Introduction
Acute liver failure (ALF) is defined as severe liver
damage induced by multiple factors and has a mortality rate of
80–90% (1). Current studies have
verified that hepatocyte apoptosis is important in the pathology of
ALF (1–5). The basic function of the
mitochondrion is energy metabolism, which provides all of the
energy necessary for life. A variety of studies have indicated
that, in addition to energy metabolism, modulation of cellular
apoptosis is the second main function of mitochondria (2,6,7). For
example, the release of cytochrome c and pro-apoptotic
proteins into the cytoplasm, calcium mobility and the generation of
reactive oxygen species (ROS) result in an alteration in
mitochondrial permeability and ATP depletion (3).
Hepatocytes are enriched with mitochondria that
comprise 13–20% of the liver volume. The liver is the chemical
center of the human body, consuming 20% of oxygen in the entire
body and is important in the metabolism of sugar, fat, protein,
water, salt and vitamins. The energy supply to the liver
predominantly originates from the oxidization of fatty acids. The
liver has a central position in lipid metabolism and is the
location of fatty acid β-oxidization (4). Therefore, investigating the
association between apoptosis and energy metabolism in hepatocyte
mitochondria during ALF has important practical value for
understanding the mechanisms underlying ALF, providing a basis for
the early diagnosis of ALF and developing a reasonable therapy for
ALF.
Metabolic pathways in the body consist of a series
of chemical reactions catalyzed by enzymes, of which the speed and
direction are determined by one or several key enzymes. The
modulation of energy metabolism, however, is primarily achieved by
modulating the activities of key enzymes (8). Citrate synthase (CS) is the key
enzyme and the first rate-limiting enzyme in the tricarboxylic acid
cycle (TCA). The CS of eukaryotes is coded by the nuclear genome,
synthesized in cytoplasmic ribosomes and exerts its function in the
mitochondrial matrix (9). The CS
is the rate-limiting enzyme of the TCA cycle and its activity can
modulate the cycle (10–12). Carnitine palmitoyltransferase-1
(CPT-1) is located in the outer membrane of mitochondria and
catalyzes long-chain fatty acyl-CoA and carnitine to synthesize
fatty acyl carnitine, which is the first rate-limiting reaction of
the oxidation procedure of fatty acids in mitochondria (13). Cytochrome c oxidase (COX) is
the final complex of electron transmission in the respiratory chain
and the key enzyme in oxidative phosphorylation in mitochondria
(14), and also plays an important
role in energy production (15).
These three enzymes are rate-limiting and are the key enzymes in
mitochondrial energy metabolism. Their activities can reflect the
mitochondrial energy metabolic function. Measuring the alterations
in the activities of these three enzymes can indirectly reflect
alterations in mitochondrial function. Previous studies have
indicated that decreases in the activities of CS, CPT-1 and COX
induced oxidative stress. This created excessive ROS (2), which increased the Ca2+
concentration in the cytoplasm (16,17),
initiated caspase reactions (5,18),
modulated the expression of the apoptosis-modulation proteins p53
and Bcl-2 (19,20) and produced cell apoptosis through
lipid peroxidation, amino acid oxidation/glycosylation of
intracellular biomembranes, including the mitochondrial membrane,
endoplasmic reticulum membrane and nuclear membrane, and base
oxidation of DNA (21). In
addition, oxidative stress produced an accumulation of excessive
acidic substances, activated lysosomal pathways leading to cellular
death, increased cellular sensitivity to tumor necrosis factor
(TNF)-α (22) and also initiated
apoptosis (23). Therefore,
determining the association between apoptosis and mitochondrial
energy metabolism warrants further investigation, particularly
during the course of ALF.
The present study aimed to observed the metabolic
characteristics of hepatocyte death during ALF and verify that the
change in energy metabolism is involved in ALF, particularly the
enhanced activities of CS, CPT-1 and COX during hepato-cyte
apoptosis.
Materials and methods
Animals and treatment
Male Sprague-Dawley (SD) rats weighing 200±20 g were
provided by the Experimental Animal Center of Beijing Youan
Hospital, Capital Medical University (Beijing, China). The animals
were housed at a room temperature of 22±2°C, with a light cycle
between 07:00 and 19:00 and free access to food and water. All
experimental procedures were approved by the Ethics Committee of
Animal Care and Usage of Capital Medical University. A total of 40
rats were randomly divided into five groups (n=8): Group A, control
group; Group B, 4 h after administration of D-galactosamine
(D-GalN) + lipopolysaccharide (LPS; ALF 4 h group); Group C, 8 h
after administration of D-GalN + LPS (ALF 8 h group); Group D, 12 h
after administration of D-GalN + LPS (ALF 12 h group); and Group E,
24 h after administration of D-GalN + LPS (ALF 24 h group). The
rats in Groups B–E were injected intraperitoneally (i.p.) with
D-GalN (800 mg/kg; Sigma-Aldrich, St. Louis, MO, USA) and LPS (100
µg/kg; Sigma-Aldrich). The rats in Group A (control) were
injected i.p. with 1 ml of 0.9% saline. At selected time points
following D-GalN/LPS treatment, rats were anesthetized and blood
was collected. The liver was isolated and used immediately to
prepare mRNA. The mRNA and liver tissues were stored at −75°C for
later analysis.
Assays of biochemical parameters
Serum alanine amino-transferase (ALT), aspartate
aminotransferase (AST) and total bilirubin (TBIL) were examined
using an automatic biochemical analyzer (Olympus AU5400; Olympus,
Tokyo, Japan); the prothrombin time (PT) and prothrombin activity
(PTA) were determined using an automatic coagulation analyzer
(SYSMEX CA-7000; Siemens Healthcare, Erlangen, Germany).
Histopathological analysis
Liver tissues were removed immediately and fixed in
10% neutral formalin solution for at least 24 h, then embedded in
paraffin wax and sectioned (4 µm thickness) for
histopathological evaluation. Liver sections were stained with
hematoxylin and eosin (H&E) using the Hematoxylin and Eosin
Staining kit (C0105; Beyotime Institute of Biotechnology, Haimen,
China), then the sections were analyzed by light microscopy (CKX41
Optical Microscope; Olympus).
Mitochondrial extraction
Mitochondrial extraction was immediately performed
on 400 mg of liver tissue from the middle portion of the right lobe
using a mitochondria extraction kit (Beijing Solarbio Science &
Technology Co., Ltd., Beijing, China) according to the
manufacturer's instructions. Half of the mitochondria were kept in
the 50 µl storage solution from the kit, from which 10
µl of solution was used to measure the activities of CS,
CPT-1 and COX using spectrophotometry (UV-2600 Ultraviolet-Visible
Spectrometer; Unico (Shanghai) Instrument Co., Ltd., Shanghai,
China) following protein quantification. The other half of the
mitochondria was fixed with glutaraldehyde for electron microscopic
observation of mitochondrial ultrastructure.
In situ apoptosis detection by TUNEL
Fresh liver tissue from the right lobe was fixed
with 4% paraformaldehyde for 24 h, and following
paraffin-embedding, was cut into 4 µm sections for TUNEL
(Boehringer Mannheim, Ingelheim am Rhein, Germany). Under the
mediation of terminal deoxynucleotidyl transferase, biotinylated
deoxyuridine triphosphate was labeled at the 3′-OH terminal formed
following the DNA break. Using specific binding of biotin and
avidin, peroxidase was linked to the DNA breakpoint. Substrate was
finally added to observe apoptotic cells with brown nuclei labeled
by 3,3′-diaminobenzidine staining under light microscopy. In total,
10 fields of view were randomly selected from sections of every
sample and the apoptotic cells were counted under a lens
(magnification, ×200) to calculate the apoptotic index (AI,
expressed as a percentage) in every 100 cells. Apoptotic cells fit
the following criteria: Single, no surrounding inflammatory cells,
membrane shrinkage and nuclei densely stained with brown particles
or debris.
Measurement of CS, CPT-1 and COX
The activities of CS, CPT-1 and COX were examined
using a CS assay kit (GMS50130.1; Genmed Scientifics, Inc.,
Shanghai, China), a CPT-1 assay kit (GMS50118.1.2; Genmed
Scientifics, Inc.) and a COX assay kit (GMS10014.2; Genmed
Scientifics, Inc.), respectively, according to the manufacturers'
instructions.
Western blot analysis
Protein was extracted from liver tissue with RIPA
buffer [20 mM Tris, pH 7.4, 137 mM NaCl, 0.5% sodium dodecyl
sulfate (SDS), 10% glycerol and 1% Triton X-100] with phosphatase
inhibitor 1 (100X), phosphatase inhibitor 2 (100X) and a protease
inhibitor cocktail (100X). Protein concentrations were determined
using a Bio-Rad detergent compatible Protein Assay kit (microtiter
plate; Bio-Rad Laboratories, Inc., Hercules, CA, USA). Proteins (40
mg sample) in SDS-loading buffer [50 mM Tris (pH 7.6), 10%
glycerol, 1% SDS] were subjected to SDS-12% polyacrylamide gel
electrophoresis and transferred onto a polyvinylidene difluoride
membrane (Bio-Rad Laboratories, Inc.). The membrane was blocked
with 5% dry milk and 0.1% Tween 20. Antibodies against COX1 (cat.
no. sc-23982; Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA),
CPT1 (cat. no. ab83862), CS (cat. no. ab129088) and β-actin (cat.
no. ab6276) (Abcam, San Francisco, CA, USA) were used for western
blot analysis. Membranes were probed with the primary antibodies
(1:1,000) in 10 ml blocking buffer overnight at 4°C. Following
washing, membranes were further probed with the appropriate
secondary antibody (1:2,000; cat. nos. cat. 115-035-003 and
111-035-003; Jackson ImmunoResearch Laboratories, Inc., West Grove,
PA, USA) in 10 ml of blocking buffer for 1 h at room temperature.
SuperSignal® West Pico Chemiluminescent Substrate
(Thermo Fisher Scientific, Rockford, IL, USA) was used for
chemiluminescence development.
Statistical analysis
The results are presented as the mean ± standard
deviation. Statistical comparisons among groups were performed by
analysis of variance followed by the t-test for independent samples
between two groups. The bilateral 95% confidence interval was
calculated using SPSS 13.0 software (SPSS, Inc., Chicago, IL, USA).
P<0.05 (two tailed) was considered to indicate a statistically
significant difference.
Results
Characteristic alterations of ALF induced
by D-GalN/LPS administration
The effects of D-GalN/LPS administration on the
liver of rats were initially assessed. Serum ALT, AST, TBIL, PT and
PTA and histological changes in the liver of rats were investigated
following D-GalN/LPS treatment. The results indicated that serum
ALT, AST, TBIL and PT were significantly elevated, whereas PTA was
significantly reduced, in a time-dependent manner following
D-GalN/LPS exposure (Table I). ALT
and AST began to increase at 4 h, reached a peak at 12 h and
decreased at 24 h after ALF. TBIL increased at 8 h and peaked at 24
h and PT began to increase while PTA decreased at 4 h after
ALF.
 | Table IAlterations in serum ALT, AST, TBIL,
PT and PTA in the liver of D-GalN/LPS-treated rats. |
Table I
Alterations in serum ALT, AST, TBIL,
PT and PTA in the liver of D-GalN/LPS-treated rats.
| Group | ALT (U/l) | AST (U/l) | TBIL
(µmol/l) | PT (sec) | PTA (%) |
|---|
| Control | 40.05±3.93 | 132.89±19.62 | 1.41±0.77 | 9.68±0.83 | 112.69±13.06 |
| ALF 4 h |
121.25±52.33a |
315.84±106.46a | 1.54±0.87 | 16.28±4.93a | 66.44±24.23a |
| ALF 8 h |
827.18±359.48a |
2489.06±1181.33a | 6.76±3.63a | 19.55±6.93a | 58.88±20.12a |
| ALF 12 h |
7778.60±4026.25a |
9461.90±5238.06a | 52.71±10.43a | 41.35±12.61a | 27.96±9.99a |
| ALF 24 h | 73.28±31.92b | 116.08±12.18 | 64.23±5.59a | 71.99±21.55a | 15.60±4.44a |
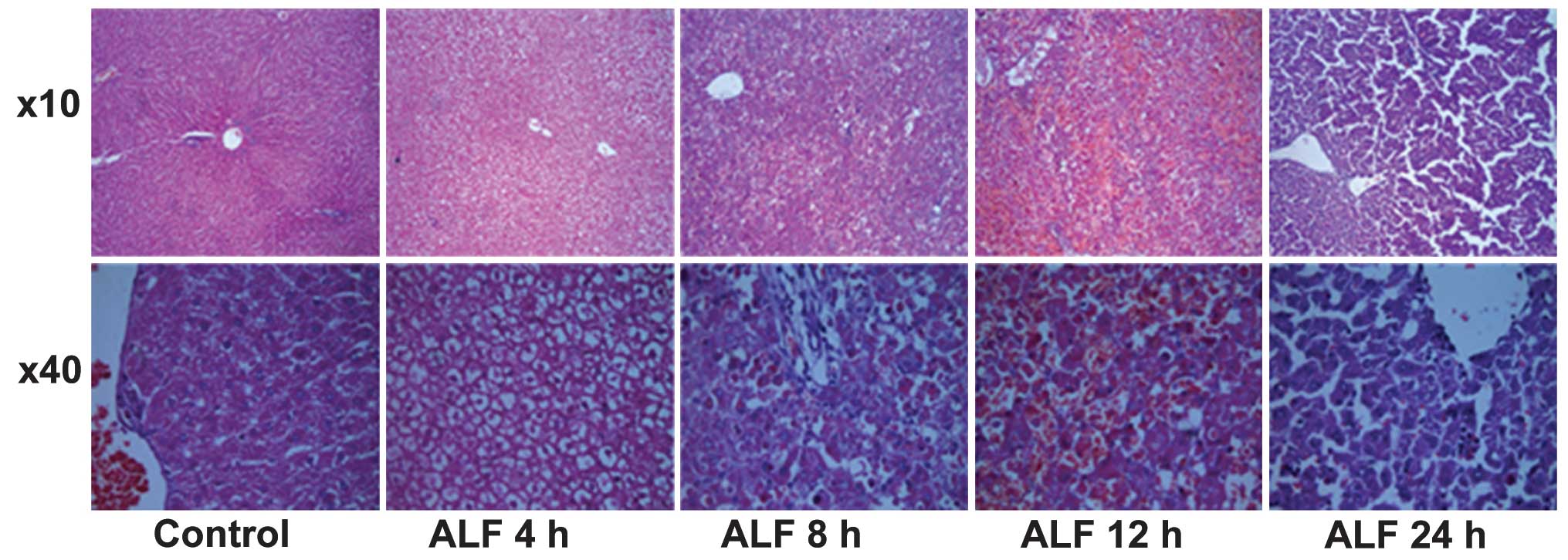
Histological examination revealed that the livers
from control group animals demonstrated complete hepatic lobules
without degeneration or necrosis by H&E staining. At ALF 4 h,
hepatic tissue underwent ballooning degeneration without clear
inflammatory invasion in the sinusoids. At ALF 8 h, there was
apparent hepatocyte apoptosis, partial hyperemia in the sinusoids,
piecemeal necrosis and inflammatory invasion. At ALF 12 h, a large
quantity of hepatocytes underwent piecemeal necrosis and there was
rupture of hepatic cords. At ALF 24 h, there was marked necrosis of
hepatocytes and severe hyperemia in the sinusoids, the boundary of
hepatocytes became obscure, the fiber mesh scaffold collapsed and
there was clear inflammatory cell invasion (Fig. 1).
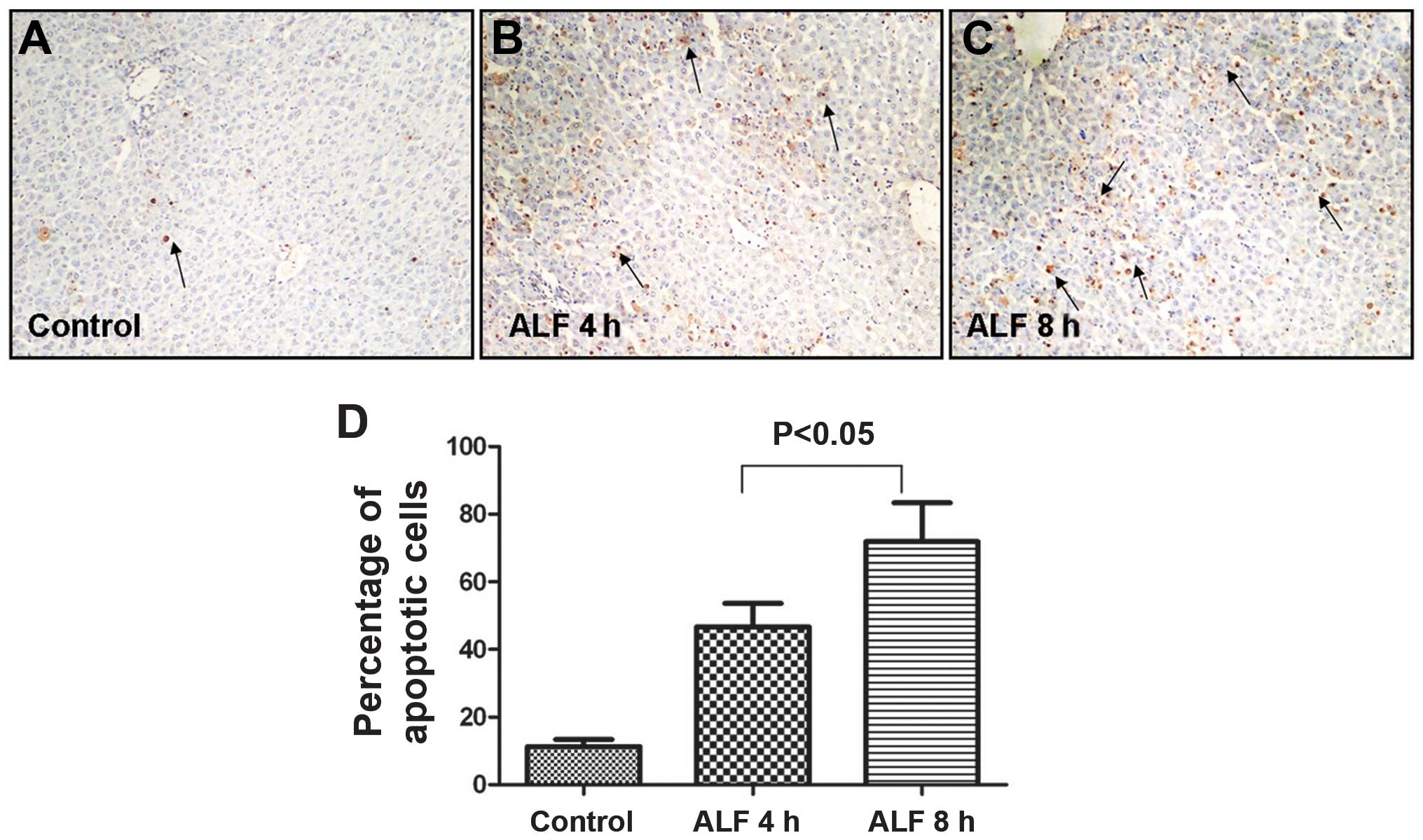
Hepatocyte apoptosis during the early
stages of ALF
TUNEL staining demonstrated that cellular injury
occurred predominantly in an apoptotic pattern during the early
stages of ALF. As shown in (Fig.
2A–C), apoptotic cells were predominantly located around the
central vein and were characterized by their shrunken, round shape,
small cell body, concentrated cytoplasm, increased density of
nuclear chromatin, which congregated near the nuclear membrane, and
their invaginated cellular membrane, which separated the cell into
membrane-encapsulated apoptotic bodies varying in size. The AI was
significantly different between the control group (10.60±2.61), ALF
4 h (48.90±10.76) and ALF 8 h groups (74.60±14.85; Fig. 2D).
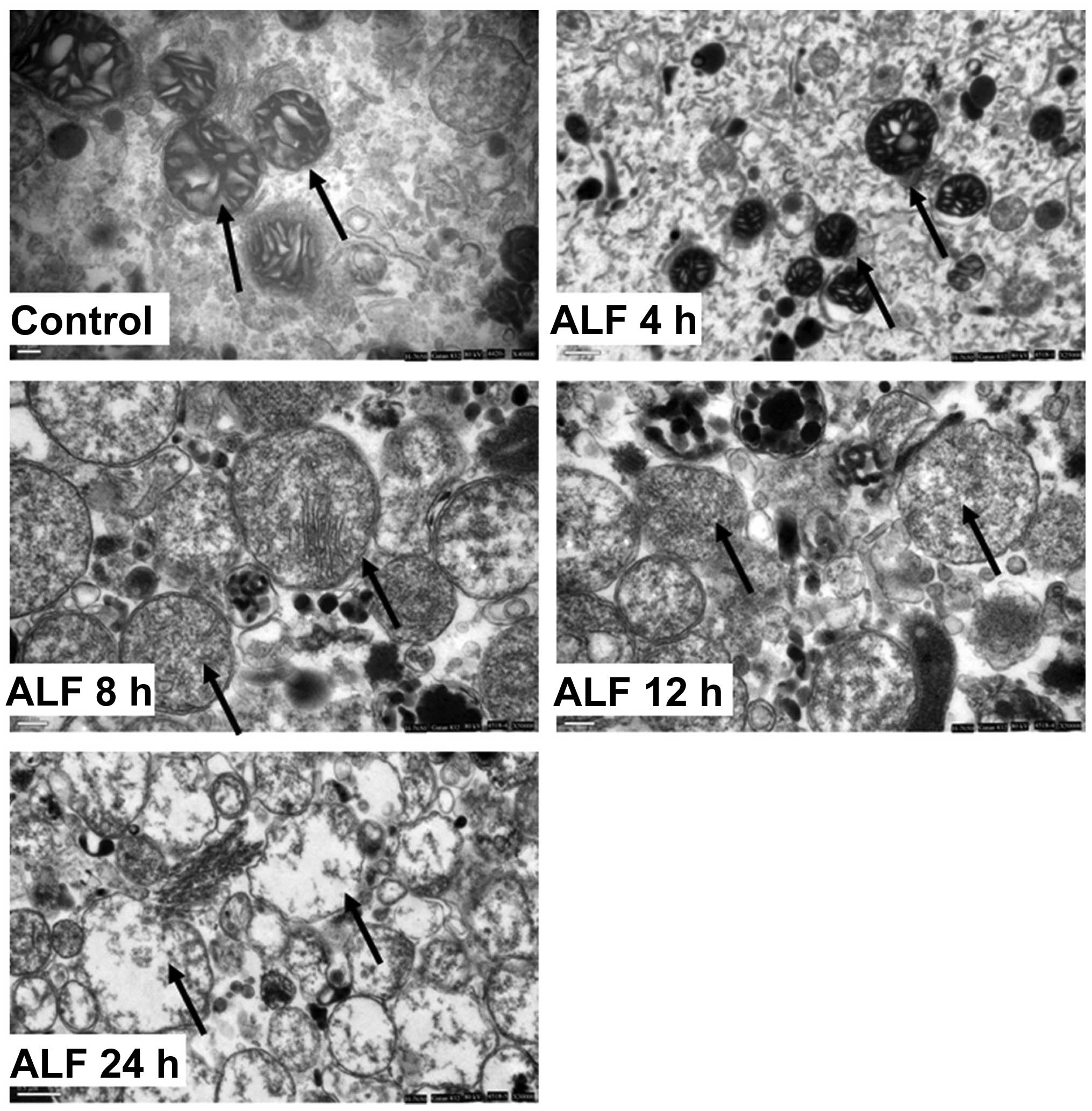
Morphological alterations in liver
mitochondria during ALF
The hepatocyte mitochondria from the control group
were round or oval with complete membrane structures and concentric
circles or longitudinally arranged ridges that were tidy and
compact. In the ALF 4 h group, the mitochondria were reduced in
number, fairly structured, fairly complete and had a complete
structure of ridges that had a fairly tidy arrangement but with
concentrated stroma. In the ALF 8 h group, the mitochondria were
swollen and clearly damaged but had somewhat complete outer
membranes and disorganized ridge structures. In the ALF 12 h group,
the mitochondria were swollen and their inner structures were not
easily visible. In the ALF 24 h group, the outer membrane was
disrupted and the mitochondria had collapsed (Fig. 3).
Activities of CS, CPT-1 and COX in liver
mitochondria and their expression in liver tissue
To assess the dynamic alterations of CS, CPT-1 and
COX during the course of ALF, the present study examined the
activities of CS, CPT-1 and COX in liver mitochondria and the
expression level of CS, CPT-1 and COX in liver tissue. As shown in
(Fig. 4A and B) the activities of
CS, CPT-1 and COX in liver mitochondria began to increase at 4 h
(Fig. 4C) peaked at 8 h, began to
decrease at 12 h and decreased markedly at 24 h. The present study
also determined the expression level of CS, CPT-1 and COX in liver
tissue during D-GalN/LPS-induced hepatic failure. As shown in
Fig. 4D, compared with the control
group, the expression levels of CS, CPT-1 and COX were upregulated
at 4 h and then decreased after 8 h in liver tissue.
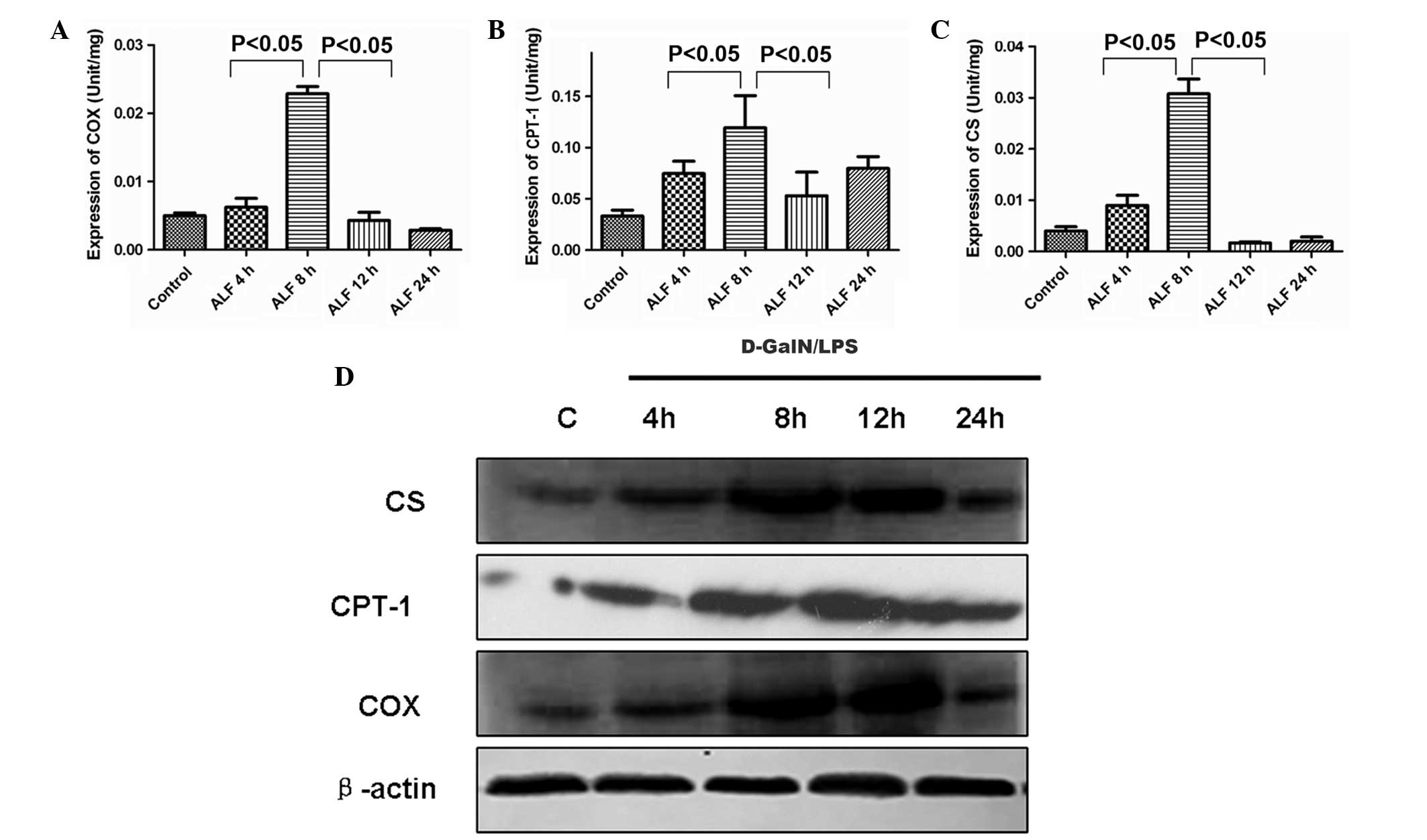 | Figure 4Activities of CS, CPT-1 and COX in
liver mitochondria and their protein expression level in liver
tissue. (A–C) The activity of CS, CPT-1 and COX in liver
mitochondria began to increase at 4 h, peaked at 8 h and began to
decrease at 12 h. (D) Western blotting demonstrated that the
protein expression levels of CS, CPT-1 and COX were upregulated at
4 h and then decreased after 12 h in liver tissue. ALF, acute liver
failure; CS, citrate synthase; CPT-1, carnitine
palmitoyltransferase-1; COX, cytochrome c oxidase; D-GalN,
D-galactosamine; LPS, lipopolysaccharide. |
Discussion
Mitochondria are critical during hepatocyte death
and the dysfunction of mitochondria is an obligatory process in
hepa-tocyte death. The increase in mitochondrial permeability and
dysfunction can induce hepatocyte apoptosis and necrosis.
Mitochondria undertake the metabolic function of producing energy
through the respiratory chain and the mitochondrial inner and outer
membranes can simultaneously quarantine pro-apoptotic proteins in
the periplasmic space. However, the metabolic function and
characteristics of mitochondria and their association with
hepatocyte apoptosis remain to be elucidated.
The combined administration of D-GalN and LPS is a
common method of establishing an ALF model in rodent animals
(24). LPS can directly stimulate
Kupffer cells to produce TNF-α and other inflammatory factors,
producing hepatocyte injury (25).
D-GalN is a specific hepatic toxicant that can deplete hepatic
uridine and induce hepatic injury (26). Combined administration of D-GalN
and LPS can enhance sensitivity to LPS and result in ALF (26). The present study successfully
duplicated this animal model.
ALF is characterized by broad hepatocyte death or
acute disruption of liver function and is the common outcome of all
severe liver diseases. Regardless of the pathogenesis, the final
manifestation is large-scale death of hepatocytes and other liver
cells. Currently, it is known that there are two types of
hepatocyte death: Apoptosis and necrosis. Apoptosis is a
physiological cellular suicide process that is induced by in
vivo and/or in vitro factors, is active and
energy-consuming, highly ordered, genetically controlled and
involves a series of enzymes (27). Necrosis, however, is an acute,
irreversible, and passive process that is characterized by
metabolic loss and destruction of cellular integrity (28). Previous studies indicated that
energy metabolism is critical in determining patterns of cell death
(29–32). Although there have been numerous
studies investigating ALF (13,16–18,28,33),
the mechanism underlying hepatocyte death during ALF remains to be
elucidated. Particularly, studies regarding the energy metabolic
characteristics and the association between key enzymes in energy
metabolism and hepatocyte apoptosis during ALF are controversial.
For example, previous studies testified that fatty acid metabolism
altered during ALF, however, this was predominantly determined by
measuring the plasma ketone level to reflect the oxidation level of
fatty acids in the liver (33,34);
and the plasma ketone level is affected by numerous factors,
including glucose supplementation and starvation.
The present study indicated that apoptosis and
necrosis occurred in hepatocytes during ALF, but with different
patterns at different stages. During the early stages of ALF,
hepatocyte apoptosis was predominant. Under light microscopy,
apoptosis was apparent at ALF 8 h, while necrosis was visible
during and after ALF 12 h. TUNEL staining also indicated that
apoptosis was most significant at ALF 8 h. The increase in serum
ALT and AST was due to the release of intracellular enzymes into
the blood following necrosis. The present study indicated that
serum ALT and AST peaked at ALF 12 h, while the activities of
mitochondrial COX, CS and CPT-1 peaked at ALF 8 h, earlier than the
ALT and AST peaks, indicating that the activities of COX, CS and
CPT-1 were enhanced during apoptosis and decreased during necrosis.
This implies that apoptosis is an active and energy-consuming
process (35,36). However, this result disagreed with
previous studies suggesting that decreased activities of COX, CS
and CPT-1 induced apoptosis, the production of ROS and the release
of cytochrome c and pro-apoptosis factors (37–40).
The reason for this discrepancy may be that apoptosis is an active
and energy-consuming process that requires a large quantity of ATP
supplementation at this stage, and this signal stimulates the
synthesis of enzymes to ensure the smooth progression of apoptosis,
as reflected in the enhancement of enzymatic activities. In
addition, cell apoptosis is a complex process and numerous
important factors are involved, including toxins, cytokines and
hormones. Therefore, hepatocyte apoptosis during the early stage of
ALF, may be mediated by other factors, while the energy metabolism
of mitochondria provides the necessary energy for apoptosis. By
contrast, enhanced enzymatic activities may also be a compensatory
mechanism.
The present study only observed the association
between the activities of CS, CPT-1 and COX and hepatocyte
apoptosis; it did not investigate the association between enzyme
activities and other factors, including cytokines, pro-apoptotic
proteins and inhibitors of apoptosis proteins. In addition, the
association between the enzymatic activity and signaling pathways
of hepatocyte apoptosis and necrosis is not clear. The present
study could not explain why the enzymatic activities in different
mitochondrial locations were enhanced, while the mitochondrial
structure altered. The presumption is that the enzymatic activity
is closely associated with the structure and permeability of the
mitochondrial outer membranes, but is less associated with the
structure of the mitochondrial inner membrane and ridge. Therefore,
the roles of key enzymes in different locations and alterations in
mitochondrial structures, in particular the permeability of the
mitochondrial outer membranes, should be investigated further to
determine the mechanism underlying changes in enzymatic activity
during ALF. Further studies investigating how hepatocyte energy
metabolism affects the transmission of death signals will provide a
basis for the early diagnosis and development of an improved
therapeutic strategy for ALF.
Abbreviations:
|
ALF
|
acute liver failure
|
|
CS
|
citrate synthase
|
|
CPT-1
|
carnitine palmitoyltransferase-1
|
|
COX
|
cytochrome c oxidase
|
|
ROS
|
reactive oxygen species
|
|
TCA
|
tricarboxylic acid cycle
|
|
D-GalN
|
D-galactosamine
|
|
LPS
|
lipopolysaccharide
|
|
TNF-α
|
tumor necrosis factor-α
|
|
ALT
|
alanine aminotransferase
|
|
AST
|
aspartate aminotransferase
|
|
TBIL
|
total bilirubin
|
|
PT
|
prothrombin time
|
|
PTA
|
prothrombin activity
|
|
TUNEL
|
terminal deoxynucleotidyl
transferase-mediated dUTP nick end labeling assay
|
Acknowledgments
This study is supported by the Chinese State key
Projects for Basic Research (grant no. 2007CB512801), the Chinese
National Research Grant of the Eleventh Five Year Plan for the Key
Projects in Infectious Diseases (grant no. 2008ZX100002-005-03) and
the Technology Projects of Fengtai Distinct, Beijing (grant no.
xm101210).
References
|
1
|
Liver Failure and Artificial Liver Group;
Chinese Society of Infectious Diseases; Chinese Medical
Association; Severe Liver Diseases and Artificial Liver Group:
Chinese Society of Hepatology, Chinese Medical Association:
Diagnostic and treatment guidelines for liver failure. Zhonghua Gan
Zang Bing Za Zhi. 14:643–646. 2006.In Chinese.
|
|
2
|
Orrenius S: Reactive oxygen species in
mitochondria-mediated cell death. Drug Metab Rev. 39:443–455. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kushnareva Y and Newmeyer DD:
Bioenergetics and cell death. Ann NY Acad Sci. 1201:50–57. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ryu SY, Peixoto PM, Teijido O, Dejean LM
and Kinnally KW: Role of mitochondrial ion channels in cell death.
Biofactors. 36:255–263. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ohta S: A multi-functional organelle
mitochondrion is involved in cell death, proliferation and disease.
Curr Med Chem. 10:2485–2494. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhao BC: Role of liver in lipid
metabolism. Biochemistry (Chin). Zhou AR: 18. 5th edition. People's
Medical Publishing House; Beijing: pp. 3652003
|
|
7
|
Zhao BC: Regulation of activities of
enzymes. Biochemistry (Chin). Zhou AR: 4. 5th edition. People's
Medical Publishing House; Beijing: pp. 63–64. 2003
|
|
8
|
Wiegand G, Remington S, Deisenhofer J and
Huber R: Crystal structure analysis and molecular model of a
complex of citrate synthase with oxaloacetate and
S-acetonyl-coenzyme. A J Mol Biol. 174:205–219. 1984. View Article : Google Scholar
|
|
9
|
Cheng TL, Liao CC, Tsai WH, Lin CC, Yeh
CW, Teng CF and Chang WT: Identification and characterization of
the mitochondrial targeting sequence and mechanism in human citrate
synthase. J Cell Biochem. 107:1002–1015. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Donald LJ and Duckworth HW: Molecular
cloning of the structural gene for Acinetobacter citrate synthase.
Biochem Biophys Res Commun. 141:797–803. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bloxham DP, Parmelee DC, Kumar S, Wade RD,
Ericsson LH, Neurath H, Walsh KA and Titani K: Primary structure of
porcine heart citrate synthase. Proc Natl Acad Sci USA.
78:5381–5385. 1981. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kerner J and Hoppel C: Fatty acid import
into mitochondria. Biochim Biophys Acta. 1486:1–17. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ostermeier C, Iwata S and Michel H:
Cytochrome c oxidase. Curr Opin Struct Biol. 6:460–466. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kadenbach B, Barth J, Akgün R, Freund R,
Linder D and Possekel S: Regulation of mitochondrial energy
generation in health and disease. Biochim Biophys Acta.
1271:103–109. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kadenbach B, Hüttemann M, Arnold S, Lee I
and Bender E: Mitochondrial energy metabolism is regulated via
nuclear-coded subunits of cytochrome c oxidase. Free Radic Biol
Med. 29:211–221. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Jezek P and Hlavatá L: Mitochondria in
homeostasis of reactive oxygen species in cell, tissues and
organism. Int J Biochem Cell Biol. 37:2478–2503. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Feldstein AE, Werneburg NW, Canbay A,
Guicciardi ME, Bronk SF, Rydzewski R, Burgart LJ and Gores GJ: Free
fatty acids promote hepatic lipotoxicity by stimulating TNF-alpha
expression via a lysosomal pathway. Hepatology. 40:185–194. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Malhi H, Bronk SF, Werneburg NW and Gores
GJ: Free fatty acids induce JNK-dependent hepatocyte lipoapoptosis.
J Biol Chem. 281:12093–12101. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Turrens JF: Mitochondrial formation of
reactive oxygen species. J Physiol. 552:335–344. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hsieh CL, Huang CN, Lin YC and Peng RY:
Molecular action mechanism against apoptosis by aqueous extract
from guava budding leaves elucidated with human umbilical vein
endothelial cell (HUVEC) model. J Agric Food Chem. 55:8523–8533.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Rana SV: Metals and apoptosis: Recent
developments. J Trace Elem Med Biol. 22:262–284. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Giordano A, Calvani M, Petillo O, Grippo
P, Tuccillo F, Melone MA, Bonelli P, Calarco A and Peluso G: tBid
induces alterations of mitochondrial fatty acid oxidation flux by
malonyl-CoA-independent inhibition of carnitine
palmitoyl-transferase-1. Cell Death Differ. 12:603–613. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Harada H and Grant S: Apoptosis
regulators. Rev Clin Exp Hematol. 7:117–138. 2003.
|
|
24
|
Deutschman CS, Haber BA, Andrejko K,
Cressman DE, Harrison R, Elenko E and Taub R: Increased expression
of cytokine-induced neutrophil chemoattractant in septic rat liver.
Am J Physiol. 271:R593–R600. 1996.PubMed/NCBI
|
|
25
|
Decker K and Keppler D: Galactosamine
hepatitis: Key role of the nucleotide deficiency period in the
pathogenesis of cell injury and cell death. Rev Physiol Biochem
Pharmacol. 71:77–106. 1974. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Qiu Z, Kwon AH, Tsuji K, Kamiyama Y,
Okumura T and Hirao Y: Fibronectin prevents
D-galactosamine/lipopolysac-charide-induced lethal hepatic failure
in mice. Shock. 25:80–87. 2006. View Article : Google Scholar
|
|
27
|
Canbay A, Friedman S and Gores GJ:
Apoptosis: The nexus of liver injury and fibrosis. Hepatology.
39:273–278. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Malhi H, Gores GJ and Lemasters JJ:
Apoptosis and necrosis in the liver: A tale of two deaths?
Hepatology. 43(2 Suppl 1): pp. S31–S44. 2006, View Article : Google Scholar
|
|
29
|
Vaquero J and Blei AT: Etiology and
management of fulminant hepatic failure. Curr Gastroenterol Rep.
5:39–47. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Van Herreweghe F, Festjens N, Declercq W
and Vandenabeele P: Tumor necrosis factor-mediated cell death: To
break or to burst, that's the question. Cell Mol Life Sci.
67:1567–1579. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Davis CW, Hawkins BJ, Ramasamy S, Irrinki
KM, Cameron BA, Islam K, Daswani VP, Doonan PJ, Manevich Y and
Madesh M: Nitration of the mitochondrial complex I subunit NDUFB8
elicits RIP1- and RIP3-mediated necrosis. Free Radic Biol Med.
48:306–317. 2010. View Article : Google Scholar
|
|
32
|
Zhang DW, Shao J, Lin J, Zhang N, Lu BJ,
Lin SC, Dong MQ and Han J: RIP3, an energy metabolism regulator
that switches TNF-induced cell death from apoptosis to necrosis.
Science. 325:332–336. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Clemmesen O: Splanchnic circulation and
metabolism in patients with acute liver failure. Dan Med Bull.
49:177–193. 2002.PubMed/NCBI
|
|
34
|
Saibara T, Maeda T, Onishi S and Yamamoto
Y: Plasma exchange and the arterial blood ketone body ratio in
patients with acute hepatic failure. J Hepatol. 20:617–622. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Lee SJ, Kwon CH and Kim YK: Alterations in
membrane transport function and cell viability induced by ATP
depletion in primary cultured rabbit renal proximal tubular cells.
Korean J Physiol Pharmacol. 13:15–22. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zischka H, Larochette N, Hoffmann F,
Hamöller D, Jägemann N, Lichtmannegger J, Jennen L, Müller-Höcker
J, Roggel F, Göttlicher M, et al: Electrophoretic analysis of the
mitochondrial outer membrane rupture induced by permeability
transition. Anal Chem. 80:5051–5058. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Marín-García J, Goldenthal MJ, Damle S, Pi
Y and Moe GW: Regional distribution of mitochondrial dysfunction
and apoptotic remodeling in pacing-induced heart failure. J Card
Fail. 15:700–708. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Chinta SJ, Rane A, Yadava N, Andersen JK,
Nicholls DG and Polster BM: Reactive oxygen species regulation by
AIF- and complex I-depleted brain mitochondria. Free Radic Biol
Med. 46:939–947. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Puig T, Relat J, Marrero PF, Haro D,
Brunet J and Colomer R: Green tea catechin inhibits fatty acid
synthase without stimulating carnitine palmitoyltransferase-1 or
inducing weight loss in experimental animals. Anticancer Res.
28:3671–3676. 2008.
|
|
40
|
Mazzarelli P, Pucci S, Bonanno E, Sesti F,
Calvani M and Spagnoli LG: Carnitine palmitoyltransferase I in
human carcinomas: A novel role in histone deacetylation? Cancer
Biol Ther. 6:1606–1613. 2007. View Article : Google Scholar
|