Introduction
Ovarian cancer is a highly lethal gynecological
malignancy (1,2). Cisplatin (DDP) is the basal
chemotherapeutic agent used to treat ovarian cancer, but due to an
increase in resistance to cisplatin (3), there is now an urgent need to explore
novel therapeutic interventions to treat ovarian cancer.
Chrysin (ChR), an active natural bioflavonoid found
in honey and extracts of numerous plants, has a number of
biological activities, including anti-oxidant (4), anti-inflammatory (5,6), and
anti-cancer activities (7–10). 8-bromo-7-methoxychrysin (BrMC) a
novel ChR derivative, has been reported to have anti-cancer
activities with more potent bioactivity than the lead compound
(11–14). It has been proposed that
BrMC-induced cell cycle arrest and apoptosis may be the mechanisms
of its anticancer effects (15,16).
However, the precise underlying molecular mechanisms by which BrMC
induces apoptosis in ovarian cancer cells are not fully
elucidated.
Fork-head box O3a (FOXO3a) is a fork-head
transcription factor of the FOXO subfamily characterized by a
'winged-helix' DNA-binding domain. FOXO3a is activated through
phosphorylation by several stress kinases such as Akt kinase, which
can upregulate multiple genes, of which B cell lymphoma 2 (Bcl-2)
interacting mediator of cell death (Bim) is the key promoter of
cell apoptosis. Akt has an oncogenic function, initially identified
as a proto-oncogene in the mouse leukemia virus Akt8 (17). Phosphorylated Akt isoforms are seen
at increased levels in the majority of human tumor types, and in
ovarian cancers (18–20). Bim, found in a wide variety of
tissues, is a member of the Bcl-2 homology domain 3 (BH3)-only
family of pro-apoptotic proteins (21,22).
Bim activates pro-apoptotic proteins Bcl-2 associated X protein
(Bax) and Bcl-2 homologous antagonist/killer (Bak), which in-turn
exert their apoptotic activities with other pro-apoptotic and
anti-apoptotic Bcl-2 family proteins (23). Akt is a key upstream regulator that
initiates FOXO3a dephosphorylation and nuclear translocation,
thereby enhancing the FOXO3a activity, leading to overexpression of
FOXO3a-responsive genes such as Bim (24,25).
Hence, FOXO3a links reduced Akt and increased Bim expression
levels, which results in the induction of apoptosis in cancer cells
(22,26,27).
In the present study, the focus was on understanding
the role of BrMC in promoting cycle arrest and apoptosis in ovarian
cancer cells. The role of the Akt/FOXO3a/Bim axis as a signaling
cascade mediating the anti-apoptotic activity of
cisplatin-sensitive and -resistant ovarian cancer was also
studied.
Materials and methods
Materials
BrMC was synthesized as described previously
(28). BrMC has a molecular weight
of 347 g/mol, appeared as yellow crystals and had a purity of
99.0%, which was determined as previously described (14). ChR was purchased from the Sigma
Chemical Co. (St Louis, MO, USA). BrMC and ChR were kept in
dimethyl sulfoxide (DMSO) and diluted to a final concentration of
0.1% in DMSO. Propidium iodide (PI), MTT, DMSO, a selevtice caspase
3 inhibitor z-DEVD-fmk and LY294002 were also obtained from Sigma.
Cell Apoptosis ELISA Detection kit (Roche Applied Sciences,
Penzberg, Germany) was purchased. Polyclonal rabbit anti-Bax (cat.
no. ab10813), mouse p53-upregulated modulator of apoptosis (PUMA;
cat. no. F02210), rabbit anti-NOXA (cat. no. BY-7074R), rabbit
anti-Bcl-2 (cat. no. HZ8392123) and rabbit anti-Bcl-extra large
(Bcl-XL; cat. no. ab32370), polyclonal rabbit anti-Bim (cat. no.
PC-033), rabbit anti-β-actin (cat. no. ab8229), rabbit
anti-caspase-3 (cat. no. bs-0081R), and rat anti-caspase-9 (cat.
no. C7729) were purchased from Santa Cruz Biotechnology, Inc.
(Santa Cruz, CA, USA). Caspase-3 specific inhibitor
Z-Asp-Glu-Val-Asp-CH2F (Z-DEVD-fmk) was obtained from
Calbiochem (La Jolla, CA, USA). Mouse monoclonal antibodies against
FOXO3a (cat. no. 04-1007), phospho-FOXO3a-Thr32 (cat. no.
JBC1381985), and antibodies against Akt (cat. no. 05-591) and
phospho-Akt (Ser473; cat. no. 17-457) were purchased from Millipore
(Bedford, MA, USA). Dulbecco's modified Eagle's medium (DMEM),
fetal bovine serum (FBS), and Lipofectamine™ 2000 were purchased
from Invitrogen Life Technologies (Carlsbad, CA, USA). The enhanced
chemoluminescence (ECL) western blot detection kit was purchased
from NENTM Life Science (Dordrecht, Netherlands).
Cells and cell culture
The human ovarian cancer cell line A2780 was
purchased from the China Center for Type Culture Collection (CCTCC;
Wuhan, China). The cells were seeded in DMEM supplemented with 10%
FBS (Hyclone), 100 U/ml penicillin and 100 µg/ml
streptomycin (Hyclone) and incubated at 37°C in a humidified
atmosphere of 5% CO2. Cisplatin-resistant cells were
obtained by unremitting treatments with increasing concentrations
of cisplatin (1, 5 and 10 µg/ml) (29). In brief, A2780 cells were exposed
to cisplatin (5 µg/ml) reproducibly at identical conditions
to those described above. Although cell death occured with
treatment, after four to six weeks of regular replacement of
culture medium (every two days) the drug-surviving cells aquired a
normal growth pattern. The concentration of cisplatin was then
increased to (7.5–10 µg/ml) and the process was repeated.
Following establishment, the chemo-resistant variants were treated
with cisplatin every month to maintain their high level of
chemo-resistance.
Histone/DNA ELISA
The cisplatin-induced rate of apoptosis in both
control and cisplatin-resistant cell lines were detected using an
ELISA detection kit (Roche Applied Sciences), according to the
manufacturer's instructions. Briefly, cells were cultured in
96-well plates at a density of 1×104 cells/well for 48 h
and the test agents were added to the culture medium containing 10%
FBS. After 24 h, the cytoplasm of cells in the control (untreated)
and treatment (cisplatin-treated) groups was transferred to
streptavidin-pre-coated 96-well plates, which had been previously
incubated with a biotinylated histone antibody and
peroxidase-tagged mouse anti-human DNA for 2 h at room temperature.
The absorbance was measured at 405 nm using an ELX-800 ELISA plate
reader (Bio-Tek, Winooski, VA, USA). In addition, the apoptosis
rate induced by BrMC treatment under identical conditions was
estimated using the same method.
MTT assay
Ovarian-cell viability was assessed by the MTT assay
(30). Cells were cultured in a
96-well plate at a density of 5,000 cells/well. Following
incubation for 24 h to allow for cell attachment, different
concentrations of BrMC (1.0, 2.5, 5.0 and 10.0 µmol/l) were
added to each well and cultured for 48 h. The medium was removed
and then incubated with 5.0 mg/ml MTT for 4 h. Following
centrifugation, the supernatant was removed. Finally, DMSO (100
µl) was added and absorbance at 570 nm wavelength (A570) was
measured by means of an enzyme-linked immune detector plate reader
(ELX-800 type; Bio-Tek, Shanghai, China). The relative cell
proliferation inhibition rate was caluculated as follows:
Inhibition rate = (1-average A570 of the experimental group/average
A570 of the control group)×100%. The IC50 (defined as
the drug concentration at which 50% cell viability was inhibited)
was assessed from the dose-response curves using GraphPad Prism
software (Version 4, GraphPad Software; La Jolla, CA, USA).
Cell cycle analysis by flow
cytometry
A2780 and A2780/DDP cells were seeded in six-well
plates at a density of 10,000 cells/well for 24 h, and then
different concentrations of BrMC (2.5, 5.0 and 10.0 µmol/l)
and ChR (50 µmol/l) were added and incubated for 48 h. The
cells were harvested and kept at 4°C for 12 h. Then PI (Sigma) was
added, and the apoptotic rate was analyzed by using a flow
cytometer (EPICSXL; Beckman Coulter, Inc., Brea, CA, USA), with
FC500 CXP software (Beckman Coulter, Inc.).
RNA interference
Akt and FOXO3a were purchased from Millipore; Bim
and control small interfering RNA (siRNA) were purchased from Santa
Cruz Biotechnology Inc. Human ovarian cancer A2780 and A2780/DDP
cells were transfected with Akt, FOXO3a, Bim and control siRNA
using Lipofectamine™ 2000, according to the manufacturer's
instructions (31). The cells were
then collected and processed for western blot analysis and
histone/DNA ELISA.
Western blot analysis
Total cell extracts for western blot analysis were
obtained, as described above for flow cytometry. Cell lysates
containing 50 µg protein were run on a 7.5–12.5% SDS-PAGE
and blotted onto polyvinylidene fluoride membranes (Millipore).
Western blot analysis was carried out as previously described
(15) and Anti-Bim, anti-Akt,
anti-FOXO3a, anti-phospho-FOXO3a-Thr32 and anti-caspase-3 were used
as primary antibodies. The blots were stripped at 37°C for 2 h.
After being washed with Tris-buffered saline and Tween 20 for 30
min, the corresponding secondary antibody was added and incubated
at room temperature for 1 h. The bound antibody was visualized
using chemiluminescent substrate (ECL; GE Healthcare, Arlington
Heights, IL, USA) and re-probed with an anti-actin antibody to
normalize for differences in protein loading. Changes in the levels
of the desired proteins were estimated using GelPro
Analyzer® image analysis (Media Cybernetics, Inc.,
Bethesda, MD, USA) of the immunoreactive bands and corrected for
β-actin loading control. Immunoblotting was performed a minimum of
two times for each protein using independently prepared lysates to
ensure reproducibility of the results.
Statistical analysis
The data were statistically analyzed using the SPSS
15.0 software package (SPSS Inc., Chicago, IL, USA) and presented
as the mean ± standard deviation. The means of multiple groups were
compared using one-way analysis of variance following the equal
check of variance and comparisons between the means were performed
using the least significant difference method. Statistical
comparison was also performed using a two-tailed t-test when
appropriate. P<0.05 was considered to indicate a statistically
significant difference between values.
Results
BrMC induces apoptosis in
cisplatin-sensitive and -resistant ovarian cancer cells
The growth inhibitory effect of cisplatin was
determined in various cisplatin-sensitive (A2780) and -resistant
(A2780/DDP) human ovarian cell lines. Cell viability was detected
by the MTT assay after 48 h of treatment with cisplatin. As shown
in Fig. 1A, cisplatin caused a
dose-dependent reduction of cell viability in A2780 cells, while
A2780/DDP cells exhibited resistance to cisplatin. To examine
cisplatin-induced apoptosis, the cells were treated with cisplatin
and apoptosis was confirmed by a histone/DNA fragmentation ELISA
assay at various concentrations. Cisplatin effectively induced
apoptosis in cisplatin-sensitive ovarian cancer cells, but not in
cisplatin-resistant cells (Fig.
1B). To investigate the effects of BrMC on apoptosis in both
ovarian cancer cell lines, the ovarian cancer cells were treated
with BrMC and the cell apoptosis process was analyzed by a DNA
fragmentation ELISA assay. BrMC induced cell apoptosis in both
ovarian cancer cell lines, regardless of their differences in
chemosensitivity (Fig. 1C). Flow
cytometric (FCM) analysis with PI staining further revealed that
there was no difference in BrMC-induced apoptosis between
cisplatin-sensitive and -resistant cells (Fig. 1D). To determine whether
mitochondria had an important role in BrMC-induced cell apoptosis,
the release of cytochrome c was detected by cell fractionation
analysis. The results revealed that BrMC induced the release of
cytochrome c in a time-dependent manner in both chemo-sensitive and
-resistant cells (Fig. 1E and F),
suggesting that BrMC initiated apoptotic cell death through
mitochondrial dysfunction.
 | Figure 1BrMC induces apoptosis in
cisplatin-sensitive and -resistant human ovarian cells. (A and B)
Effect of cisplatin on apoptosis in ovarian cancer cell lines in a
dose-dependent manner. Cells were treated with the indicated
concentrations of cisplatin for 48 h. ELISA was used to determine
histone/DNA fragmentation. Data are presented as the mean ± SD
(n=4). *P<0.05, **P<0.01 vs. 0.1% DMSO;
#P<0.05, ##P<0.01 vs. treatment with
2.5 µmol/l BrMC. (C and D) Effect of BrMC on apoptotic death
in cisplatin-sensitive and -resistant cells. Apoptosis was detected
using propidium iodide and flow cytometric analysis. Cells were
cultured in the presence or absence of BrMC (2.5 µM) for
different time periods, and apoptosis was measured. All data are
depicted graphically as the mean ± standard error of the mean for
at least three independent experiments. *P<0.05,
**P<0.01. (E and F) Analyses of cyt c release.
Following BrMC treatment for different periods of time, cells were
subjected to sub-cellular fractionation. The specific antibodies to
cyt c, β-actin and Cox IV were used. Densitometric analysis of the
western blots was performed and the amount of cyt c was compared
with that of the loading control. Data are representative of at
least three independent experiments. BrMC,
8-bromo-7-methoxychrysin; SD, standard deviation; DMSO, dimethyl
sulfoxide; DDP, cisplatin; cyt c, cytochrome c; Cox IV, cytochrome
c oxidase IV; Cyto, cytosol; Mito, mitochondria. |
Bim is required for BrMC-induced
apoptosis in chemo-sensitive and -resistant ovarian cancer
cells
Mitochondrial dysfunction has an important role in
apoptosis in ovarian cells (32).
Numerous studies have reported that it regulated gene expression of
Bcl-2-family proteins involved in chrysin-induced apoptosis
(32) and that the BH3-only
proteins were necessary for chrysin-induced apoptosis in cancer
cells (33). However, it remains
elusive whether BH3 proteins have the same function in ovarian
cancer cells following BrMC treatment. Therefore, the present study
investigated the expression of Bcl-2-family proteins in
cisplatin-sensitive and -resistant cells following BrMC treatment.
The levels of pro-apoptotic proteins, including Bax, PUMA and NOXA,
showed a slight change in A2780 cells at various time-points
following BrMC treatment (Fig.
2A). No major changes were recognized in anti-apoptotic Bcl-2
and Bcl-XL proteins. However, the expression levels of Bim showed a
marked increase following BrMC treatment, providing evidence that
Bim was involved in apoptotic cell death in ovarian cancer cells
treated with BrMC. BrMC induced Bim was expressed in the same
manner in A2780/DDP cells (Fig.
2B). These results implied that Bim had an important role in
BrMC-induced apoptosis in ovarian cancer cells.
 | Figure 2BrMC-induced ovarian cancer cell
apoptosis involved in Bim expression. (A and B) The expression
levels of Bcl-2 family proteins in chemosensitive and -resistant
ovarian cancer cells were examined by the time-dependent analysis
and western blot analysis. A2780 and A2780/DDP cells were treated
at the indicated concentrations with BrMC (5 and 10 µM) for
24 h. β-Actin was used as a control. (C) Under the same conditions
as A, cells were transiently transfected with a control
non-specific siRNA or Bim-specific siRNA for 48 h, and treated with
or without BrMC for 24 h. Cell lysates were obtained and assayed
for Bim, caspase-9 and cleaved caspase-3 by western blot. (D)
Effects of Bim on the apoptotic rate in A2780 and A2780/DDP cells.
All data were depicted graphically as the mean ± standard error of
the mean for at least three independent experiments.
*P<0.05, **P<0.01, vs. treatment with
siRNA alone; #P<0.05, vs. treatment with BrMC (5
µmol/l) and Bim siRNA. BrMC, 8-bromo-7-methoxychrysin;
Bcl-2, B-cell lymphoma 2; DDP, cisplatin; siRNA, small interfering
RNA; PUMA, p53-upregulated modulator of apoptosis; Bax, Bcl-2
associated X protein; Bim, Bcl2-interacting mediator of cell death;
Bcl-XL, B cell lymphoma extra large; Casp, caspase; Con, control
non-specific siRNA; BIM, bim-specific siRNA. |
To confirm the effect of Bim on apoptosis in ovarian
cancer cells, gene silencing experiments were performed using a
small interfering RNA (siRNA) specific for Bim that targeted all
known isoforms of its transcript. Similar to the results in
Fig. 2A, the expression levels of
Bim were high in the control vector-transfected A2780 and A2780/DDP
cells following BrMC treatment, while Bim expression levels in
cells transfected with Bim siRNA were relatively low (Fig. 2C). Down-regulation of Bim inhibited
both caspase-9 and caspase-3 activity induced by BrMC. Similarly,
BrMC induced an increase in the percentage of cells in sub-G1 phase
in the control vector-transfected A2780 and A2780/DDP cells, but
not in Bim siRNA-transfected cells (Fig. 2D). These results provided further
evidence that induction of Bim expression is necessary for
BrMC-induced apoptosis in ovarian cancer cells.
Akt dephosphorylation is involved in cell
apoptotic death in chemo-sensitive and -resistant ovarian cancer
cells
Recent studies have indicated that Akt modulates Bim
activation either directly or indirectly (34,35).
Moreover, chrysin induced apoptosis in leukemia cells by decreasing
Akt activity and expression in cells (36). Based on these observations, it was
speculated that Akt may be involved in BrMC-induced apoptosis by
regulating Bim expression. Analysis was performed to determine
whether Akt was involved in BrMC-induced apoptosis in ovarian
cancer cells. As shown in Fig. 3A,
BrMC induced the downregulation of Akt phosphorylation at Ser 473
in cisplatin-sensitive A2780 cells in a time-dependent manner.
However, expression levels of total Akt protein exhibited no
change. Similarly, BrMC induced the reduction of p-Akt in A2780/DDP
cells (Fig. 3B), regardless of
differences in chemo-sensitivity. To validate the role of Akt in
BrMC-induced apoptosis, LY294002, a phosphoinositide 3-kinase
inhibitor, was used to inhibit the phosphorylation of Akt by
upstream signaling molecules and the pro-apoptotic effect of BrMC
in ovarian cancer cells was examined. BrMC alone downregulated
p-Akt and induced apoptosis, while LY294002 alone was not
sufficient to induce apoptosis, though it partially decreased
p-Akt. Further experiments proved that the combination of LY294002
and BrMC completely eliminated p-Akt and enhanced the induction of
apoptosis in A2780 and A2780/DDP cells (Fig. 3C and D). This indicated that the
effect of the combination of LY294002 and BrMC on p-Akt and
apoptosis was more significant than that of LY294002 alone, and
that the Akt signaling pathway was involved in BrMC-induced
apoptosis in ovarian cancer cells.
FOXO3a mediates upregulation of Bim
during BrMC-induced apoptosis in chemosensitive and -resistant
ovarian cancer cells
Akt mediates Bim activation via two primary
pathways: (1) Akt phosphorylates
Bim directly and inhibits Bim activation (34,35);
(2) Akt phosphorylates FOXO3A,
leading to its cytoplasmic retention by 14-3-3 proteins (11,37).
This prevents FOXO3a translocation to the nucleus to induce Bim
transcription, demonstrating that Akt indirectly regulates Bim
activation. Therefore, the effect of BrMC on FOXO3a phosphorylation
was examined. Fig. 4A and B,
demonstrate the dose-dependent inhibition of FOXO3a phosphorylation
by BrMC. As phosphorylation suppresses the transcriptional activity
of FOXO3a, the expression of Bim was subsequently examined. As
shown in Fig. 4A and B, in
parallel with reduced FOXO phosphorylation, Bim was upregulated in
response to BrMC treatment in A2780 and A2780/DDP cells.
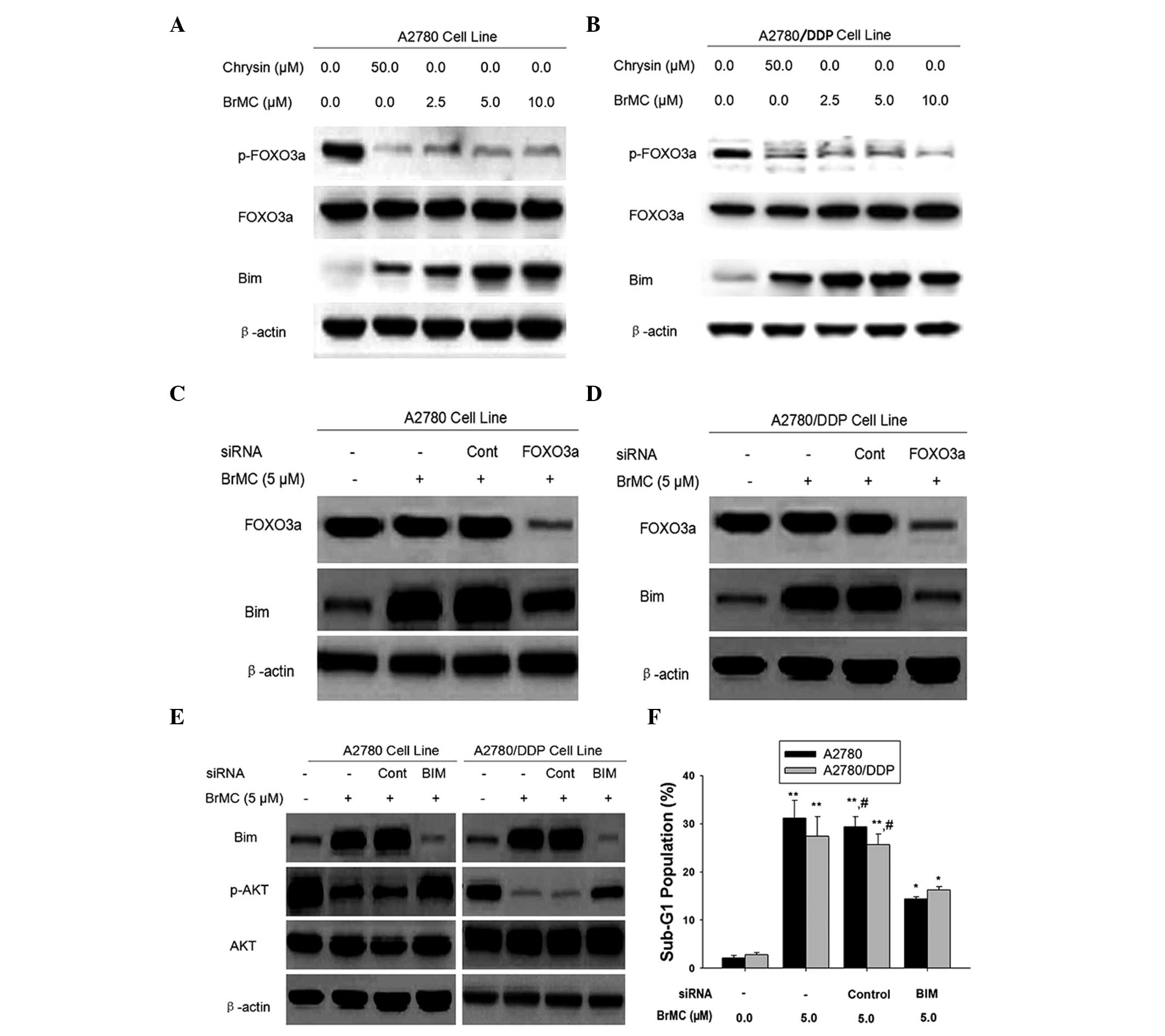 | Figure 4FOXO3a protein mediates Bim
activation in ovarian cancer cells. Western blot analysis of (A)
A2780 and (B) A2780/DDP cells treated with BRMC for 24 h. The
expression of p-FOXO3a, t-FOXO3a and Bim was analyzed and β-actin
was used as a loading control. (C and D) Cells were transiently
transfected with a control non-specific siRNA or FOXO3a-specific
siRNA for 48 h and treated with or without BrMC for 24 h. Cell
lysates were obtained and assayed for FOXO3a and Bim by western
blot. (E) Effect of Bim-siRNA on Akt phosphorylation. Cells were
transfected with a control non-specific siRNA or Bim-specific siRNA
for 48 h and were exposed to BrMC for 24 h. All data are
representative of three independent experiments. (F) Effects of
Bim-siRNA on the apoptotic rate of the A2780 and A2780/DDP cells.
All data are depicted graphically as the mean ± standard error of
the mean for at least three independent experiments.
*P<0.05, **P<0.01, vs. apoptotic rate
of the A2780 and A2780/DDP cells treated with siRNA alone;
#P<0.05, vs. treatment with BrMC (5 µm) and
Bim siRNA.. p/t-FOXO3a, phosphorylated/total forkhead box O3a;
BrMC, 8-bromo-7-methoxychrysin; Bim, Bcl2-interacting mediator of
cell death; siRNA, small interfering RNA; DDP, cisplatin; Cont,
control; BIM, bim-specific siRNA; z-DEVD-fmk, Caspase-3 specific
inhibitor Z-Asp-Glu-Val-Asp-CH2F. |
To examine the effect of FOXO3a activation on the
expression of Bim, A2780 and A2780/DDP cells were transfected with
FOXO3a siRNA followed by treatment with BrMC for 24 h and Bim
expression was then measured by western blot analysis (Fig. 4C and D). FOXO3a siRNA attenuated
the induction of Bim expression by BrMC treatment (Fig. 4C and D). These data suggested that
BrMC may regulate the expression of the FOXO3a transcriptional
target, Bim. However, knockdown of Bim inhibited Akt
dephosphorylation and apoptosis in chemo-sensitive and -resistant
ovarian cancer cells (Fig. 4E and
F). These results indicated that BrMC-induced Bim expression
prevented the phosphorylation of Akt, indicating that Bim regulated
Akt activation during BrMC stimulation in ovarian cancer cells.
Bim regulates Akt dephosphorylation by
caspase-3 activity during BrMC-induced apoptosis in chemosensitive
and -resistant ovarian cancer cells
It was reported that caspase-3 regulated the
phosphorylation of Akt through the cleavage of the regulatory A
subunit of (PP2A/A), increasing PP2A activity (38). In the caspase inhibitor assay,
cells were pretreated with caspase inhibitors (20 µM
ZVAD-CHO) for 1 h prior to the addition of the agents tested.
Results of the present study demonstrated that Bim was involved in
the regulation of Akt dephosphorylation (Fig. 4E). Therefore, it was proposed that
Bim may regulate Akt phosphorylation by caspase-3-mediated PP2A
activation in ovarian cells. To validate this hypothesis, it was
first determined whether PP2A mediated Akt phosphorylation.
Pretreatment of the cells with okadaic acid (OA), an inhibitor of
PP2A, resulted in reversal of BrMC-induced Akt dephosphorylation in
A2780 cells (Fig. 5A). At the same
time, OA also rescued Akt phosphorylation in A2780/DDP cells during
BrMC treatment (Fig. 5B).
 | Figure 5Caspase-3 regulates the
phosphorylation of Akt associated with PP2A. (A and B) A2780 and
A2780/DDP cells were pre-treated with indicated concentrations of
okadaic acid for 1 h and exposed to BrMC for 24 h, then detected
for Akt and p-Akt (Ser473) levels by western blot analysis. β-Actin
was used as a control. (C) To determine the Akt phosphorylation.
A2780/DDP cells were pre-treated with or without various doses of
caspase 3 inhibitor z-DEVD-fmk (20 or 80 µM) for 1 h and
exposed to BrMC (5 µM) for 24 h, then p-Akt (Ser473) levels
were detected using western blot analysis. (D) The apoptotic rate
of the A2780 and A2780/DDP cells following pre-treatment with
different doses of z-DEVD-fmk. All data are depicted graphically as
the mean ± standard error of the mean of at least three independent
experiments. *P<0.05, **P<0.01, vs.
apoptotic rate of the A2780 and A2780/DDP cells treated with BrMC
(5 µm) alone; #P<0.05, vs. treatment with BrMC
(5 µm) and z-DEVD-fmk (20 µm). (E and F) Detection of
the binding of Akt with PP2A. Cells were incubated with or without
BrMC for 48 h, 20 µM of z-DEVD-fmk was added 1 h prior to
treatment with the drug. The cells were lysed with lysis buffer for
immunoprecipitation with anti-Akt antibody followed by immunoblot
assay with anti-PP2A/C and anti-Akt antibodies. Data are
representative of at least three independent experiments. p-Akt,
phosphorylated Akt; CF, cleaved form of PP2A; PP2A, protein
phosphotase 2A; BrMC, 8-bromo-7-methoxychrysin; DDP, cisplatin;
z-DEVD-fmk, Caspase-3 specific inhibitor
Z-Asp-Glu-Val-Asp-CH2F. |
To determine whether caspase-3 mediates Akt
phosphory-lation, cells were pre-treated with z-DEVD-fmk, a
specific caspase-3 inhibitor. The results indicated that inhibition
of caspase-3 activation restored Akt phosphorylation in the
presence of BrMC (Fig. 5C and
D).
In addition, as shown in Fig. 5E and F, a low but detectable
quantity of the catalytic C subunit of PP2A (PP2A/C) was
co-precipitated with Akt in untreated cells, indicating the
occurrence of a physiological association between these two
proteins. Of note, the PP2A-Akt association was greatly improved
after 24 h treatment with BrMC, whereas this interaction was
significantly decreased by the inhibition of caspase-3 and PP2A
activation. This suggested that the dephos-phorylation of Akt was a
consequence of caspase-mediated PP2A activation and that the
caspase-3-PP2A-Akt pathway provided a positive feedback loop for
apoptosis. These results also indicated that Bim-caspase-Akt may
represent a novel pathway that regulates BrMC-induced apoptosis in
ovarian cancer cells.
Discussion
The present study found that BrMC had a stronger
effect than ChR on the inhibition of apoptosis in the colon cancer
cell line HT-29 and gastric cancer cell line SGC-7901.
Understanding the mechanism by which BrMC induces apop-tosis may
identify potential novel targets for cancer therapies. The present
study demonstrated that BrMC induced apoptotic cell death of
cisplatin-sensitive and -resistant ovarian cancer cells in a
dose-dependent manner and induced the release of cytochrome c in a
time-dependent manner. The apoptotic effect of BrMC was found to be
greater than that of ChR, regardless of their differences in
chemosensitivity. Furthermore, it was shown that BrMC induced
apoptosis of cisplatin-sensitive and -resistant ovarian cancer
cells accompanied with the upregulation of Bim. Conversely, the
downregulation of Bim inhibited caspase-9 and caspase-3 activities
induced by BrMC. It was also found that the downregulation of p-Akt
at Ser 473 in BrMC-treated cells occurred in a time-dependent
manner. In addition, it was indicated that BrMC eliminated p-Akt
and enhanced induction of apoptosis. Moreover, BrMC inhibited the
expression of p-FOXO3a in cells, which in parallel increased Bim
expression with increasing BrMC dose, therefore demonstrating that
the FOXO3a transcriptional target was upregulated. Furthermore, Bim
may regulate Akt phosphorylation by caspase-3-mediated PP2A
activation in ovarian cells (Fig.
5).
It was demonstrated that Bim had a very important
role in cisplatin-sensitive and -resistant ovarian cancer cells
during BrMC-induced apoptosis through detecting the expression
levels of Bcl-2-family proteins. It has been reported that
upregulation of Bim expression contributes to nitric oxide
synthatse releasing compound-induced apoptosis in MDA-MB-453 cells
(39). Therefore, results of the
present study confirmed that the upregulation of Bim prompted
apop-tosis induced by BrMC treatment. Furthermore, the present
study showed that downregulation of Bim by siRNA inhibited
BrMC-induced cell apoptosis.
Akt fuctioning as a stress kinase can upregulate
multiple genes that promote cell apoptosis (40). It has been reported by several
studies that Akt modulates Bim activation either directly or
indirectly, resulting in cell death (22,34,35).
The present study showed that pre-treatment with LY294002 to
inhibit the phosphorylation of Akt enhanced BrMC-induced apoptosis
in chemosensitive cells. These activities were accompanied by a
decrease in p-Akt, but not total Akt. Furthermore, the data
indicated that knockdown of Bim inhibited Akt dephosphorylation and
apoptosis in chemo-sensitive and -resistant ovarian cancer cells.
The FOXO3a transcription factor is a tumor suppressor that is
inactivated in the majority of human cancers (40). FOXO3a can directly up-regulate Bim
expression levels and induce apoptosis in cells. Akt-dependent
phosphorylation of FOXO3A leads to its cytoplasmic retention by
14-3-3 proteins and loss of target gene activation (11,37).
Consequently, FOXO3a cannot translocate into the nucleus and induce
Bim transcription, which therefore indicates indirect Akt
regulation of Bim activation. In the present study, it was
demonstrated that BrMC inhibited FOXO3a phosphorylation. Moreover,
knockdown of FOXO3a by transfection with siRNA blocked the
BrMC-induced downregulation of Bim expression and inhibited ovarian
cancer cell apoptosis. These findings indicated that FOXO3a was a
key regulator of BrMC-induced apoptosis in ovarian cancer
cells.
Conversely, observation of the mechanism of p-Akt
showed that Bim siRNA prevented caspase-3 cleavage. In addition, a
PP2A-linked pathway to determine Bim-dependent phosphorylation of
Akt during BrMC-induced ovarian cancer cell apoptosis was detected
as well as the factors that mediate Akt/FOXO3a expression during
cell apoptosis.
In conclusion, the present study indicated that BrMC
regulated the Akt/FOXO3a signal cascade pathway through mediating
the expression of its target genes, including Bim, caspase-3 and
PP2A, leading to apoptosis in ovarian cancer cells. A thorough
understanding is required to assess the anticancer activities of
BrMC in vivo. The mechanisms of BrMC-induced apoptosis may
lead to the identification and development of novel therapeutic
compounds for the treatment and prevention of ovarian cancer.
Acknowledgments
The authors gratefully acknowledge the assistance of
Dr Jianguo Cao for instructing the experiments and providing the
laboratory facilities. The authors would also like to thank Mrs
Kaiqun Ren for analyzing the data.
References
|
1
|
Lan C, Chenggang W, Yulan B, Xiaohui D,
Junhui Z and Xiao W: Aberrant expression of WWOX protein in
epithelial ovarian cancer: a clinicopathologic and
immunohistochemical study. Int J Gynecol Pathol. 31:125–132. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Vlahovic G, Meadows KL, Uronis HE, et al:
A phase I study of bevacizumab, everolimus and panitumumab in
advanced solid tumors. Cancer Chemother Pharmacol. 70:95–102. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Egawa-Takata T, Endo H, Fujita M, et al:
Early reduction of glucose uptake after cisplatin treatment is a
marker of cisplatin sensitivity in ovarian cancer. Cancer Sci.
101:2171–2178. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Park SS, Bae I and Lee YJ:
Flavonoids-induced accumulation of hypoxia-inducible factor
(HIF)-1alpha/2alpha is mediated through chelation of iron. J Cell
Biochem. 103:1989–1998. 2008. View Article : Google Scholar
|
|
5
|
Shin EK, Kwon HS, Kim YH, Shin HK and Kim
JK: Chrysin, a natural flavone, improves murine inflammatory bowel
diseases. Biochem Biophys Res Commun. 381:502–507. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lotito SB and Frei B: Dietary flavonoids
attenuate tumor necrosis factor alpha-induced adhesion molecule
expression in human aortic endothelial cells. Structure-function
relationships and activity after first pass metabolism. J Biol
Chem. 281:37102–37110. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lee SJ, Yoon JH and Song KS: Chrysin
inhibited stem cell factor (SCF)/c-Kit complex-induced cell
proliferation in human myeloid leukemia cells. Biochem Pharmacol.
74:215–225. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhang T, Chen X, Qu L, Wu J, Cui R and
Zhao Y: Chrysin and its phosphate ester inhibit cell proliferation
and induce apoptosis in Hela cells. Bioorg Med Chem. 12:6097–6105.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Monasterio A, Urdaci MC, Pinchuk IV,
López-Moratalla N and Martínez-Irujo JJ: Flavonoids induce
apoptosis in human leukemia U937 cells through caspase- and
caspase-calpain-dependent pathways. Nutr Cancer. 50:90–100. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Khoo BY, Chua SL and Balaram P: Apoptotic
effects of chrysin in human cancer cell lines. Int J Mol Sci.
11:2188–2199. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zheng X, Meng WD, Xu YY, Cao JG and Qing
FL: Synthesis and anticancer effect of chrysin derivatives. Bioorg
Med Chem Lett. 13:881–884. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zheng X, Cao JG, Liao DF, Zhu BY and Liu
HT: Synthesis and anticancer effect of gem-difluoromethylenated
chrysin derivatives. Chin Chem Lett. 11:1431–1442. 2006.In
Chinese.
|
|
13
|
Xiang H, Zheng X and Cao J: Induction of
apoptosis of human gastric carcinoma SGC-7901 cell line by
8-bromo-7-methoxychrysin. Zhongguo Yaolixue Tongbao. 24:1370–1373.
2008.
|
|
14
|
Ai XH, Zheng X, Tang XQ, et al: Induction
of apoptosis of human gastric carcinoma SGC-7901 cell line by 5,
7-dihydroxy-8-nitrochrysin in vitro. World J Gastroenterol.
13:3824–3828. 2007.PubMed/NCBI
|
|
15
|
Yang XH, Zheng X, Cao JG, Xiang HL, Liu F
and Lv Y: 8-Bromo-7-methoxychrysin-induced apoptosis of
hepatocellular carcinoma cells involves ROS and JNK. World J
Gastroenterol. 16:3385–3393. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Xiao G, Tang X, Yao C and Wang C:
Potentiation of arsenic trioxide-induced apoptosis by
8-bromo-7-methoxychrysin in human leukemia cells involves depletion
of intracellular reduced glutathione. Acta Biochim Biophys Sin
(Shanghai). 43:712–721. 2011. View Article : Google Scholar
|
|
17
|
Staal SP: Molecular cloning of the akt
oncogene and its human homologues AKT1 and AKT2: amplification of
AKT1 in a primary human gastric adenocarcinoma. Proc Natl Acad Sci
U S A. 84:5034–5037. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Bast RC Jr, Hennessy B and Mills GB: The
biology of ovarian cancer: new opportunities for translation. Nat
Rev Cancer. 9:415–428. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
19
|
Levanon K, Crum C and Drapkin R: New
insights into the pathogenesis of serous ovarian cancer and its
clinical impact. J Clin Oncol. 26:5284–5293. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Landen CN Jr, Birrer MJ and Sood AK: Early
events in the pathogenesis of epithelial ovarian cancer. J Clin
Oncol. 26:995–1005. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
O'Connor L, Strasser A, O'Reilly LA, et
al: Bim: a novel member of the Bcl-2 family that promotes
apoptosis. EMBO J. 17:384–395. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Qi XJ, Wildey GM and Howe PH: Evidence
that Ser87 of BimEL is phosphorylated by Akt and regulates BimEL
apoptotic function. J Biol Chem. 281:813–823. 2006. View Article : Google Scholar
|
|
23
|
Fletcher JI and Huang DC: Controlling the
cell death mediators Bax and Bak: puzzles and conundrums. Cell
Cycle. 7:39–44. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
van Gorp AG, Pomeranz KM, Birkenkamp KU,
Hui RC, Lam EW and Coffer PJ: Chronic protein kinase B (PKB/c-akt)
activation leads to apoptosis induced by oxidative stress-mediated
Foxo3a transcriptional up-regulation. Cancer Res. 66:10760–10769.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Essafi A, Fernàndez de Mattos S, Hassen
YA, et al: Direct transcriptional regulation of Bim by FoxO3a
mediates STI571-induced apoptosis in Bcr-Abl-expressing cells.
Oncogene. 24:2317–2329. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Cardone MH, Roy N, Stennicke HR, et al:
Regulation of cell death protease caspase-9 by phosphorylation.
Science. 282:1318–1321. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Sunters A, Fernàndez de Mattos S, Stahl M,
et al: FoxO3a transcriptional regulation of Bim controls apoptosis
in paclitaxel-treated breast cancer cell lines. J Biol Chem.
278:49795–49805. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Simonin K, Brotin E, Dufort S, et al:
Mcl-1 is an important determinant of the apoptotic response to the
BH3-mimetic molecule HA14-1 in cisplatin-resistant ovarian
carcinoma cells. Mol Cancer Ther. 8:3162–3170. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhang N, Wu ZM, McGowan E, et al: Arsenic
trioxide and cisplatin synergism increase cytotoxicity in human
ovarian cancer cells: therapeutic potential for ovarian cancer.
Cancer Sci. 100:2459–2464. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wang Z, Ahmad A, Banerjee S, et al: FoxM1
is a novel target of a natural agent in pancreatic cancer. Pharm
Res. 27:1159–1168. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ning Y, Li Q, Xiang H, Liu F and Cao J:
Apoptosis induced by 7-difluoromethoxyl-5,4′-di-n-octyl genistein
via the inactivation of FoxM1 in ovarian cancer cells. Oncol Rep.
27:1857–1864. 2012.PubMed/NCBI
|
|
32
|
Polier G, Ding J, Konkimalla BV, et al:
Wogonin and related natural flavones are inhibitors of CDK9 that
induce apoptosis in cancer cells by transcriptional suppression of
Mcl-1. Cell Death Dis. 2:e1822011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhao XC, Cao XC, Liu F, et al: Regulation
of the FOXO3a/Bim signaling pathway by 5,7-dihydroxy-8-nitrochrysin
in MDA-MB-453 breast cancer cells. Oncol Lett. 5:929–934.
2013.PubMed/NCBI
|
|
34
|
Yuan Z, Wang F, Zhao Z, et al:
BIM-mediated Akt phosphorylation is a key modulator of arsenic
trioxide-induced apoptosis in cisplatin-sensitive and -resistant
ovarian cancer cells. PLoS One. 6:e205862011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Chen KF, Yeh PY, Yeh KH, Lu YS, Huang SY
and Cheng AL: Down-regulation of phospho-Akt is a major molecular
determinant of bortezomib-induced apoptosis in hepatocellular
carcinoma cells. Cancer Res. 68:6698–6707. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Woo KJ, Jeong YJ, Park JW and Kwon TK:
Chrysin-induced apoptosis is mediated through caspase activation
and Akt inactivation in U937 leukemia cells. Biochem Biophys Res
Commun. 325:1215–1222. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Fraser M, Bai T and Tsang BK: Akt promotes
cisplatin resistance in human ovarian cancer cells through
inhibition of p53 phosphorylation and nuclear function. Int J
Cancer. 122:534–546. 2008. View Article : Google Scholar
|
|
38
|
Liu W, Akhand AA, Takeda K, et al: Protein
phosphatase 2A-linked and -unlinked caspase-dependent pathways for
downregulation of Akt kinase triggered by 4-hydroxynonenal. Cell
Death Differ. 10:772–781. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Ramos AM, Fernández C, Amràn D, Sancho P,
de Blas E and Aller P: Pharmacologic inhibitors of PI3K/Akt
potentiate the apoptotic action of the antileukemic drug arsenic
trioxide via glutathione depletion and increased peroxide
accumulation in myeloid leukemia cells. Blood. 105:4013–4020. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kornblau SM, Singh N, Qiu Y, et al: Highly
phosphorylated FOX03A is an adverse prognostic factor in acute
myeloid leukemia. Clin Cancer Res. 16:1865–1874. 2010. View Article : Google Scholar : PubMed/NCBI
|















