Introduction
Renal carcinoma is the most common type of kidney
cancer in adults and is responsible for ~90–95% of the cases of
kidney cancer (1). As a result of
advances in chemotherapy, the long-term survival rates for patients
with renal carcinoma, exhibiting no detectable metastases at
diagnosis, have significantly improved. However, for patients that
present with metastasis or exhibit disease recurrence, the
long-term survival rates are low (2,3).
There is an ongoing requirement for novel therapeutic strategies
and a more thorough understanding of the mechanisms, which are
involved in the development of renal carcinoma and the metastatic
process, particularly to the lungs (4).
Ten-eleven translocation methylcytosine dioxygenase
1 (TET1) is a member of the TET family of enzymes (TET1/2/3),
which, in humans, is encoded by the TET1 gene that possesses a
zinc-binding CXXC domain containing eight conserved cysteine
residues, which bind zinc (5).
TET1 is identified as a 5-methylcytosine (5-mC) hydroxylase, which
catalyzes the conversion of the modified DNA base, 5-mC, into
5-hydroxy-methylcytosine (5-hmC) in vitro or in vivo
(6,7). Notably, substantial downregulation of
all three TET genes has been reported in various types of solid
tumor (8–12), and TET1 has been demonstrated to be
an essential tumor suppressor in prostate and breast cancer
(13,14). Until now, there have been no
reports regarding the expression levels of TET1 and its association
with clinical outcome in renal carcinoma.
Tumor tissue from patients with renal carcinoma has
been found to exhibit a significantly higher incidence of
hypermethylation in several genes, compared with corresponding
normal tissue (15). Methylation
of the promoter region of the P14ARF gene has been associated with
repression of its protein expression and a poor prognosis in
patients with renal carcinoma (16).
The present study aimed to demonstrate the
significance of TET1 in the prognosis of renal cancer, and to
investigate the roles of TET1 in the proliferative and migratory
abilities of renal carcinoma cells as well as its pro-apoptotic
effect.
Materials and methods
Tumor samples
The tumor samples were obtained from the tumor
tissues of patients diagnosed with renal carcinoma, which had been
preserved in the Cancer Tissue Bank between 2007 and 2013 at
Changzhou First People's Hospital (Jiangsu, China). Among the 54
selected tumor samples, 35 were from male patients and 19 were from
female patients. The average age of the patients was 36 years.
Informed consent for the experimental use of surgical samples was
obtained from all patients. All the tissue specimens were sampled
from the tumors at the time of surgery, snap frozen and stored at
−80°C until retrieval for experiments. The clinical specimens were
divided into two groups; a low tumor grade group and a high tumor
grade tumor group, graded according to the Price grading system.
Normal renal tissue was used as a control, which was also obtained
from the Cancer Tissue Bank in Changzhou First People's
Hospital.
Cell culture, plasmid construction and
transfection
The ACHN and 293T human renal carcinoma cell lines
were obtained from the Cell Bank of Type Culture Collection of
Chinese Academy of Sciences (Shanghai, China). The cultures were
grown in Dulbecco's modified Eagle's Medium (DMEM), supplemented
with 10% fetal calf serum (FCS; Life Technologies, Gaithersburg,
MD, USA), penicillin (80 U/ml) and streptomycin (100 U/ml), at 37°C
in a 5% CO2 atmosphere. All transfections were performed
using Lipofectamine 2000 (Invitrogen Life Technologies, Carlsbad,
CA, USA). The TET1 gene cloned in vector pCMV6-XL5 was purchased
from OriGene Technologies (Rockville, MD, USA) and the siRNA
against TET1 was synthesized by GenePharma Co., Ltd. (Shanghai,
China). The sequence of TET1 siRNA was 5′-GCTCGCGAGCTATAGAAGAAT-3′.
The ACHN cells were transfected with the indicated plasmids prior
to the subsequent western blot and flow cytometric analyses (BD
Accuri™ C6 flow cytometer; BD Biosciences, Ann Arbor, MI, USA).
Immunohistochemistry and western
blotting
The paraformaldehyde-fixed (10%) and
paraffin-embedded tissues were sectioned at 3 µm. The
paraffin sections were dried at 58°C for 2–4 h, then dewaxed with
xylene and hydrated with alcohol. Antigen retrieval solution (0.01
mol/l sodium citrate, pH 6.0) was added after the sections were
washed with phosphate-buffered saline (PBS). Subsequently, they
were boiled at a constant temperature of 95°C for 15 min. To
inhibit endogenous peroxidase activity, the sections were incubated
with H2O2 (3%) at room temperature for 10
min. Following washing with PBS, 10% fetal calf serum was added,
which was followed by a 30 min incubation at 37°C and washing with
PBS three times. The expression levels and subcellular
distributions of TET1 in the renal carcinoma tissue sections were
detected using primary antibody (rabbit monoclonal anti-TET1; cat.
no. ab157004; diluted 1:500), biotinylated secondary antibody (goat
polyclonal anti-rabbit IgG; cat. no. ab150077; diluted 1:1,000)
(Abcam Inc., Cambridge, MA, USA) and ABC reagent (Vector
Laboratories, Inc., Burlingame, CA, USA), with 3,3′diaminobenzidine
tetrahydrochloride (Sigma-Aldrich, St. Louis, MO, USA) as the
substrate. The expression levels were measured by counting the
number of cells with detectable immunoreactivity under an optical
microscope (Olympus IX71; Olympus, Tokyo, Japan). The total
proteins were extracted from the tissues or cells using lysis
buffer (Beyotime Institute of Biotechnology, Shanghai, China),
containing phenylmethylsulfonyl fluoride, and were subsequently
quantified using the Bradford method (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA). The proteins (20 µg) were separated on
10% SDS-PAGE gels (Sangon Biotech Co., Ltd., Shanghai, China),
transferred onto polyvinilydine fluoride or nitrocellulose
membranes (EMD Millipore, Billerica, MA, USA) and were visualized
using a BCIP/NBT kit (Gibco Life Technologies, Carlsbad, CA, USA).
The band densities were quantified by gray analysis using ImageJ
version 1.41o (National Institutes of Health, Bethesda, MD, USA).
The primary antibodies used were monoclonal anti-TET1 (dilution,
1:1,000) and anti-actin (dilution 1:100; cat. no. ab8227; Abcam
Inc.).
Viability and apoptosis detection
The rate of cell proliferation was assessed using
flow cytometry. Briefly, the cells (1×106) were stained
with propidium iodide (PI) and were monitored using flow cytometry.
The proliferation index was calculated using the following formula:
Proliferation index = (S + G2/M) / (S + G2/M + G0/G1). The levels
of apoptosis were monitored according to PI and annexin V binding,
according to the manufacturer's instructions (FITC Annexin V
Apoptosis Detection Kit I; BD Biosciences).
Cell proliferation assay
The cell counting kit-8 (CCK-8) assay was performed
with a commercial kit (Beyotime Institute of Biotechnology) to
analyze proliferation. The cells (1×105) were seeded in
96-well plates, and cultivated at 37°C for 1, 2, 3 and 4 days,
respectively. After 10 µl CCK-8 reagent was added into each
well, the plate was incubated at 37°C for 1 h, and the absorption
was read at 450 nm. For each cell line, five wells were used to
evaluate cell proliferation, which was performed in triplicate in
independent groups.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
The total RNA was extracted from the cells or
tissues using TRIzol reagent (Invitrogen Life Technologies) and
reverse transcribed using M-MLV Reverse Transcriptase (Promega,
Madison, WI, USA). RT-qPCR was performed using an ABI 7500 fast
sequence detection system (Applied Biosystems, Foster City, CA,
USA) with a SYBR green fluorescent label. The reaction mixtures
contained 10 µl 2X Power SYBR Green PCR Master mix (Applied
Biosystems), 3–5 pmol of each primer (Sangon Biotech Co., Ltd.) and
0.25 µl of the RT reaction product. The samples were run in
triplicate in optically clear 96-well plates (Corning Incorporated,
Corning, New York, NY, USA). The thermocycling parameters were as
follows: 95°C for 10 min, followed by 40 cycles of 95°C for 15 sec
and 60°C for 1 min, and a detection step at 72°C for 30 sec. The
actin transcript was used in each sample as an internal reference
standard to control for variations in mRNA content and quantity.
Each RNA sample was run in triplicate. The primer sequences used
were as follows: TET1, forward 5′-ACTCAACTCTCAGAAGCCCC-3′ and
reverse 5′-GGTGGCTCTCCCTGTAAGTT-3′; actin, forward
5′-CGCGGATCCATGGATGATGATATCGCCGCG-3′ and reverse
5′-CCGCTCGAGGAAGCATTTGCGGTGGACGAT-3′.
Migration assay
The cells were trypsinized in 3 ml 0.05% trypsin,
washed in PBS, and resuspended in FCS-free DMEM, supplemented with
0.1% bovine serum albumin. The migration assays were performed in a
24-well plate. The cell suspensions (5×105 cells/ml; 100
µl) were added into triplicate wells in the upper chambers
of Transwells. The lower chambers of the Transwells (Corning
Incorporated) were filled with 600 µl 15% FCS-containing
DMEM. Following incubation at 37°C for 15 h, the cells that had
migrated through the filter into the lower wells were fixed with
90% ethanol, stained with
3-(4,5-dimethylthiazol-2-)-2,5-diphenyltetrazolium bromide (5
mg/ml, 20 µl). The average numbers of cells, which had
migrated was determined by calculating the numbers in four
randomly-selected fields under a Olympus IX71 microscope
(magnification, ×400), with the number of cells expressed as a
percentage of the sum of the cells in the upper and lower
wells.
Statistical analysis
The association between the expression levels of
TET1 and the clinical characteristics were assessed using
χ2 tests. Univariate survival analysis was performed
using Kaplan-Meier's method and analyzed using a log-rank test to
assess survival differences between the groups. P<0.05 was
considered to indicate a statistically significant difference.
Analyses were performed using SPSS 10.0 statistical software for
Windows (SPSS, Inc., Chicago, IL, USA).
Results
Expression levels of TET1 are low in
renal carcinoma, particularly in high grade tumors
Renal carcinoma samples were collected from patients
diagnosed with low tumor grade and from patients with high tumor
grade, as defined by the Price grading system. The expression of
TET1 was determined in the two different groups of renal carcinoma
using immunohistochemistry. Immunoreactivity for TET1 in the high
tumor grade tissue samples was lower, compared with that in the low
tumor grade tissue samples. Notably, the two tumor groups exhibited
low immunoreactivity for TET1, compared with the control tissues
(Fig. 1A). In agreement with the
immunohistochemical results, the mRNA expression levels of TET1
were significantly lower in the high tumor grade tissues, compared
with the low tumor grade tissues (Fig.
1B; P<0.05). In addition, the results of the western
blotting revealed that the protein expression levels of TET1 were
lower in the high tumor grade tissues, compared with the low grade
tissues (Fig. 1C), which was
consistent with the results from the immunohistochemistry and
RT-PCR analyses. These analyses demonstrated that the expression
levels of TET1 were lower in the renal carcinoma samples than in
the control samples, particularly in the group containing tissues
from patients diagnosed with high tumor grades. This suggested that
the expression levels of TET1 may be involved in renal
carcinogenesis.
Low expression levels of TET1 are
associated with poor prognosis in renal carcinoma
The statistical analyses demonstrated that the
expression level of TET1 correlated negatively with the grade of
the tumor. Univariate survival analysis revealed significant
associations (P<0.05) between the patient prognosis, the tumor
size and the expression levels of TET1. However, no significant
associations were observed among genders or ages at diagnosis. The
survival probability was reduced in patients with renal carcinoma
tissues exhibiting lower expression levels of TET1 (Fig. 2).
ACHN renal carcinoma cell line exhibits
low levels of TET1 and high proliferation and migration
abilities
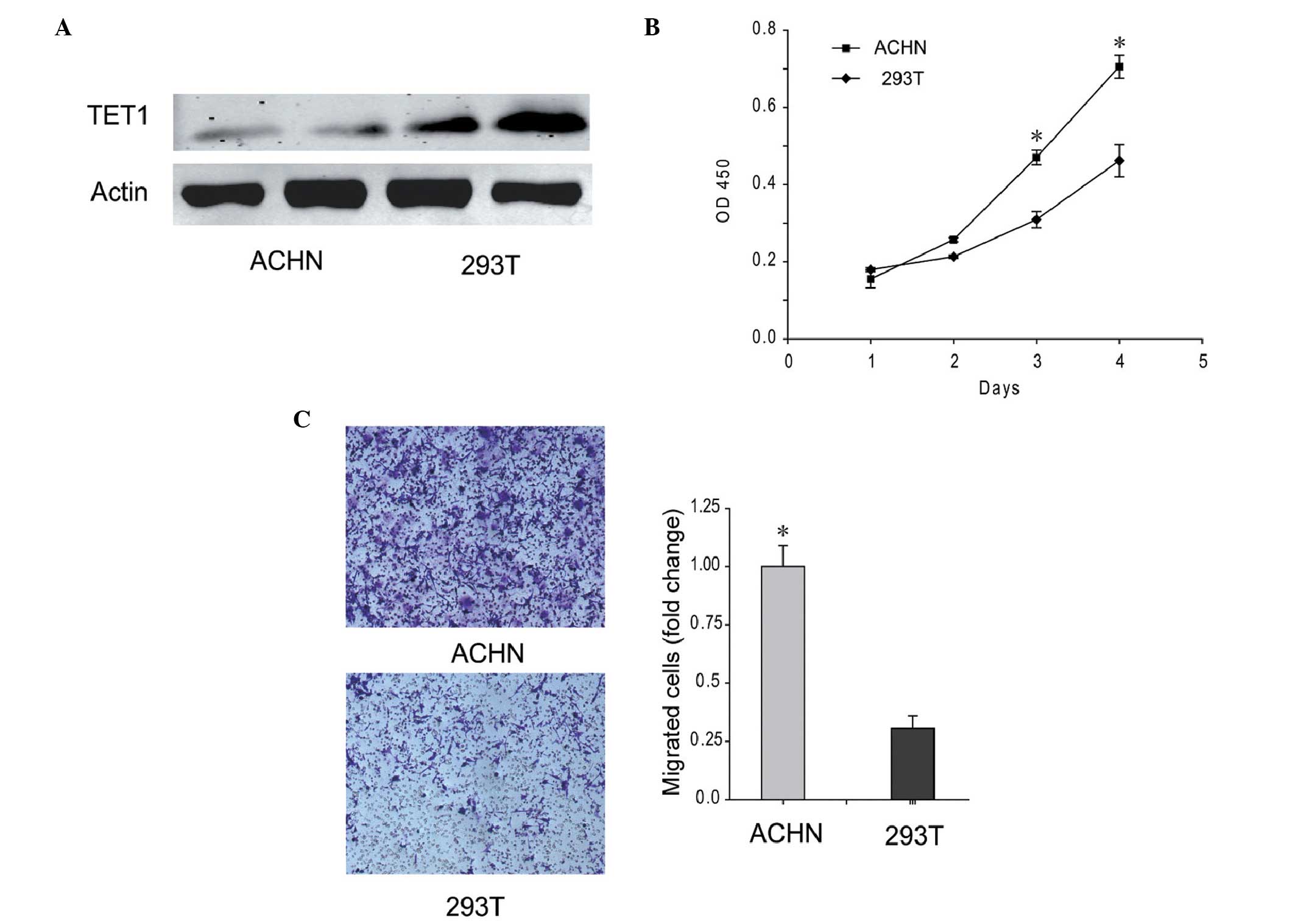
The expression of TET1 was low in the ACHN renal
carcinoma cell line, which exhibited the ability to metastasize to
the lungs. As shown in Fig. 3A,
the protein expression level of TET1 in the ACHN cells was
decreased, and was ~33.67% of the level detected in the 293T cells.
In addition, the proliferation and migration abilities of the ACHN
cells were increased, compared with 293T cells(Fig. 3B and C). These results indicated
that low expression levels of TET1 improved the proliferation and
migration abilities of the ACHN cells.
Overexpression of TET1 in ACHN cells
reduces proliferation and migration abilities, and increases
apoptosis
The present study also investigated the role of TET1
in renal carcinoma cells. Exogenous TET1 was expressed at high
levels in the ACHN renal carcinoma cell line and was 217.34% of the
level observed in the control cells (Fig. 4A). The proliferation index of the
ACHN cells overexpressing TET1 was markedly decreased (P<0.05)
compared with ACHN expressing background levels of TET1 (Fig. 4B). Additionally, the metastatic
ability was also reduced when TET1 was overexpressed in the ACHN
cells (Fig. 4C). The results of
the flow cytometric analysis demonstrated that 12.5% of the ACHN
cells containing exogenous TET1 were at the early stage of
apoptosis, whereas only 1.28% of ACHN cells expressing background
levels of TET1 were at the early stage (Fig. 4D). Taken together, these results
suggested that the overexpression of TET1 inhibited renal
carcinoma.
Discussion
Of the patients with renal carcinoma, the mortality
rate from pulmonary metastasis is >30% within 5 years after
diagnosis (3). However, the cause
of renal carcinoma remains to be fully elucidated. Downregulation
of the expression of TET has been observed in several types of
cancer, including human breast cancer, gastric cancer and prostate
cancer (10,12,17,18).
In the present study, the expression levels of TET1 were found to
be lower in renal carcinoma (Figs.
1 and 3A). Until now,
understanding of the function and mechanism of TET1 in renal cancer
has been limited.
TET1 is identified as a 5-mC hydroxylase, which
cata-lyzes the conversion of the modified DNA base, 5-mC, into
5-hmC, in vitro or in vivo (6,7).
Methylation at the C5 position of the cytosine bases is an
epigenetic modification of the mammalian genome, which is important
in transcriptional regulation (19). Hypermethylation of CpG islands
within the promoter and 5′ regions of genes is an important
epigenetic mechanism for suppressing gene expression (20). TET1 exerts a role in DNA
demethylation, by catalyzing the conversion of the modified DNA
base, 5-mC, into 5-hmC in an iron- and α-ketoglutarate-dependent
manner (7,21). The conversion of 5-mC into 5-hmC
has been suggested as the initial step of active DNA demethylation
in mammals (21).
Genetic alternations in the status of DNA
methylation, termed epigenetic alterations, are the most common
molecular alterations in human neoplasia (22). Previous evidence suggests that
epigenetic alterations, including methylation and histone
modification, of genes involved in cell cycle regulation and
apoptosis may contribute to the pathogenesis of renal carcinoma
(23). Patients with renal
carcinoma exhibit a significantly higher (P<0.05) incidence of
hypermethylation in several genes, compared with corresponding
normal tissues (15). The present
study demonstrated that lower expression levels of TET1 in tumor
samples from patients with high tumor grading were associated with
increased tumor size, metastasis and survival artes (Fig. 2). It appeared that TET1 may exert
its antitumor role by demethylating the hypermethylated CpG islands
within the promoter, and by activating gene expression,
particularly of those genes involved in cell cycle regulation and
apoptosis. The present study focused on the function of TET1 in
renal cancer, however, the epigenetic regulatory mechanism of TET1
in renal cancer require investigation in the future.
In the present study, the ACHN cell line was
selected for the functional investigation of TET1 in renal
carcinoma, which demonstrated low expression levels of TET1 and
high proliferation and migration abilities (Fig. 3). The overexpression of TET1
reduced the proliferation and migration of ACHN cells (Fig. 4B and C). In, addition, 12.5% of the
cells overexpressing TET1 were positively-labeled with annexin V
and negatively-labeled by PI, which suggested that cells
overexpressing TET1 undergo early apoptosis (Fig. 4D). In conclusion, TET1 may exert
its tumor inhibitory effects by downregulating the proliferation
and migration ability of renal carcinoma cells and inducing the
cells to undergo early apoptosis. These findings suggested that
TET1 exerted its effects through the demethylation of
hypermethylated CpG islands within the promoter and activating gene
expression, particularly of those involved in cell cycle regulation
and apoptosis, in an epigenetic manner.
Taken together, the findings of the present study
demonstrated that lower expression levels of TET1 were associated
with a high tumor grade, poor prognosis and other
clinicopathological features in renal carcinoma, and that
heterogenous TET1 exerts a tumor inhibitory role in renal tumor
cells.
References
|
1
|
Naito S, Yamamoto N, Takayama T, Muramoto
M, Shinohara N, Nishiyama K, Takahashi A, Maruyama R, Saika T,
Hoshi S, et al: Prognosis of Japanese metastatic renal cell
carcinoma patients in the cytokine era: A cooperative group report
of 1463 patients. Eur Urol. 57:317–326. 2010. View Article : Google Scholar
|
|
2
|
Wang LL: Biology of osteogenic sarcoma.
Cancer J. 11:294–305. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Shor AC, Keschman EA, Lee FY, Muro-Cacho
C, Letson GD, Trent JC, Pledger WJ and Jove R: Dasatinib inhibits
migration and invasion in diverse human sarcoma cell lines and
induces apoptosis in bone sarcoma cells dependent on SRC kinase for
survival. Cancer Res. 67:2800–2808. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Assouad J, Petkova B, Berna P, Dujon A,
Foucault C and Riquet M: Renal cell carcinoma lung metastases
surgery: Pathologic findings and prognostic factors. Ann Thorac
Surg. 84:1114–1120. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ono R, Taki T, Taketani T, Taniwaki M,
Kobayashi H and Hayashi Y: LCX, leukemia-associated protein with a
CXXC domain, is fused to MLL in acute myeloid leukemia with
trilineage dysplasia having t (10;11) (q22;q23). Cancer Res.
62:4075–4080. 2002.PubMed/NCBI
|
|
6
|
Ito S, D'Alessio AC, Taranova OV, Hong K,
Sowers LC and Zhang Y: Role of Tet proteins in 5mC to 5hmC
conversion, ES-cell self-renewal and inner cell mass specification.
Nature. 466:1129–1133. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tahiliani M, Koh KP, Shen Y, Pastor WA,
Bandukwala H, Brudno Y, Agarwal S, Iyer LM, Liu DR, Aravind L, et
al: Conversion of 5-methylcytosine to 5-hydroxymethylcytosine in
mammalian DNA by MLL partner TET1. Science. 324:930–935. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Guo JU, Su Y, Zhong C, Ming GL and Song H:
Hydroxylation of 5-methylcytosine by TET1 promotes active DNA
demethylation in the adult brain. Cell. 145:423–434. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Haffner MC, Chaux A, Meeker AK, Esopi DM,
Gerber J, Pellakuru LG, Toubaji A, Argani P, Iacobuzio-Donahue C,
Nelson WG, et al: Global 5-hydroxymethylcytosine content is
significantly reduced in tissue stem/progenitor cell compartments
and in human cancers. Oncotarget. 2:627–637. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yang H, Liu Y, Bai F, Zhang JY, Ma SH, Liu
J, Xu ZD, Zhu HG, Ling ZQ, Ye D, et al: Tumor development is
associated with decrease of TET gene expression and
5-methylcytosine hydroxylation. Oncogene. 32:663–669. 2013.
View Article : Google Scholar
|
|
11
|
Lian CG, Xu Y, Ceol C, Wu F, Larson A,
Dresser K, Xu W, Tan L, Hu Y, Zhan Q, et al: Loss of
5-hydroxymethylcytosine is an epigenetic hallmark of melanoma.
Cell. 150:1135–1146. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Fu HL, Ma Y, Lu LG, Hou P, Li BJ, Jin WL
and Cui DX: TET1 Exerts its tumor suppressor function by
interacting with p53-EZH2 pathway in gastric cancer. J Biomed
Nanotechnol. 10:1217–1230. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hsu CH, Peng KL, Kang ML, Chen YR, Yang
YC, Tsai CH, Chu CS, Jeng YM, Chen YT, Lin FM, et al: TET1
suppresses cancer invasion by activating the tissue inhibitors of
metalloproteinases. Cell Rep. 2:568–579. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sun M, Song CX, Huang H, Frankenberger CA,
Sankarasharma D, Gomes S, Chen P, Chen J, Chada KK, He C, et al:
HMGA2/TET1/HOXA9 signaling pathway regulates breast cancer growth
and metastasis. Proc Natl Acad Sci USA. 110:9920–9925. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hou P, Ji M, Yang B, Chen Z, Qiu J, Shi X
and Lu Z: Quantitative analysis of promoter hypermethylation in
multiple genes in osteosarcoma. Cancer. 106:1602–1609. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Oh JH, Kim HS, Kim HH, Kim WH and Lee SH:
Aberrant methylation of p14ARF gene correlates with poor survival
in osteosarcoma. Clin Orthop Relat Res. 442:216–222. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Frycz BA, Murawa D, Borejsza-Wysocki M,
Marciniak R, Murawa P, Drews M, Kołodziejczak A, Tomela K and
Jagodziński PP: Decreased expression of ten-eleven translocation 1
protein is associated with some clinicopathological features in
gastric cancer. Biomed Pharmacother. 68:209–212. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Sun M, Song CX, Huang H, Frankenberger CA,
Sankarasharma D, Gomes S, Chen P, Chen J, Chada KK, He C, et al:
HMGA2/TET1/HOXA9 signaling pathway regulates breast cancer growth
and metastasis. Proc Natl Acad Sci. 110:9920–9925. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ivanov M, Barragan I and Ingelman-Sundberg
M: Epigenetic mechanisms of importance for drug treatment. Trends
Pharmacol Sci. 35:384–396. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Jones PA and Laird PW: Cancer epigenetics
comes of age. Nat Genet. 21:163–167. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ito S, Shen L, Dai Q, Wu SC, Collins LB,
Swenberg JA, He C and Zhang Y: Tet proteins can convert
5-methylcytosine to 5-formylcytosine and 5-carboxylcytosine.
Science. 333:1300–1303. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hou P, Ji M, Yang B, Chen Z, Qiu J, Shi X
and Lu Z: Quantitative analysis of promoter hypermethylation in
multiple genes in osteosarcoma. Cancer. 106:1602–1609. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Rao-Bindal K and Kleinerman ES: Epigenetic
regulation of apoptosis and cell cycle in osteosarcoma. Sarcoma.
2011:6794572011. View Article : Google Scholar : PubMed/NCBI
|