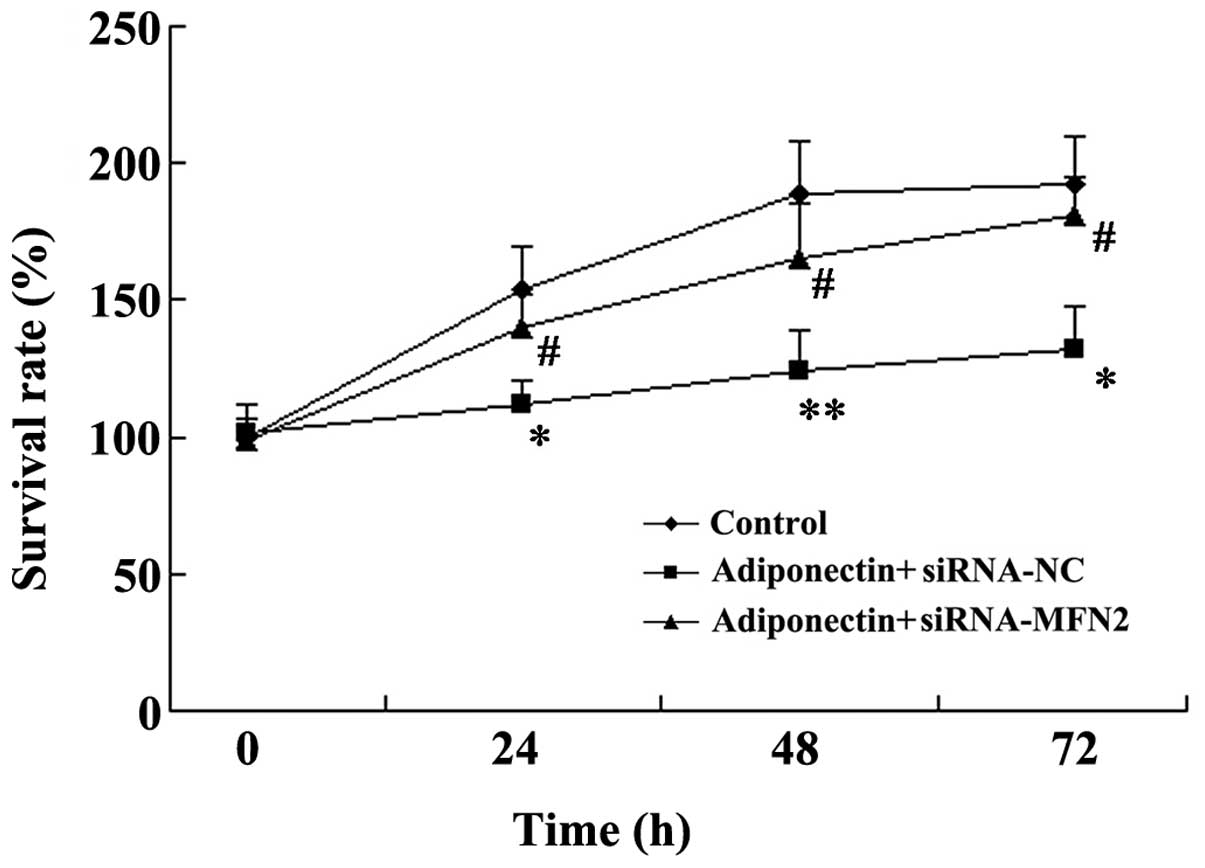
An MTT assay was used to examine VSMC proliferation.
As shown in Fig. 4, the VSMC
survival rates decreased significantly following treatment with
adiponectin (P<0.05). However, MFN2-knockdown attenuated the
inhibitory effect of adiponectin on VSMC proliferation, and the
VSMC survival rates were significantly increased in the adiponectin
+ siRNA-MFN2 group, compared with the adiponectin + siRNA-control
group (P<0.05; Fig. 4).
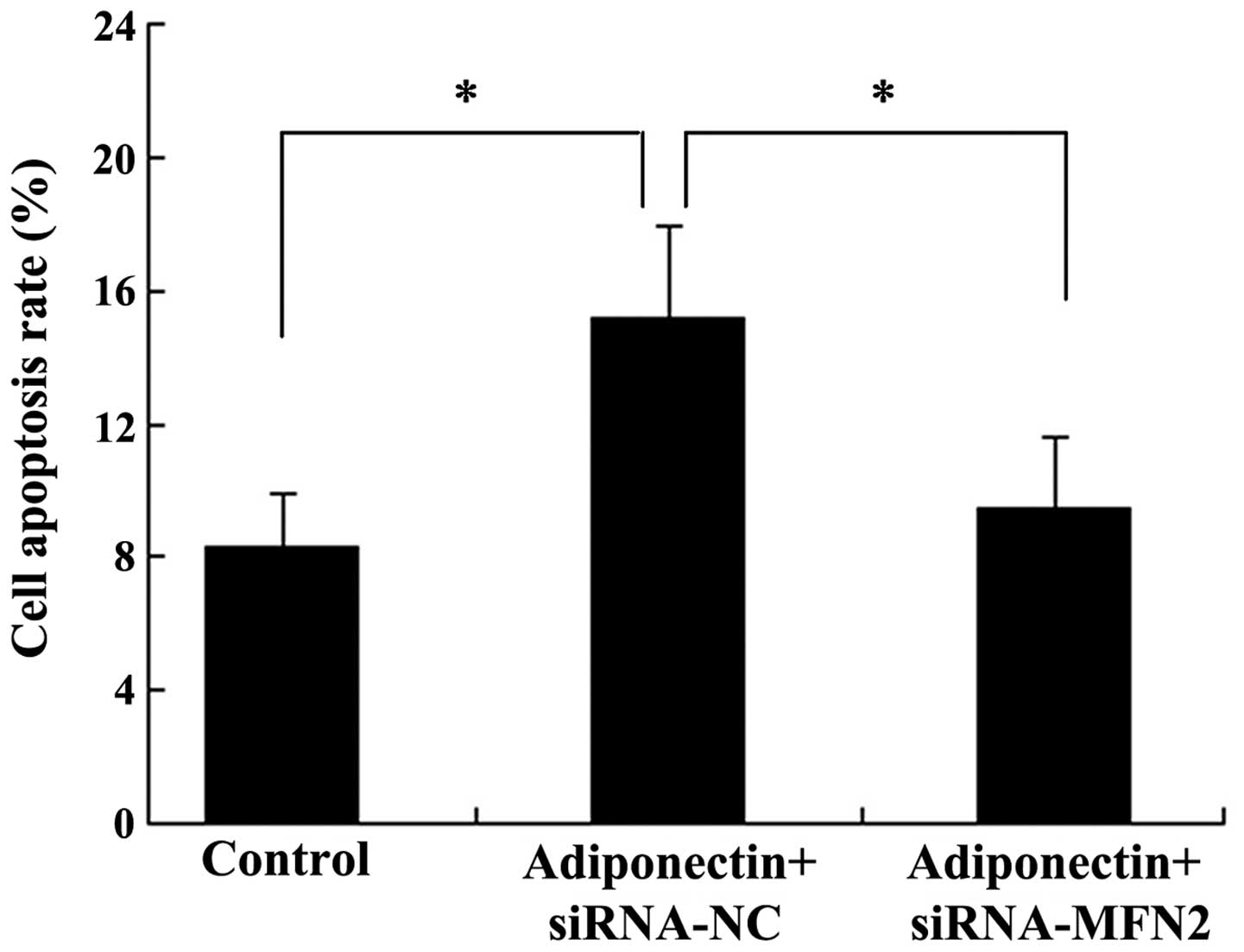
MFN2 knockdown attenuates
adiponectin-induced apoptosis in VSMCs
Subsequently, the present study examined whether
MFN2 was involved in the effect of adiponectin on VSMC apoptosis.
The VSMCs were transfected either with siRNA-MFN2 or siRNA-control
for 24 h, followed by treatment with 40 µg/ml adiponectin
for another 24 h. Subsequently, the VSMCs were harvested for flow
cytometric analysis. As shown in Fig.
5, the apoptotic rate of the cells in the control group was
8.3±1.6%, whilst treatment with adiponectin significantly induced
cell apoptotic rates to 15.2±2.7% (P<0.05). The apoptotic rate
of the cells in the adiponectin + siRNA-MFN2 group (9.5±2.1%) was
significantly different from the adiponectin + siRNA-control group
(15.2±2.7%) (P<0.05).
MFN2 knockdown attenuates the inhibitory
effect of adiponectin on the Ras-Raf-Erk1/2 signaling pathway
Western blot analysis was used to assess the role of
MFN2-knockdown in the inhibitory effect of adiponectin on the
Ras-Raf-Erk1/2 signaling pathway. As shown in Fig. 6, adiponectin significantly
inhibited the relative expression levels of Ras (P<0.05),
p-c-Raf (P<0.01) and p-Erk1/2 (P<0.01) in the VSMCs. However,
MFN2-knockdown attenuated the inhibitory effect of adiponectin, and
the relative expression levels of Ras, p-c-Raf and p-Erk1/2 were
significantly increased in the adiponectin + siRNA-MFN2 group,
compared with the adiponectin + siRNA-control group
(P<0.05).
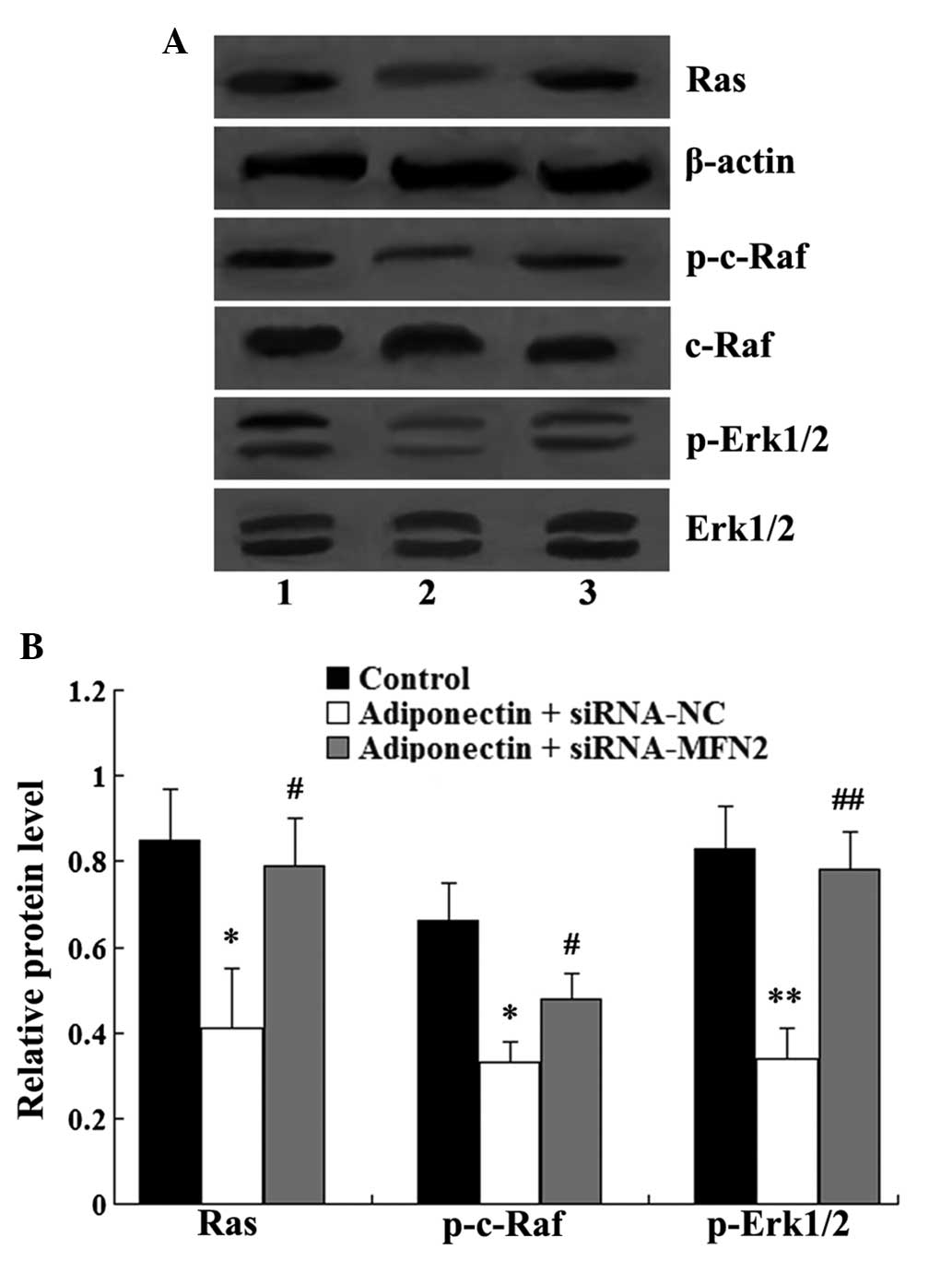 | Figure 6MFN2 knockdown attenuates the
inhibitory effect of adiponectin on the Ras-Raf-Erk1/2 signaling
pathway. (A) Western blot analysis of Ras, c-Raf, Erk1/2, p-c-Raf
and p-Erk1/2 in the VSMCs. Lane 1, control group; lane 2,
adiponectin + siRNA-NC group; lane 3, adiponectin + siRNA-MFN2
group. (B) Relative protein levels of Ras, p-c-Raf and p-Erk1/2.
The relative expression level of Ras was normalized to β-actin,
while the relative expression levels of p-c-Raf and p-Erk1/2 were
normalized to total c-Raf and Erk1/2. Data are expressed as the
mean ± standard deviation. *P<0.05 and
**P<0.01, compared with the control group;
#P<0.05 and ##P<0.01, compared with the
adiponectin + siRNA-NC group. VSMC, vascular smooth muscle cell;
siRNA, small interfering RNA; MFN2, mitofusin-2; NC, negative
control; ERK, extracellular signal-regulated kinase; p-,
phosphorylated. |
Discussion
It has been previously demonstrated that adiponectin
is involved in the protection of vascular injury by suppressing
neointimal formation (16,17). The present study demonstrated that
adiponectin exhibited an inhibitory effect on VSMC proliferation
and induced cell apoptosis, and these findings were consistent with
those of previous studies (13–15,18).
As mentioned previously, certain genes and signaling
pathways have been implicated in the effect of adiponectin on cell
growth (19–22). In the present study, MFN2 and its
downstream signaling cascade, the Ras-Raf-Erk1/2 signaling pathway,
were investigated.
MFN2 is a mitochondrial membrane protein, which is
primarily involved in mitochondrial fusion (23). MFN2 acts as a suppressor in cell
proliferation and as a promoter of cell apoptosis (24–28).
It was revealed in the present study that siRNA-mediated
downregulation of MFN2 in the VSMCs attenuated the effect of
adiponectin on VSMC proliferation and apoptosis, therefore the
results demonstrated that MFN2 was involved in the effect of
adiponectin on VSMC proliferation and apoptosis. In addition, it
was revealed that adiponectin significantly increased the
expression of MFN2 in a concentration-dependent manner.
Previous studies have indicated that the
Ras-Raf-Erk1/2 signaling pathway is the downstream signaling
cascade of MFN2 (29,30). Following activation of Ras, it
couples with Raf directly and the cellular Raf protein is
phosphorylated, the downstream signaling transduction pathways are
then activated, leading to the phosphorylation of Erk1 and Erk2.
The activated Erk1/2 protein contributes to the growth of VSMCs
(31,32). However, MFN2 acts as a negative
regulator of the Ras-Raf-Erk1/2 signaling pathway through its
interaction with Ras (29). In the
present study, the results demonstrated that, in the VSMCs,
adiponectin significantly inhibited the expression levels of Ras,
p-c-Raf and p-Erk1/2, however, MFN2 knockdown attenuated the
inhibitory effect of adiponectin. This demonstrated that MFN2
mediated the effects of adiponectin on the Ras-Raf-Erk1/2 signaling
pathway.
In conclusion, the present study demonstrated for
the first time, to the best of our knowledge, that adiponectin
exhibited an inhibitory effect on VSMC proliferation, and induced
cell proliferation via regulation of the expression of MFN2.
Adiponectin also upregulated the expression of MFN2, which
inhibited the Ras-Raf-Erk1/2 signaling pathway, leading to the
inhibition of VSMC proliferation and to the induction of VSMC
apoptosis. The results of the present study improves and expands
the current understanding of the effects of adiponectin on VSMCs
and may provide novel targets for therapeutic intervention in
vascular disease.
References
|
1
|
Kiyan Y, Limbourg A, Kiyan R, et al:
Urokinase receptor associates with myocardin to control vascular
smooth muscle cells phenotype in vascular disease. Arterioscler
Thromb Vasc Biol. 32:110–122. 2012. View Article : Google Scholar
|
|
2
|
Matsuzawa Y, Funahashi T and Nakamura T:
Molecular mechanism of metabolic syndrome X: contribution of
adipocytokines adipocyte-derived bioactive substances. Ann N Y Acad
Sci. 892:146–154. 1999. View Article : Google Scholar
|
|
3
|
Henry BA and Clarke IJ: Adipose tissue
hormones and the regulation of food intake. J Neuroendocrinol.
20:842–849. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yamauchi T, Kamon J, Waki H, et al: The
fat-derived hormone adiponectin reverses insulin resistance
associated with both lipoatrophy and obesity. Nat Med. 7:941–946.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Berg AH, Combs TP, Du X, Brownlee M and
Scherer PE: The adipocyte-secreted protein Acrp30 enhances hepatic
insulin action. Nat Med. 7:947–953. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Maeda N, Shimomura I, Kishida K, et al:
Diet-induced insulin resistance in mice lacking adiponectin/ACRP30.
Nat Med. 8:731–737. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Matsuzawa Y: Establishment of a concept of
visceral fat syndrome and discovery of adiponectin. Proc Jpn Acad
Ser B Phys Biol Sci. 86:131–141. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wulster-Radcliffe MC, Ajuwon KM, Wang J,
Christian JA and Spurlock ME: Adiponectin differentially regulates
cytokines in porcine macrophages. Biochem Biophys Res Commun.
316:924–929. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Matsuda M, Shimomura I, Sata M, et al:
Role of adiponectin in preventing vascular stenosis The missing
link of adipo-vascular axis. J Biol Chem. 277:37487–37491. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chen H, Montagnani M, Funahashi T,
Shimomura I and Quon MJ: Adiponectin stimulates production of
nitric oxide in vascular endothelial cells. J Biol Chem.
278:45021–45026. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Maeda N, Shimomura I, Kishida K, et al:
Dietinduced insulin resistance in mice lacking adiponectin/ACRP30.
Nat Med. 8:731–737. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ding M, Carrão AC, Wagner RJ, et al:
Vascular smooth muscle cell-derived adiponectin: a paracrine
regulator of contractile phenotype. J Mol Cell Cardiol. 52:474–484.
2012. View Article : Google Scholar :
|
|
13
|
Matsuda M, Shimomura I, Sata M, et al:
Role of adiponectin in preventing vascular stenosis. The missing
link of adipo-vascular axis. J Biol Chem. 277:37487–37491. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wang Y, Lam KS, Xu JY, Lu G, Xu LY, Cooper
GJ and Xu A: Adiponectin inhibits cell proliferation by interacting
with several growth factors in an oligomerization-dependent manner.
J Biol Chem. 280:18341–18347. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Motobayashi Y, Izawa-Ishizawa Y, Ishizawa
K, et al: Adiponectin inhibits insulin-like growth factor-1-induced
cell migration by the suppression of extracellular signal-regulated
kinase 1/2 activation, but not Akt in vascular smooth muscle cells.
Hypertens Res. 32:188–193. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Takaoka M, Nagata D, Kihara S, et al:
Periadventitial adipose tissue plays a critical role in vascular
remodeling. Circ Res. 105:906–911. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Miao CY and Li ZY: The role of
perivascular adipose tissue in vascular smooth muscle cell growth.
Br J Pharmacol. 165:643–658. 2012. View Article : Google Scholar :
|
|
18
|
Nepal S and Park PH: Regulatory role of
autophagy in globular adiponectin-induced apoptosis in cancer
cells. Biomol Ther (Seoul). 22:384–389. 2014. View Article : Google Scholar
|
|
19
|
Yamauchi T, Kamon J, Ito Y, et al: Cloning
of adiponectin receptors that mediate antidiabetic metabolic
effects. Nature. 423:762–769. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hug C, Wang J, Ahmad NS, Bogan JS, Tsao TS
and Lodish HF: T-cadherin is a receptor for hexameric and
high-molecular-weight forms of Acrp30/adiponectin. Proc Natl Acad
Sci USA. 101:10308–10313. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhou L, Deepa SS, Etzler JC, et al:
Adiponectin activates AMP-activated protein kinase in muscle cells
via APPL1/LKB1-dependent and phospholipase
C/Ca2+/Ca2+/calmodulin-dependent protein kinase kinase-dependent
pathways. J Biol Chem. 284:22426–22435. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Iwabu M, Yamauchi T, Okada-Iwabu M, et al:
Adiponectin and AdipoR1 regulate PGC-1alpha and mitochondria by Ca
(2+) and AMPK/SIRT1. Nature. 464:1313–1319. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Santel A and Fuller MT: Control of
mitochondrial morphology by a human mitofusin. J Cell Sci.
114:867–874. 2001.PubMed/NCBI
|
|
24
|
Chen KH, Guo X, Ma D, et al: Dysregulation
of HSG triggers vascular proliferative disorders. Nat Cell Biol.
6:872–883. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
25
|
Jin B, Fu G, Pan H, et al: Anti-tumour
efficacy of mitofusin-2 in urinary bladder carcinoma. Med Oncol.
28(Suppl 1): 373–380. 2011. View Article : Google Scholar
|
|
26
|
Wu L, Li Z, Zhang Y, et al:
Adenovirus-expressed human hyper-plasia suppressor gene induces
apoptosis in cancer cells. Mol Cancer Ther. 7:222–232. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang W, Zhu F, Wang S, et al: HSG provides
antitumor efficacy on hepatocellular carcinoma both in vitro and in
vivo. Oncol Rep. 24:183–188. 2010.PubMed/NCBI
|
|
28
|
Wang W, Lu J, Zhu F, et al: Pro-apoptotic
and anti-proliferative effects of mitofusin-2 via Bax signaling in
hepatocellular carcinoma cells. Med Oncol. 29:70–76. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chen KH, Dasgupta A, Ding J, Indig FE,
Ghosh P and Longo DL: Role of mitofusin 2 (Mfn2) in controlling
cellular proliferation. FASEB J. 28:382–394. 2014. View Article : Google Scholar :
|
|
30
|
Zhang GE, Jin HL, Lin XK, Chen C, Liu XS,
Zhang Q and Yu JR: Anti-tumor effects of mfn2 in gastric cancer.
Int J Mol Sci. 14:13005–13021. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ortmann J, Veit M, Zingg S, et al:
Estrogen receptor-α but not-β or GPER inhibits high glucose-induced
human VSMC proliferation: potential role of ROS and ERK. J Clin
Endocrinol Metab. 96:220–228. 2011. View Article : Google Scholar :
|
|
32
|
Sirois MG, Simons M and Edelman ER:
Antisense oligonucleotide inhibition of PDGFR-beta receptor subunit
expression directs suppression of intimal thickening. Circulation.
95:669–676. 1997. View Article : Google Scholar : PubMed/NCBI
|