Introduction
Hepatocellular carcinoma (HCC), the primary type of
liver cancer, is one of the most malignant types of human tumor
with poor prognosis (1). HCC
accounts for >80% of all liver cancer and is diagnosed in
>600,000 individuals annually globally (2,3). HCC
has become one of the leading causes of cancer-associated mortality
worldwide (4). There is currently
no clinically proven curative therapy for advanced HCC (5), and a large percentage of cases of
advanced HCC do not respond to chemotherapies, predominantly due to
the high level of intrinsic and acquired chemoresistance (6). The successful elimination of cancer
cells through apoptosis is the ultimate aim of chemotherapy
(7,8). Apoptosis is an important process,
which controls the growth and development of organisms (9), and the perturbation of apoptosis is
considered to be a promising strategy for prevention and treatment
of HCC.
Fatsioside A, a novel baccharane-type triterpenoid
glycoside, is extracted from the fruits of Fatsia japonica
(10). Fatsioside A can exert
growth inhibition, cell cycle arrest and induce apoptosis in rat
glioma C6 cells and human glioma U251 cells (10). Therefore, fatsioside A may be a
promising novel candidate for adjunctive therapy against human
tumors through the activation of cell death. However, its action on
the HepG2 human HCC line have not been investigated and the exact
mechanisms underlying the induction of apoptosis remain to be
elucidated. The present study aimed to investigate the anticancer
properties of fatsioside A in the HepG2 human HCC line, and to
examine the underlying mechanisms by focusing on the AMP-activated
protein kinase (AMPK) signaling cascade.
Under conditions of metabolic stress, including
hypoxia, heat shock, oxidative stress and exercise, in which
adenosine triphopshate is depleted, AMPK is activated and functions
as a major metabolic switch to maintain energy homeostasis
(11–16). This conserved heterotrimeric kinase
has also been demonstrated to act as an intrinsic regulator of the
mammalian cell cycle (17). In
addition, AMPK is important in cancer cell survival and apoptosis
(18–23), and a number of anticancer medicinal
herb extracts activate AMPK-dependent cell death pathways (24–26).
Previous studies have reported that fatsioside A may be a promising
novel candidate for chemotherapy against human tumors through
inducing cell death (10),
however, the potential roles and underlying mechanisms of AMPK in
mediating fatsioside A-induced cancer cell death remain to be fully
elucidated Therefore, the present study aimed to investigate
whether AMPK activation is important for fatsioside A-induced
apoptotic death in the HepG2 HCC cells.
Materials and methods
Materials
Fatsioside A was provided by the College of
Pharmaceutical Sciences, Zhejiang University (Hangzhou, China),
which was dissolved in dimethylsulfoxide (DMSO) at 0.8 mM and
diluted with fresh medium to obtain the desired concentration. The
3-(4, 5-dimetryl-thiazol-2-yl)-2, 5-diphenyltetrazolium bromide
(MTT) assay was purchased from Cell Signaling Technology, Inc.
(Danvers, MA, USA). Antibodies specific for phosphorylated
(p-)AMPKα (rabbit anti-human polyclonal antibody; cat. no.
SAB4503754; Sigma-Aldrich, St. Louis, MO, USA), AMPKα (rabbit
anti-human polyclonal antibody; cat. no. A3730; Sigma-Aldrich),
cleaved caspase-3 (rabbit anti-human polyclonal antibody; cat. no.
C9598; Sigma-Aldrich), B-cell lymphoma (Bcl)-2 (rabbit anti-human
polyclonal antibody; cat. no. PRS3335; Sigma-Aldrich) and β-actin
(mouse anti-human monoclonal antibody; cat. no. A1978;
Sigma-Aldrich). The secondary antibodies were obtained from Santa
Cruz Biotechnology, Inc. (Dallas, TX, USA).
5-aminoimidazole-4-carboxyam ide-1-β-D-ribofuranoside (AICAR) was
purchased from Sigma-Aldrich. Z-VAD-fmk and compound C were
purchased from Calbiochem (Darmstadt, Germany).
Cell culture
The HepG2 cell line was obtained from the Chinese
Academy of Sciences Cell Bank (Shanghai, China). The cells were
maintained in Dulbecco's modified Eagle's medium (DMEM;
Sigma-Aldrich), supplemented with 10% fetal bovine serum (FBS;
Invitrogen Life Technologies, Carlsbad, CA, USA),
penicillin/streptomycin (1:100; Sigma-Aldrich) and 4 mM L-glutamine
(Sigma-Aldrich) in a CO2 incubator at 37°C. When the
cells reached 80% confluence, they were divided into three plates.
The subsequent experiments were performed when the cells reached
50–60% confluence.
MTT assay
An MTT assay was used to examine the effects of
fatsioside A on the proliferation of the HepG2 cells. Briefly, the
cells were seeded into 96-well plates at a density of
5×103 cells/well in 200 µl medium. The cells in
the wells were then treated with various concentrations (40 or 80
µM) of fatsioside A and were cultured for 24 h at 37°C. At
the end of culture, 0.5 mg/ml MTT in 20 µl
phosphate-buffered saline (PBS) was added to each well and the
cells were incubated for 4 h at 37°C. An enzyme-labeled instrument
(Thermo Fisher Scientific, Waltham, MA, USA) was used to measure
the absorbance of each well at 570 nm. Data were calculated from
three independent experiments.
Cell apoptosis assay
An Annexin V Apoptosis Detection kit (Beyotime
Institute of Biotechnology, Shanghai, China) was used to analyze
the effects of fatsioside A on the apoptosis of the HepG2 cells.
Briefly, 1,000,000 HepG2 cells with the indicated treatment were
stained with fluorescein isothiocyanate (FITC)-Annexin V and
propidium iodide (PI). The early (annexin V+/PI−) and late (annexin
V+/PI+) apoptotic cells were sorted using a fluorescence-activated
cell sorting machine (FACSCalibur; BD Biosciences, Franklin Lakes,
NJ, USA). All experiments were performed in triplicate.
BrdU incorporation assay
The HepG2 cells were seeded at a density of
1×105 cells/well in 0.5 ml DMEM containing 10% FBS into
48-well tissue culture plates. The cells were serum-starved for 24
h and then exposed to various concentrations (40 or 80 µM)
of fatsioside A for 24 h at 37°C. The cell proliferation was
assessed by examining the incorporation of BrdU using a BrdU ELISA
colorimetric assay (Roche Diagnostics, Indianapolis, IN, USA),
according to the manufacturer's instructions. The ELISA optical
density value of the treatment groups were normalized to that of
the untreated control group. Each condition was assessed in
triplicate.
Trypan blue staining
The numbers of 'dead' HepG2 cells (trypan
blue-positive) following the indicated treatments were recorded,
and the percentage of cell death in the HepG2 was calculated by the
number of the trypan blue dye-positive cells divided by the total
number of the cells.
TUNEL staining
Apoptotic cells was also detected using a TUNEL kit
(Sigma-Aldrich) according to the manufacturer's instructions.
Briefly, cells were cultured on cover slips. After Fatsioside A (40
or 80 µM) treatment for 24 h, the cells were fixed in 4%
paraformaldehyde solution in PBS for 30 min at room temperature.
The cells were then incubated with a methanol solution containing
0.3% H2O2 for 30 min at room temperature to
block endogenous peroxidase activity, and then incubated in the
TUNEL reaction mixture for 60 min at 37°C, counterstained with
DAPI, then finally visualized by fluorescence microscopy (DM4000B;
Leica Microsystems GmbH, Wetzlar, Germany).
Western blot analysis
Whole cellular protein was extracted from the HepG2
cells using lysis buffer for western blot analysis. Briefly, the
cells were lysed in radioimmunoprecipitation assay buffer
(Sigma-Aldrich) for 30 min on ice. Protein levels were quantified
using the bicinchoninic acid method. Equal quantities of protein
(30 µg/lane) were separated by 5–12% sodium dodecyl
sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and then
transferred onto nitrocellulose blotting membranes (EMD Millipore,
Billerica, MA, USA). The membranes were blocked in tris-buffered
saline with Tween 20 (Sigma-Aldrich) containing 5% non-fat dry milk
(w/v) for 2 h and incubated with primary antibodies at 1:1,000
dilution overnight at 4°C, followed by incubation with
corresponding horseradish peroxidase-conjugated secondary
antibodies at room temperature for 2 h. Protein bands were
visualized using chemiluminescence detection (EMD Millipore).
RNA interference (RNAi)
The following RNAi sequences:
5′-GCAUAUGCUGCAGGUAGAU-3′ and 5′-AAGGAAAGTGAAGGTGGGCAA-3′, targeted
against human AMPKα1/2 were synthesized by Genewiz, Inc. (Suzhou,
China). Non-sense control RNAi was purchased from Santa Cruz
Biotechnology, Inc, and was used as an RNAi-negative control.
Transfection was performed, as described previously (27). Briefly, the HepG2 cells were
cultured on a six-well plate to 60% confluence in antibiotic- and
serum-free medium at 37°C. The targeted and control RNAi (100
µM) and 3.0 µl Lipofectamine PLUS Reagent (Invitrogen
Life Technologies) were diluted in 90 µl small interfering
RNA dilution buffer (Santa Cruz Biotechnology, Inc.), and 3
µl Lipofectamine LTX (Sigma-Aldrich) was added. The
transfection complex was then added to the wells containing 1 ml
DMEM for 12 h, with a final RNAi concentration of 100 nM. Growth
medium was then added to the cells, which were cultured for an
additional 48 h at 37°C. The expression levels of the target
proteins in the transfected cells were assessed using western blot
analysis. Only the cells exhibiting significant target
protein-knockdown were used for the experiments.
Statistical analysis
The data are presented as the mean ± standard
deviation of three independent experiments. Differences between two
mean values were evaluated using Student's t-test and
P<0.05 was considered to indicate a statistically significant
difference. Statistical analyses were performed using SPSS software
19.0 (SPSS, Inc., Chicago, IL, USA).
Results
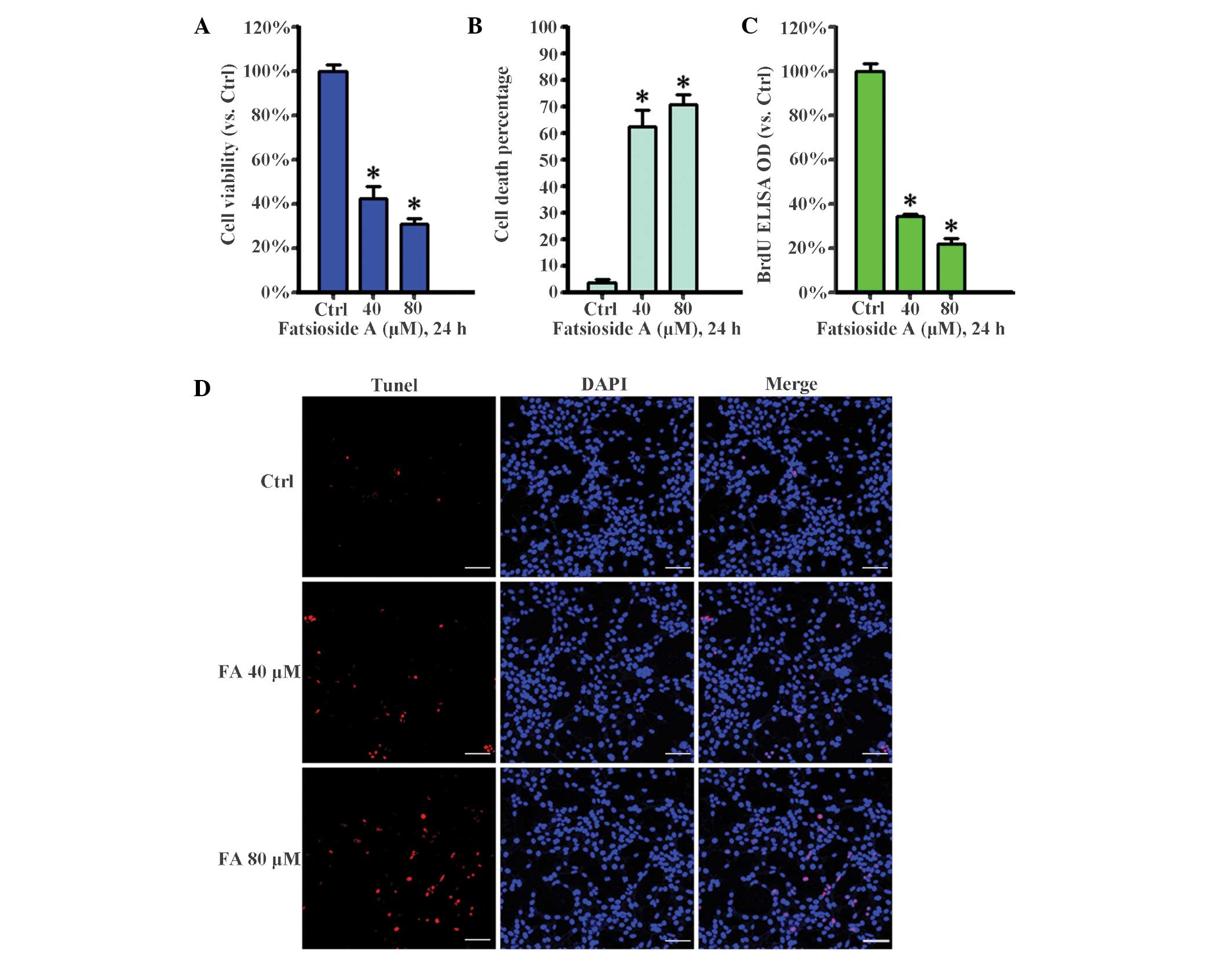
Fatsioside A inhibits the survival and
proliferation of HepG2 cells
The present study examined the effect of fatsioside
A on HepG2 cell survival and proliferation. Cell viability was
evaluated using an MTT assay. The results, as shown in Fig. 1A, clearly indicated that fatsioside
A at concentrations of 40 and 80 µM markedly inhibited HepG2
cell survival. In addition, the number of trypan blue-positive,
'dead', cells increased sharply following fatsioside A treatment at
40 and 80 µM (Fig. 1B). The
effect of fatsioside A on HepG2 cell proliferation was also
examined. Using a BrdU incorporation assay, the present study
demonstrated that fatsioside A suppressed HepG2 cell proliferation
(Fig. 1C). Furthermore, a TUNEL
assay revealed that, compared with the control group, the number of
tunnel-positive 'dead' cells increased markedly following
fatsioside A treatment at concentrations of 40 and 80 µM
(Fig. 1D). Taken together, these
results suggested that fatsioside A significantly inhibited the
survival and proliferation of the HepG2 cells.
Fatsioside A induces apoptotic and
necrotic death of HepG2 cells
The results described above indicated that
fatsioside A inhibited HepG2 cell survival and proliferation;
therefore, the present study subsequently assessed whether cell
apoptosis was involved in this effect. As shown in Fig. 2A and B, fatsioside A (40 and 80
µM) induced early (Annexin V+/PI−) and late (Annexin V+/PI+)
apoptosis in the HepG2 cells. In addition, fatsioside A also caused
caspase-3 cleavage and Bcl-2 degradation (Fig. 2C). Notably, fatsioside A also
induced necrotic (Annexin V−/PI+) HepG2 cell death (Fig. 2A and B). Furthermore, the results
of the cell viability assay results, as shown in Fig. 2D, demonstrated that z-VAD-fmk, a
general caspase inhibitor, suppressed, but did not reverse,
fatsioside A-induced HepG2 loss of viability, indicating that
apoptotic and necrotic death accounted for the fatsioside A-induced
cytotoxicity in the HepG2 cells.
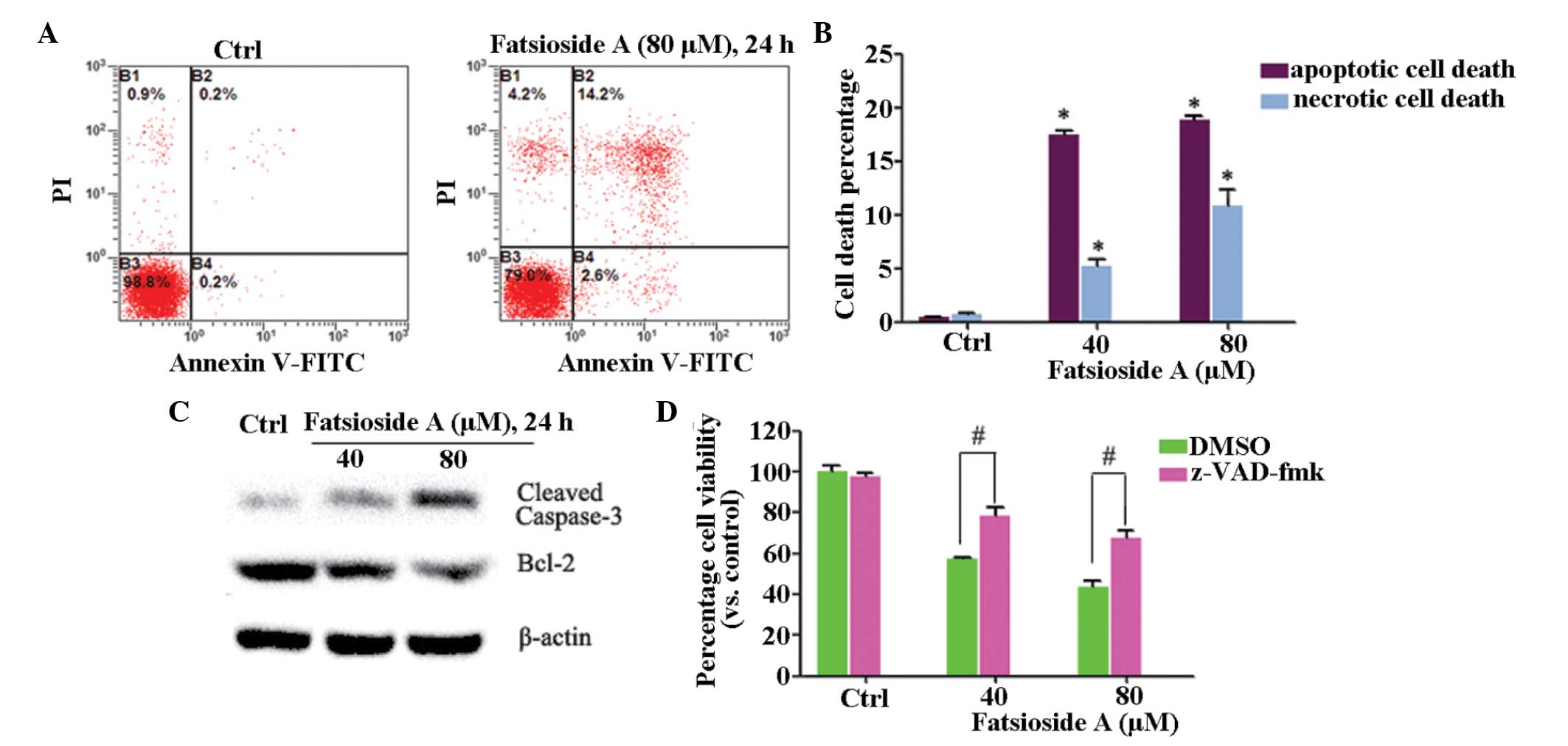 | Figure 2Fatsioside A induces apoptotic and
necrotic death of HepG2 cells. (A and B) HepG2 cells were treated
with or without fatsioside A and were cultured in Dulbecco's
modified Eagle's medium for 24 h, following which apoptotic and
necrotic cell death was assessed using an Annexin V
fluorescence-activated cell sorting. (C) Expression levels of
cleaved caspase-3, Bcl-2 and β-actin were assessed using western
blot analysis. (D) HepG2 cells were pre-treated with z-VAD-fmk (50
µM), a general caspase inhibitor, for 1 h, followed by
fatsioside A (40 and 80 µM) stimulation and further culture
for 24 h, Cell viability was then assessed using a 3-(4,
5-dimetryl-thiazol-2-yl)-2, 5-diphenyltetrazolium bromide assay.
Experiments were repeated three times and similar results were
obtained. Data are expressed as the mean ± standard deviation.
*P<0.05, vs. Ctrl; #P<0.05, vs.
fatsioside A group. Ctrl, untreated control; PI, propidium iodide;
FITC, fluorescein isothiocyanate; DMSO, dimethyl sulfoxide; Bcl-2,
B-cell lymphoma-2. |
Activation of AMPK is involved in
fatsioside A-induced cytotoxicity in HepG2 cells
As shown in Fig.
3A, fatsioside A induced significant AMPK activation in the
HepG2 cells, as the expression levels of p-AMPKα and the downstream
ACC in HepG2 cells were significantly increased following
fatsioside A treatment. Notably, AMPK inhibition by its inhibitor,
compound C or by AMPKα-RNAi suppressed fatsioside A-induced loss of
cell viability (Fig. 3B and C).
Fatsioside A-induced apoptosis and cleavage of caspase-3 was also
inhibited by AMPK inhibition (Fig. 3D
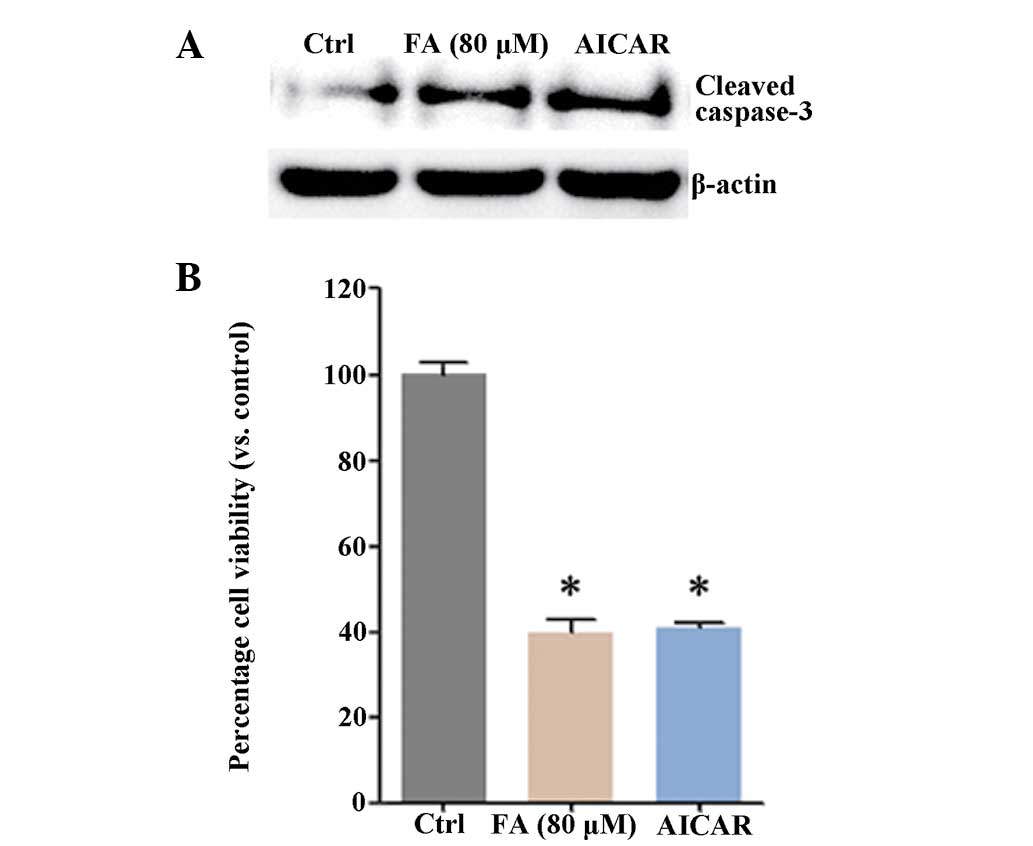
and E). The AICAR AMPK activator also inhibited HepG2 cell
survival (Fig. 4A and B). The
above results indicated that activation of AMPK was involved in
fatsioside A-induced cytotoxicity in HepG2 cells.
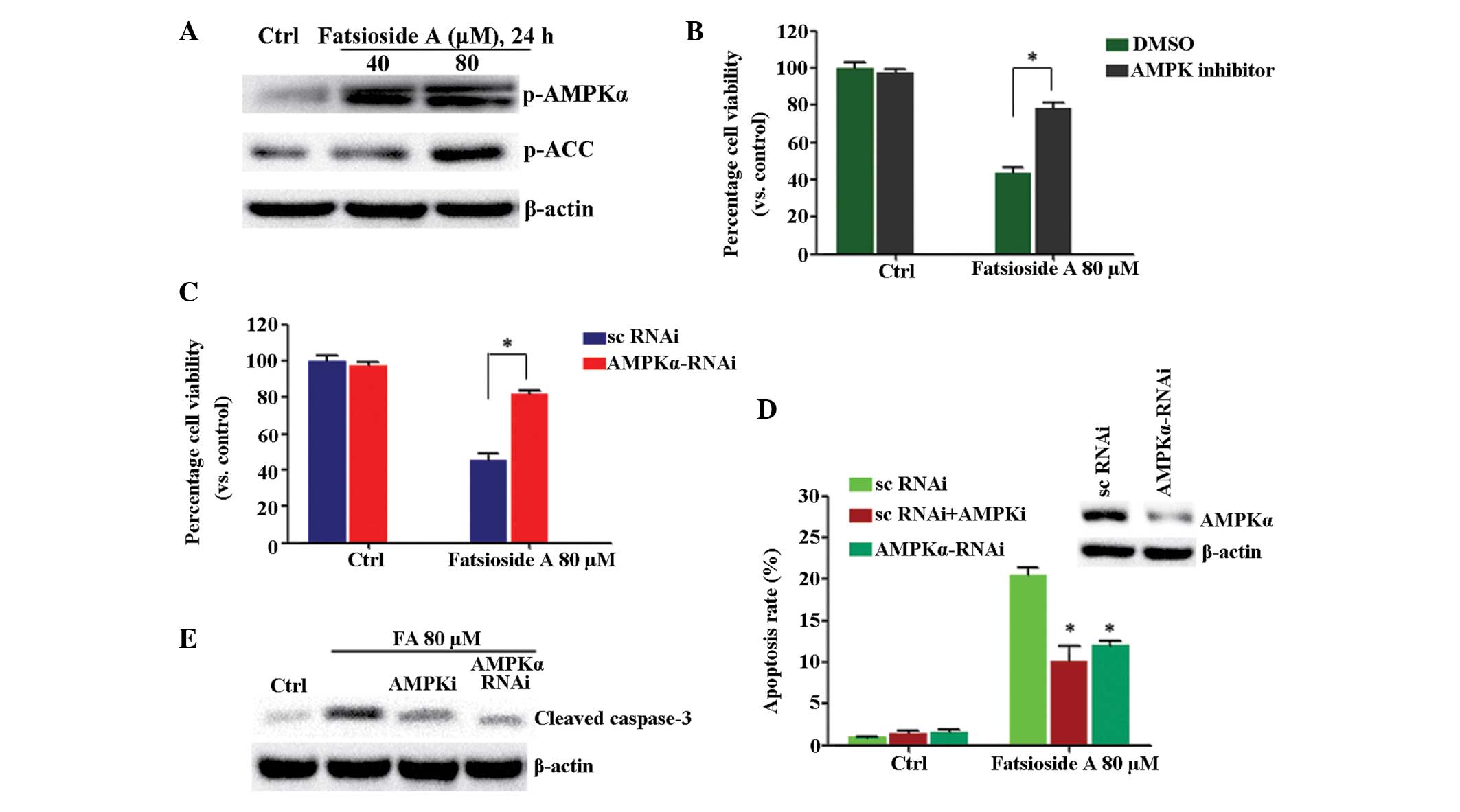 | Figure 3AMPK inhibition suppresses fatsioside
A-induced cell viability loss in HepG2 cells. (A) HepG2 cells were
either left untreated or were treated with different concentrations
of fatsioside A (40 or 80 µM) for 24 h. The phosphorylation
of AMPKα and ACC were then assessed using western blot analysis.
(B) HepG2 cells were pre-treated with the AMPK inhibitor, compound
C (10 µM), for 1 h, followed by fatsioside A (80 µM)
stimulation, and were further cultured for 24 h prior to assessment
of cell viability using an MTT assay. (C) Scramble control
(sc)RNAi- or AMPKα RNAi-transfected HepG2 cells were either left
untreated or were treated with fatsioside A (80 µM), and
were further cultured for 24 h prior to cell viability assessment
using an MTT assay. (D) Expression levels of AMPKα in these cells
were assessed using western blot analysis (upper) and were also
assessed for cell apoptosis using Annexin V fluorescence-activated
cell sorting 24 h, following treatment with fatsioside A (80
µM).*P<0.05, vs. fatsioside A + scRNAi group.
(E) Expression levels of cleaved caspase-3 in the cells were
examined using western blot analysis. Experiments were repeated
three times and similar results were obtained. Ctrl, untreated
control; FA, fatsopside A; AMPK, AMP-activated protein kinase; ACC,
acetyl-CoA carboxylase carboxylase; p-, phosphorylated; RNAi, RNA
interference; DMSO, dimethyl sulfoxide. |
Discussion
HCC is known to be one of the most life-threatening
types of tumor in humans. The effects of current antitumor
therapies in HCC are limited (28–30)
and, as HCC cells are resistant to apoptosis, patients with HCC
usually have a poor prognosis. Therefore, examination of potential
novel therapy targeting their inherent apoptosis-resistant
phenotype is essential.
The present study revealed that fatsioside A
markedly inhibited the survival and proliferation of HepG2 cells
and induced apoptotic and necrotic death of the HepG2 cells.
Furthermore, this effect was found to be exerted by activation of
the AMPK cascade.
In line with previously published data on glioma
(10), the present study observed
that fatsioside A significantly inhibited the proliferation of
HepG2 cells. One of the novel findings of the present study is the
confirmation that the fatsioside A-induced reduction in the
viability of HepG2 cells occurred through apoptosis, which was
elucidated following the application of Annexin V-FITC/PI double
staining. Apoptosis is a physiological phenomenon (31). The significance of apoptosis is to
remove senescent cells and over functioning cells, including
activated T cells (32–34). Deregulation of apoptosis is
associated with the pathogenesis of a number of disorders,
including tumor cell growth. Thus, one of the predominant
strategies used to treat tumors is to induce the apoptosis of tumor
cells (35). The results of the
present study suggested that fatsioside A can disturb the inherent
apoptosis-resistant ability of HepG2 cells. Of note, it was also
found that fatsioside A induced necrotic death of the HepG2 cells,
as the z-VAD-fmk caspase inhibitor suppressed, but did not reverse,
fatsioside A-induced loss of HepG2 viability.
Previous studies have demonstrated that cellular
stress-activated AMPK promotes cell apoptosis (36), and such an effect by AMPK occurs
through regulating the downstream signals of AMPK, including c-Jun
N-terminal kinase, p53 and mammalian target of rapamycin (37–39).
In addition, anticancer chemotherapies, including taxol and
temozolomide activate the AMPK-dependent apoptosis pathways
(40,41). Resveratrol, capsaicin and EGCG
anticancer plant extract-induced cancer cell death also requires
AMPK activation (18,42,43).
In the present study, significant AMPK activation was observed in
the fatsioside A-treated HepG2 cells. Inhibition of AMPK by RNAi or
compound C suppressed fatsioside A-induced apoptosis in the HepG2
cells. By contrast, HepG2 cell viability was inhibited by the AICAR
AMPK activator. These results suggested that AMPK activation is
required for the induction of the anticancer effects of fatsioside
A in HepG2 cells.
However, of note, AMPK inhibition reduced, but did
not reverse HepG2 cytotoxicity-induced by fatsioside A in the
present study. This may be due to the incomplete inhibition of AMPK
by the methods used in the present study (RNAi or compound C),
however, it is more likely that AMPK activation is among several
mechanisms activated by fatsioside A to mediate HepG2 cell death.
Other signals, which are independent of AMPK activation and induced
by fatsioside A require further investigation. Although the present
study confirmed AMPK activation by fatsioside A, the potential
upstream signal for this activation remains to be elucidated.
Acknowledgments
The authors would like to thank Dr Liu from the
Department of Pharmacology and Neurology, Emory University
(Atlanta, GA, USA) for the critical reading and modification of the
manuscript.
References
|
1
|
Zhang JP, Yan J, Xu J, Pang XH, Chen MS,
Li L, Wu C, Li SP and Zheng L: Increased intratumoral
IL-17-producing cells correlate with poor survival in
hepatocellular carcinoma patients. J Hepatol. 50:980–989. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Srivatanakul P, Sriplung H and Deerasamee
S: Epidemiology of liver cancer: An overview. Asian Pac J Cancer
Prev. 5:118–125. 2004.PubMed/NCBI
|
|
3
|
Ferenci P, Fried M, Labrecque D, Bruix J,
Sherman M, Omata M, Heathcote J, Piratsivuth T, Kew M, Otegbayo JA,
et al: Hepatocellular carcinoma (HCC): A global perspective. J Clin
Gastroenterol. 44:239–245. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yang JD and Roberts LR: Hepatocellular
carcinoma: A global view. Nat Rev Gastroenterol Hepatol. 7:448–458.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Llovet JM: Updated treatment approach to
hepatocellular carcinoma. J Gastroenterol. 40:225–235. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Villanueva A and Llovet JM: Targeted
therapies for hepatocellular carcinoma. Gastroenterology.
140:1410–1426. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sun Y and Peng Z: Programmed cell death
and cancer. Postgrad Med J. 85:134–140. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ashkenazi A and Herbst RS: To kill a tumor
cell: The potential of proapoptotic receptor agonists. J Clin
Invest. 118:1979–1990. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Pérez-Garijo A, Martín FA and Morata G:
Caspase inhibition during apoptosis causes abnormal signalling and
developmental aberrations in Drosophila. Development.
131:5591–5598. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yu S, Ye X, Xin W, Xu K, Lian XY and Zhang
Z: Fatsioside A, a rare baccharane-type glycoside inhibiting the
growth of glioma cells from the fruits of fatsia japonica. Planta
Med. 80:315–320. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Papandreou I, Lim AL, Laderoute K and
Denko N: Hypoxia signals autophagy in tumor cells via AMPK
activity, independent of HIF-1, BNIP3 and BNIP3L. Cell Death
Differ. 15:1572–1581. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Laderoute KR, Amin K, Calaoagan JM, Knapp
M, Le T, Orduna J, Foretz M and Viollet B: 5-AMP-activated protein
kinase (AMPK) is induced by low-oxygen and glucose deprivation
conditions found in solid-tumor microenvironments. Mol Cell Biol.
26:5336–5347. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Schulz E, Anter E, Zou MH and Keaney JF
Jr: Estradiol-mediated endothelial nitric oxide synthase
association with heat shock protein 90 requires adenosine
monophosphate-dependent protein kinase. Circulation. 111:3473–3480.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ceolotto G, Gallo A, Papparella I, Franco
L, Murphy E, Iori E, Pagnin E, Fadini GP, Albiero M, Semplicini A
and Avogaro A: Rosiglitazone reduces glucose-induced oxidative
stress mediated by NAD (P) H oxidase via AMPK-dependent mechanism.
Arterioscl Throm Vasc Biol. 27:2627–2633. 2007. View Article : Google Scholar
|
|
15
|
Horie T, Ono K, Nagao K, Nishi H,
Kinoshita M, Kawamura T, Wada H, Shimatsu A, Kita T and Hasegawa K:
Oxidative stress induces GLUT4 translocation by activation of
PI3K/Akt and dual AMPK kinase in cardiac myocytes. J Cell Physiol.
215:733–742. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Jorgensen SB and Rose AJ: How is AMPK
activity regulated in skeletal muscles during exercise? Front
Biosci. 13:5589–5604. 2007.
|
|
17
|
Jones RG, Plas DR, Kubek S, Buzzai M, Mu
J, Xu Y, Birnbaum MJ and Thompson CB: AMP-activated protein kinase
induces a p53-dependent metabolic checkpoint. Mol Cell. 18:283–293.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hwang JT, Ha J, Park IJ, Lee SK, Baik HW,
Kim YM and Park OJ: Apoptotic effect of EGCG in HT-29 colon cancer
cells via AMPK signal pathway. Cancer Lett. 247:115–121. 2007.
View Article : Google Scholar
|
|
19
|
Hwang JT, Ha J and Park OJ: Combination of
5-fluorouracil and genistein induces apoptosis synergistically in
chemo-resistant cancer cells through the modulation of AMPK and
COX-2 signaling pathways. Biochem Bioph Res Commun. 332:433–440.
2005. View Article : Google Scholar
|
|
20
|
Shaw RJ, Kosmatka M, Bardeesy N, Hurley
RL, Witters LA, DePinho RA and Cantley LC: The tumor suppressor
LKB1 kinase directly activates AMP-activated kinase and regulates
apoptosis in response to energy stress. Proc Natl Acad Sci USA.
101:3329–3335. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sid B, Glorieux C, Valenzuela M,
Rommelaere G, Najimi M, Dejeans N, Renard P, Verrax J and Calderon
PB: AICAR induces Nrf2 activation by an AMPK-independent mechanism
in hepato-carcinoma cells. Biochem Pharmacol. 91:168–180. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Xu Q, Liu X, Zheng X, Yao Y, Wang M and
Liu Q: The transcriptional activity of Gli1 is negatively regulated
by AMPK through Hedgehog partial agonism in hepatocellular
carcinoma. Int J Mol Med. 34:733–741. 2014.PubMed/NCBI
|
|
23
|
Cheng J, Huang T, Li Y, Guo Y, Zhu Y, Wang
Q, Tan X, Chen W, Zhang Y, Cheng W, et al: AMP-activated protein
kinase suppresses the in vitro and in vivo proliferation of
hepatocellular carcinoma. PLoS One. 9:e932562014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Johnson JJ: Carnosol: A promising
anti-cancer and anti-inflammatory agent. Cancer Lett. 305:1–7.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kim T, Davis J, Zhang AJ, He X and Mathews
ST: Curcumin activates AMPK and suppresses gluconeogenic gene
expression in hepatoma cells. Biochem Biophys Res Commun.
388:377–382. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
HemaIswarya S and Doble M: Potential
synergism of natural products in the treatment of cancer. Phytother
Res. 20:239–249. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chiu YL, Ali A, Chu CY, Cao H and Rana TM:
Visualizing a correlation between siRNA localization, cellular
uptake, and RNAi in living cells. Chem Biol. 11:1165–1175. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Bruix J and Sherman M; American
Association for the Study of Liver Diseases: Management of
hepatocellular carcinoma: An update. Hepatology. 53:1020–1022.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Bruix J and Sherman M; Practice Guidelines
Committee; American Association for the Study of Liver Diseases:
Management of hepatocellular carcinoma. Hepatology. 42:1208–1236.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
El-Serag HB and Rudolph KL: Hepatocellular
carcinoma: Epidemiology and molecular carcinogenesis.
Gastroenterology. 132:2557–2576. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Elmore S: Apoptosis: A review of
programmed cell death. Toxicol Pathol. 35:495–516. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Skulachev V: Bioenergetic aspects of
apoptosis, necrosis and mitoptosis. Apoptosis. 11:473–485. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Hunter AM, LaCasse EC and Korneluk RG: The
inhibitors of apoptosis (IAPs) as cancer targets. Apoptosis.
12:1543–1568. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Plumas J, Chaperot L, Richard MJ, Molens
JP, Bensa JC and Favrot MC: Mesenchymal stem cells induce apoptosis
of activated T cells. Leukemia. 19:1597–1604. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Fesik SW: Promoting apoptosis as a
strategy for cancer drug discovery. Nat Rev Cancer. 5:876–885.
2005. View
Article : Google Scholar : PubMed/NCBI
|
|
36
|
Motoshima H, Goldstein BJ, Igata M and
Araki E: AMPK and cell proliferation-AMPK as a therapeutic target
for atherosclerosis and cancer. J Physiol. 574:63–71. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Lee YM, Uhm KO, Lee ES, Kwon J, Park SH
and Kim HS: AM251 suppresses the viability of HepG2 cells through
the AMPK (AMP-activated protein kinase)-JNK (c-Jun N-terminal
kinase)-ATF3 (activating transcription factor 3) pathway. Biochem
Bioph Res Commun. 370:641–645. 2008. View Article : Google Scholar
|
|
38
|
Duan X, Ponomareva L, Veeranki S and
Choubey D: IFI16 induction by glucose restriction in human
fibroblasts contributes to autophagy through activation of the
ATM/AMPK/p53 pathway. Plos One. 6:e195322011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Sugiyama M, Takahashi H, Hosono K, Endo H,
Kato S, Yoneda K, Nozaki Y, Fujita K, Yoneda M, Wada K, et al:
Adiponectin inhibits colorectal cancer cell growth through the
AMPK/mTOR pathway. Int J Oncol. 34:339–344. 2009.PubMed/NCBI
|
|
40
|
Rocha GZ, Dias MM, Ropelle ER,
Osório-Costa F, Rossato FA, Vercesi AE, Saad MJ and Carvalheira JB:
Metformin amplifies chemotherapy-induced AMPK activation and
antitumoral growth. Clin Cancer Res. 17:3993–4005. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zhang WB, Wang Z, Shu F, Jin YH, Liu HY,
Wang QJ and Yang Y: Activation of AMP-activated protein kinase by
temozolomide contributes to apoptosis in glioblastoma cells via p53
activation and mTORC1 inhibition. J Biol Chem. 285:40461–40471.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Hwang JT, Kwak DW, Lin SK, Kim HM, Kim YM
and Park OJ: Resveratrol induces apoptosis in chemoresistant cancer
cells via modulation of AMPK signaling pathway. Ann NY Acad Sci.
1095:441–448. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Kim YM, Hwang JT, Kwak DW, Lee YK and Park
OJ: Involvement of AMPK signaling cascade in capsaicin-induced
apoptosis of ht-29 colon cancer cells. Ann NY Acad Sci.
1095:496–503. 2007. View Article : Google Scholar : PubMed/NCBI
|