Introduction
Colon cancer is a particularly common type of cancer
with 655,000 mortalities per year (1,2).
Worldwide, colon cancer remains a major cause of cancer-related
mortality (3,4). Despite recent developments of novel
treatment strategies, the five-year survival rate of advanced colon
cancer patients remains particularly poor, due to insensitivity to
chemotherapy and susceptibility to recurrence (5–7).
Thus, novel therapeutic agents are required to treat colon
cancer.
Numerous microtubule-targeting agents that are
currently approved or are in clinical development (including
paclitaxel, vinblastine, vincristine, colchicine and
combretastatin) are plant-derived compounds (8). Cochicine, which is a natural
compound, can be isolated from either Colchicum autumnale
(meadow saffron) or Gloriosa superba (glory lily), which
belong to the lily family (9).
Colchicine prevents growth of cancer cells by antimitotic activity
through interacting with microtubules, and may contribute to the
establishment of improved cancer therapies (10–11).
Furthermore, colchicine preferentially binds to unpolymerized
tubulin heterodimers in solution, forming a stable complex that
effectively inhibits microtubule dynamics upon binding to
microtubule ends (12). Colchicine
causes microtubule depolymerization by inhibiting lateral contacts
between protofilaments (13).
Although colchicine exerts potent microtubule depolymerization
activity and possesses antimitotic properties, its mechanism of
inducing colon cancer cell apoptosis remains unclear.
The aim of the present study was to investigate the
molecular mechanism by which colchicine induces apoptosis in colon
cancer. HT-29 colon cancer cells were treated with varying
concentration of colchicine and cell proliferation was analyzed
using a Cell Counting Kit-8 (CCK-8). Apoptosis was assayed by flow
cytometry and Hoechst 33342 straining. In addition, the
mitochondrial membrane potential (Δψm) and reactive oxygen species
(ROS) were assayed by flow cytometry. In addition,
apoptosis-associated proteins were examined by western blot. The
present study investigated the apoptosis-inducing effects of
colchicine in colon cancer cells as well as the underlying
mechanism.
Materials and methods
Reagents
Purified colchicine (98.79%) was procured from
Nanjing Zelang Medical Technological Co. Ltd. (Nanjing, China).
Antibodies for p38 (cat. no. 9212), phosphorylated (P)-p38 (cat.
no. 4092), c-Jun N-terminal kinase (JNK) (cat. no. 9252), P-JNK
(cat. no. 81E11), extracellular signal-regulated kinase (ERK) (cat.
no. 9107), P-ERK (cat. no. 4370), AKT (cat. no. 9272) and P-AKT
(cat. no. 587F11) were purchased from Cell Signaling Technology,
Inc. (Danvers, MA, USA) and GAPDH (cat. no. ab181602) was obtained
from Thermo Fisher Scientific, Inc. (Pittsburgh, PA, USA).
Antibodies for caspase-3 (cat. no. ab32499), caspase-9 (cat. no.
BMS1041) and Apaf-1 (cat. no. MA515743) were purchased from Abcam
(Cambridge, MA, USA) and Hoechst 33258 and an AlexaFluor
488-conjugated Annexin V Apoptosis Detection kit were obtained from
Molecular Probes Life Technologies (Carlsbad, CA, USA).
Cell culture
The human colon cancer HT-29 cell line was obtained
from Shanghai Institute of Cell Biology (Shanghai, China). Cells
were cultured in RPMI-1640 medium with 10% fetal bovine serum
(Gibco Life Technologies, Carlsbad, CA, USA), 100 U/ml penicillin G
and 100 μg/ml streptomycin in an incubator at 37°C, 100%
humidity and an atmosphere of 5% CO2. The growth medium
was replaced every 2–3 days.
CCK-8 assay
The CCK-8 (Dojindo Molecular Technologies, Inc.,
Shanghai, China) was used to determine cell viability according to
the manufacturer's instructions. Cells were plated at a density of
5×103 cells/well in 96-well microtiter plates and
cultured overnight at 37°C in a humidified incubator with a 5%
CO2 atmosphere. Following treatment with colchicine for
24, 48 or 72 h, the culture medium was replaced with 100 μl
fresh medium followed by the addition of 10 μl CCK-8
solution. The cells were incubated for a further 2 h at 37°C, and
the optical density was recorded at an absorbance of 450 nm. All
experiments were performed in triplicate.
Cell apoptosis assay
Cell apoptosis was detected by an Annexin V-(f
luorescein isothiocyanate) FITC/propidium iodide (PI) Apoptosis
Detection kit I (BD Biosciences, Franklin Lakes, NJ, USA) according
to the manufacturer's instructions. Cells were seeded in 6-well
plates at the density of 1×105 cells/well. Following
incubation for 24 h at 37°C, colchicine was added to the 6-well
plate and incubated for another 24 h. Cells were collected and
washed twice with phosphate-buffered saline (PBS) and resuspended,
subsequently aliquots of 1×105 cells were transferred
into 5-ml culture tubes in 100 μl 1X binding buffer (BD
Biosciences). Annexin V-FITC (5 μl) and 5 μl PI were
added to the resuspended cells. Following incubation at room
temperature for 15 min in the dark, 400 μl binding buffer
was added to the resuspended cells. Flow cytometry (FACSCalibur; BD
Biosciences) was used to assess the apoptotic cells. The
quantitation of apoptotic cells was calculated using CellQuest Pro
software version 5.1 (BD Biosciences).
Hoechst staining
The nuclei were stained using Hoechst 33342 dye
(Molecular Probes Life Technologies) to visualize nuclear
morphology, as well as the induction of apoptosis. Following
treatment with colchicine, the cells were incubated with 10
μM Hoechst 33342 dye for 10 min at 37°C. Images were
obtained using a Leica DM IRB inverted fluorescence microscope
(2886; Leica Microsystems GmbH, Wetzlar, Germany; magnification,
×400).
Mitochondrial membrane potential (Δψm)
analysis
Δψm was analyzed using a JC-1 Δψm assay kit (Cayman
Chemical Company, Ann Arbor, MI, USA), according to the
manufacturer's instructions. In brief, HT-29 cells were incubated
with 0, 1, 10 and 20 μg/ml colchicine for 12 h, centrifuged
at 1,200 × g for 5 min and resuspended in RPMI-1640 medium.
Following the addition of JC-1 to each well, the cells were
incubated for an additional 30 min at 37°C. After washing with PBS,
the stained cells were assayed using a flow cytometer (FACSCalibur;
BD Biosciences).
Mitochondrial isolation and measurement
of ROS production
Mitochondria were isolated from the untreated HT-29
cells to investigate the effect of colchicine on ROS production and
the possibility of the mitochondria as a target of colchicine.
Cells were washed twice in cold PBS, resuspended in hypotonic
buffer (1 mM EDTA, 5 mM Tris-HCl (pH 7.5), 210 mM mannitol, 70 mM
sucrose, 10 μM Leu-pep, 10 μM Pep-A and100 μM
phenylmethanesulfonylfluoride; Molecular Probes Life Technologies),
homogenized, then centrifuged at 500 × g for 5 min at 4°C to pellet
the nuclear fraction. The supernatant was centrifuged at 12,000 rpm
for 15 min at 4°C. The cytosolic supernatant was disposed of and
the pellet (containing mitochondria) was resuspended in cold
reaction buffer and treated with 10 μM colchicine. Amplex
Red (Molecular Probes Life Technologies) in combination with
horse-radish peroxidase was used to assess mitochondrial ROS
production. Fluorescence (excitation, 560 nm; emission, 590 nm) was
measured using a spectrofluorometer (SpectraMax Gemini XS;
Molecular Devices, LLC, Sunnyvale, CA, USA) every 5 min for 5 h.
The readings were analyzed using GraphPad Prism 6.0 (GraphPad Inc.,
La Jolla, CA, USA) and expressed as relative fluorescence units per
microgram of protein. The data are representative of three
independent experiments.
Statistical analysis
All experiments were repeated at least three times
and values are expressed as the mean ± standard error. Statistical
analysis was performed using two-way analysis of variance with
GraphPad Prism 6.0 software. P<0.05 was considered to indicate a
statistically significant difference between values.
Results
Colchicine inhibits proliferation of
colon cancer cells
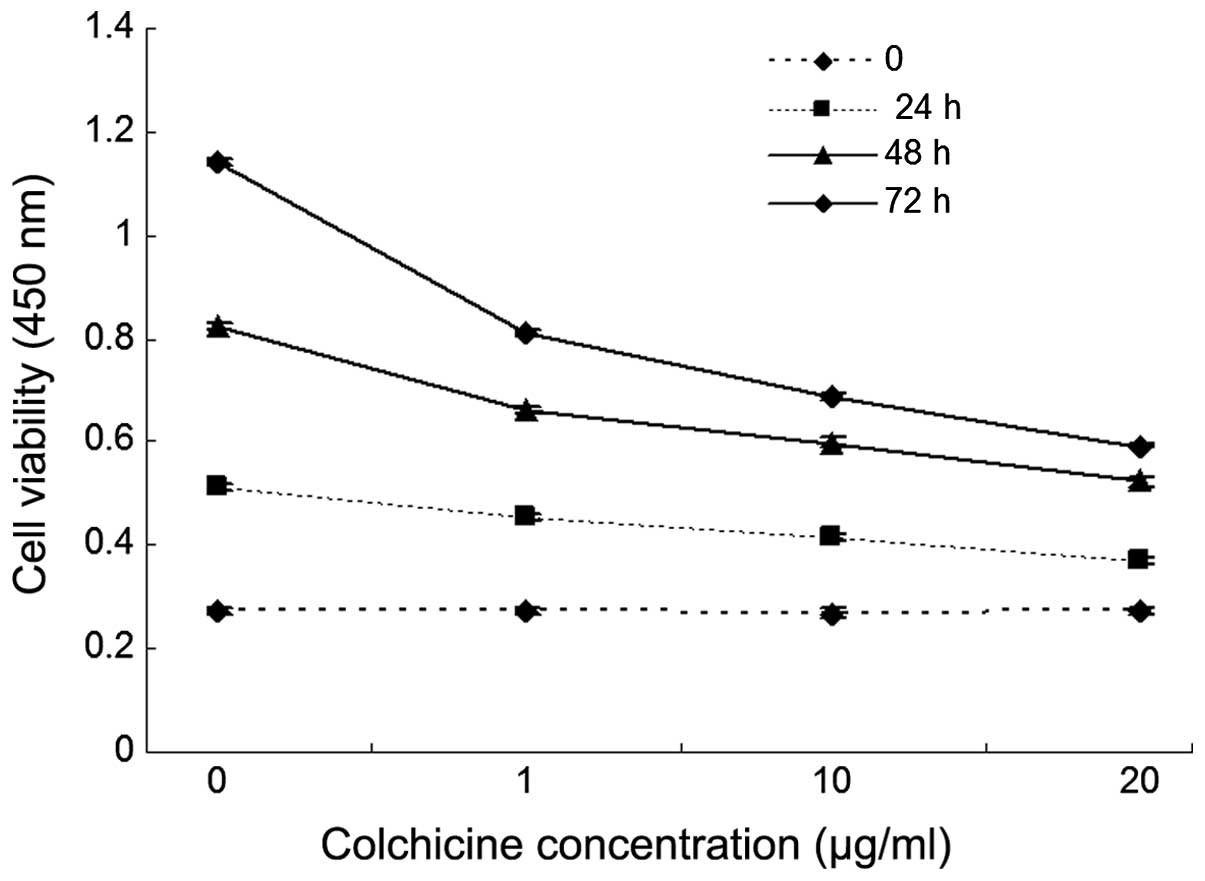
Colchicine inhibits the proliferation of colon
cancer cells in vitro. HT-29 cells were incubated with
increasing doses of colchicine (0, 1, 10 and 20 ug/ml) or a vehicle
control for 48 h, and cell viability was measured using a CCK-8
assay. As shown in Fig. 1,
colchicine significantly inhibited the proliferation of HT-29 cells
when the cells were treated with >20 μg/ml colchicine,
compared with the control, and the cell viability decreased
significantly in a dose-dependent manner. In addition, the result
revealed that colchicine inhibited HT-29 cell proliferation in a
time-dependent manner.
Colchicine induces apoptosis of colon
cancer cells
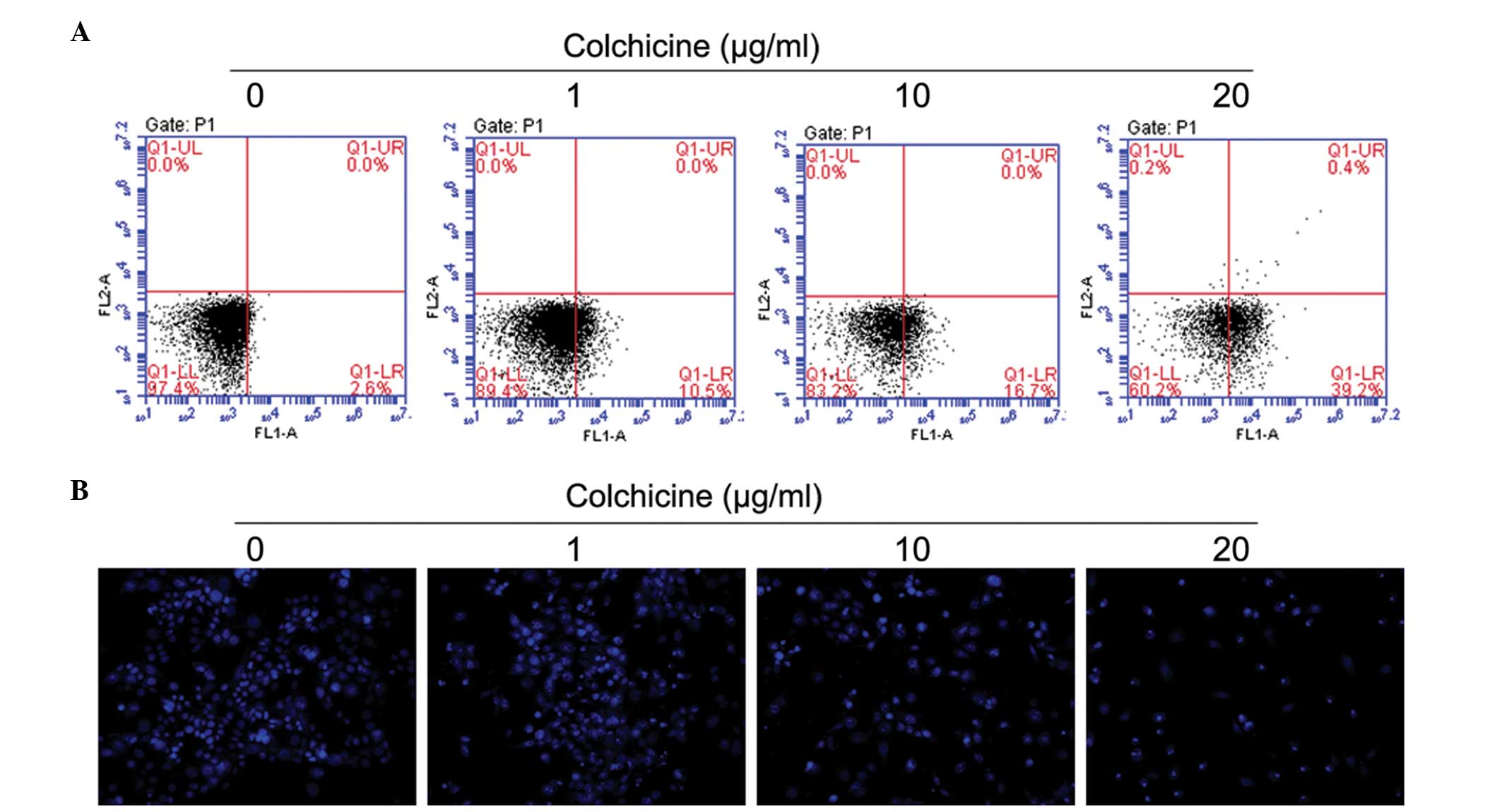
Colchicine inhibited cell proliferation and induced
cellular death in colon cancer cells; however, the underlying
mechanisms have not been investigated in detail. To establish
whether this treatment could also induce apoptosis in HT-29 cells,
flow cytometry was used to assess the percentage of apoptotic cells
induced by colchicine treatment. Similar to the results of the cell
viability assay, colchicine concentrations of >20 μg/ml
significantly increased cell apoptosis compared with the respective
control cells (Fig. 2A), and the
increase was dose- and time-dependent. Treatment of colchicine at
doses of 10 and 20 μg/ml for 24 h significantly increased
the number of early apoptotic cells (Fig. 2A: Q1-LR) in a dose-dependent manner
compared with control cells. The significant induction of apoptosis
indicated the anticancer effect of colchicine against HT-29 cells.
The colchicine-induced apoptosis in the HT-29 cells was
morphologically identified using fluorescence staining with Hoechst
33258 (Fig. 2B); chromosomal
condensation and nuclear fragmentation were observed in the
colchicine-treated cells.
Colchicine induces colon cancer cell
apoptosis via the mitochondrial pathway
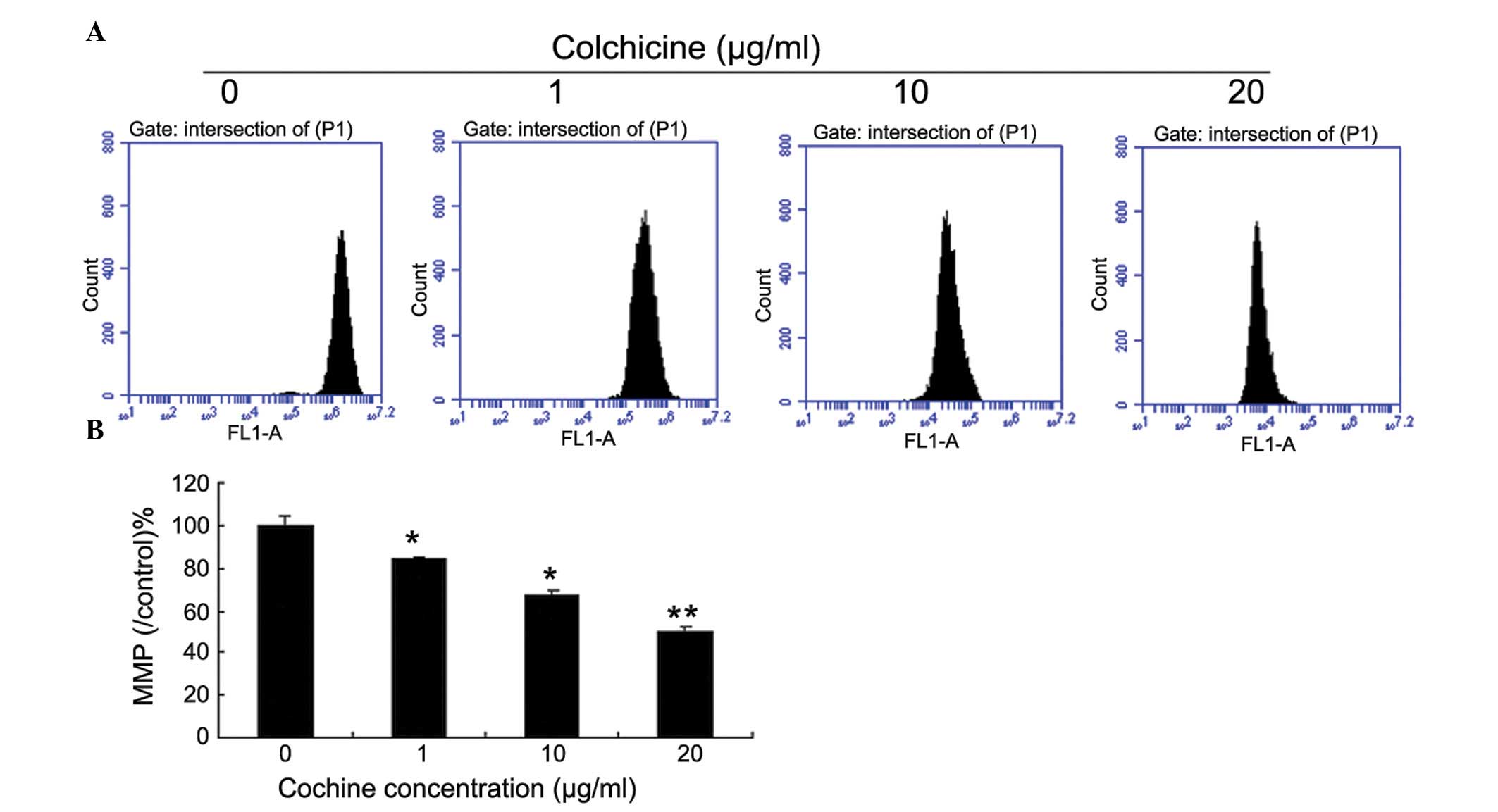
During the apoptotic process, mitochondrial membrane
pores open and Δψm is disrupted. As shown in Fig. 3, the JC-1 staining data
demonstrates that colchicine-treated cells exhibited a significant
decrease in the Δψm in a dose-dependent manner, when compared with
the control cells.
Colchicine increases ROS production in
HT-29 cells
Mitochondria are critical in the initiation and
progression of cell death processes and, therefore, may serve as
targets for numerous chemotherapeutic agents. To determine whether
the mitochondria are involved in colchicine-induced cell death,
mitochondria were isolated from HT-29 cells and treated with 0, 1,
10 or 20 μg colchicine for 12 h. Staining with Amplex Red in
combination with horse-radish peroxidase followed by flow
cytometric analysis of the fluorescence showed that colchicine
increased ROS production of HT-29 cells in a dose-dependent manner
(Fig. 4).
Colchicine promotes colon cancer cell
apoptosis vua activation of the p38 pathway
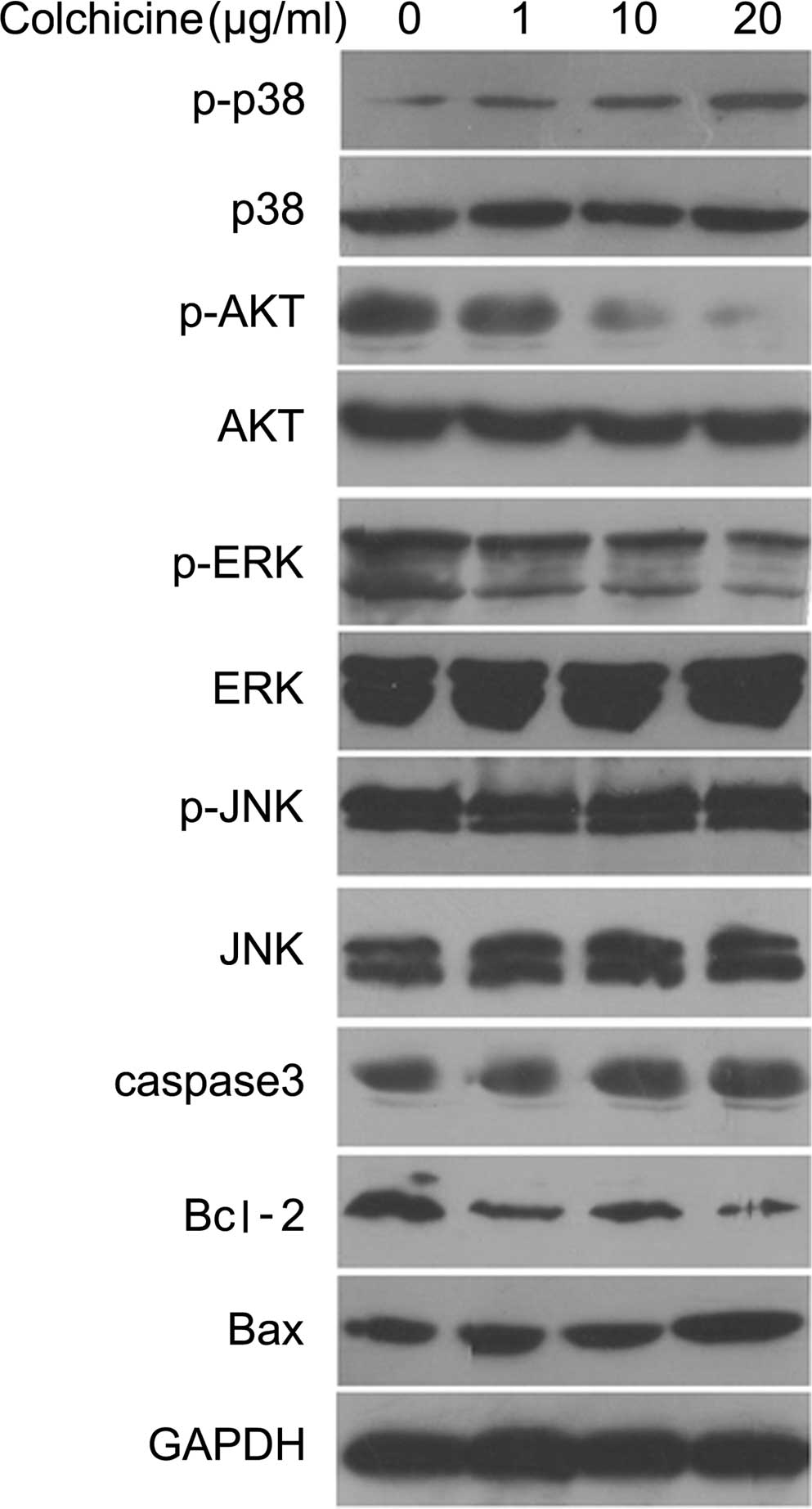
The expression of apoptosis-associated proteins and
the phosphorylation of kinases was detected by western blot and
hybridization to clarify the mechanism of HT-29 cell apoptosis
induced by colchicine treatment. As shown in Fig. 5, treatment with colchicine (0, 1,
10 and 20 μg/ml) for 12 h increased p38 phosphorylation
dose-dependently and decreased AKT phosphorylation. However,
colchicine treatment did not significantly affect phosphorylation
of the JNK and ERK proteins. Furthermore, treatment with colchicine
for 12 h evidently increased the protein expression of caspase-3
and Bax, and decreased Bcl-2 expression in a dose-dependent manner
when compared with the control group (Fig. 5).
Discussion
In recent years, numerous reports have indicated
that colchicine exhibits potent antitumor activity against various
tumor cell lines, such as liver, colon, gastric and lung cancer
cells (14–19). In the present study, the inhibitory
effect of colchicine on HT-29 colon cancer cells was investigated
and the possible molecular mechanism involved was elucidated. The
results revealed that colchicine inhibited HT-29 cell proliferation
in a time- and dose-dependent manner. Flow cytometric analysis with
Annexin V/PI staining demonstrated that colchicine treatment with
doses ranging between 1 and 20 μg/ml induced apoptosis and
increased DNA fragmentation of HT-29 cells in a dose-dependent
manner. The data regarding cell viability, cell apoptosis and DNA
fragmentation obtained in the present study, indicates that
colchicine penetrates HT-29 cells and destroys mitochondria
membrane integrity, which consequently results in cell
apoptosis.
Mitochondria are crucial in the complex process of
apoptosis (20). During this
process, mitochondrial membrane pores are opened, resulting in the
loss of Δψm (21). This loss of
Δψm causes an increase in the permeability of the mitochondrial
membrane, followed by the release of proapoptotic molecules, such
as cytochrome c. The release of cytochrome c from the
mitochondrial membrane interacts with adenosine triphosphate (ATP),
Apaf-1 and caspase-9, and subsequently activates caspase-3, which
elicits caspase-dependent apoptotic cell death (22). The current study demonstrated that
colchicine increases Δψm.
Activation of mitogen activated protein kinase
(MAPK) is associated with cell cycle arrest and apoptosis
induction. There are three known parallel pathways of the MAPK
family: ERK, JNK and p38 (23). In
the present study, the phosphorylation levels of ERK, JNK and p38
were detected by western blotting, however, only p38
phosphorylation was observed to increase as a result of colchicine
treatment. The present study demonstrated the effect of colchicine
on JNK signaling in HT-29 cells. JNK promotes Bax translocation to
the mitochondria and the release of cytochrome c from the
mitochondria. The released cytochrome c, in combination with
Apaf-1, ATP and caspase-9, forms the apoptosomes and ultimately
leads to activation of caspase-3 and death of the target cell
(24). AKT is a master regulator
involved in transcriptional regulation of the antiapoptotic
protein, Bcl-2, which is critical in preventing cell death
(25). In the present study,
phosphorylation of AKT was decreased by colchicine treatment.
In conclusion, the present study demonstrates that
colchicine may inhibit HT-29 colon cancer cell proliferation and
induce HT-29 cell apoptosis through the mitochondrial pathway via
activation of the p38 pathway.
Acknowledgments
The current study was supported by Hongkou District
key project of Shanghai City Health Bureau. The authors would like
to thank Biomedworld for their assistance with editing the
manuscript.
References
|
1
|
Giovannucci E: Modifiable risk factors for
colon cancer. Gastroenterol Clin North Am. 31:925–943. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Couzin J: Cancer T cells a boon for colon
cancer prognosis. Science. 313:1868–1869. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
François Y and Vignal J: Cancer of the
colon. Etiology, physiopathology, diagnosis, development and
prognosis, principles of the surgical treatment. Rev Prat.
41:1221–1225. 1991.
|
|
4
|
Chiu BC, Ji BT, Dai Q, Gridley G,
McLaughlin JK, Gao YT, Fraumeni JF Jr and Chow WH: Dietary factors
and risk of colon cancer in Shanghai, China. Cancer Epidemiol,
Biomarkers Prev. 12:201–208. 2003.
|
|
5
|
Barrier A, Boelle PY, Roser F, Gregg J,
Tse C, Brault D, Lacaine F, Houry S, Huguier M, Franc B, et al:
Stage II colon cancer prognosis prediction by tumor gene expression
profiling. J Clin Oncol. 24:4685–4691. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Jee SH, Moon SM, Shin US, Yang HM and
Hwang DY: Effectiveness of adjuvant chemotherapy with
5-FU/leucovorin and prognosis in stage II colon cancer. J Korean
Soc Coloproctol. 27:322–328. 2011. View Article : Google Scholar
|
|
7
|
Dotan E and Cohen SJ: Challenges in the
management of stage II colon cancer. Semin Oncol. 38:511–520. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Risinger AL and Mooberry SL:
Taccalonolides: Novel micro-tubule stabilizers with clinical
potential. Cancer Lett. 291:14–19. 2010. View Article : Google Scholar :
|
|
9
|
Sivakumar G: Colchicine semisynthetics:
Chemotherapeutics for cancer? Curr Med Chem. 20:892–898. 2013.
|
|
10
|
Risinger AL, Giles FJ and Mooberry SL:
Microtubule dynamics as a target in oncology. Cancer Treat Rev.
35:255–261. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Jordan MA and Wilson L: Microtubules as a
target for anticancer drugs. Nat Rev Cancer. 4:253–265. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ravelli RB, Gigant B, Curmi PA, Jourdain
I, Lachkar S, Sobel A and Knossow M: Insight into tubulin
regulation from a complex with colchicine and a stathmin-like
domain. Nature. 428:198–202. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bhattacharyya B, Panda D, Gupta S and
Banerjee M: Anti-mitotic activity of colchicine and the structural
basis for its interaction with tubulin. Med Res Rev. 28:155–183.
2008. View Article : Google Scholar
|
|
14
|
Chopra A, Anderson A and Giardina C: Novel
piperazine-based compounds inhibit microtubule dynamics and
sensitize colon cancer cells to tumor necrosis factor-induced
apoptosis. J Biol Chem. 289:2978–2991. 2014. View Article : Google Scholar :
|
|
15
|
Acharya BR, Chatterjee A, Ganguli A,
Bhattacharya S and Chakrabarti G: Thymoquinone inhibits microtubule
polymerization by tubulin binding and causes mitotic arrest
following apoptosis in A549 cells. Biochimie. 97:78–91. 2014.
View Article : Google Scholar
|
|
16
|
Rai A, Gupta TK, Kini S, Kunwar A, Surolia
A and Panda D: CXI-benzo-84 reversibly binds to tubulin at
colchicine site and induces apoptosis in cancer cells. Biochem
Pharmacol. 86:378–391. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Magalhães HI, Wilke DV, Bezerra DP,
Cavalcanti BC, Rotta R, de Lima DP, Beatriz A, Moraes MO,
Diniz-Filho J and Pessoa C:
(4-Methoxyphenyl)(3,4,5-trimethoxyphenyl)methanone inhibits tubulin
polymerization, induces G2/M arrest and triggers apoptosis in human
leukemia HL-60 cells. Toxicol Appl Pharmacol. 272:117–126. 2013.
View Article : Google Scholar
|
|
18
|
Chiang NJ, Lin CI, Liou JP, Kuo CC, Chang
CY, Chen LT and Chang JY: A novel synthetic microtubule inhibitor,
MPT0B214 exhibits antitumor activity in human tumor cells through
mitochondria-dependent intrinsic pathway. PLoS One. 8:e589532013.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wu J, Yi W, Jin L, Hu D and Song B:
Antiproliferative and cell apoptosis-inducing activities of
compounds from Buddleja davidii in Mgc-803 cells. Cell Div.
7:202012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang X: The expanding role of mitochondria
in apoptosis. Genes Dev. 15:2922–2933. 2001.PubMed/NCBI
|
|
21
|
Zamzami N, Marchetti P, Castedo M, Hirsch
T, Susin SA, Masse B and Kroemer G: Inhibitors of permeability
transition interfere with the disruption of the mitochondrial
trans-membrane potential during apoptosis. FEBS Lett. 384:53–57.
1996. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yan SL, Huang CY, Wu ST and Yin MC:
Oleanolic acid and ursolic acid induce apoptosis in four human
liver cancer cell lines. Toxicol In Vitro. 24:842–848. 2010.
View Article : Google Scholar
|
|
23
|
Roux PP and Blenis J: ERK and p38
MAPK-activated protein kinases: A family of protein kinases with
diverse biological functions. Microbiol Mol Biol Rev. 68:320–344.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Tsuruta F, Sunayama J, Mori Y, Hattori S,
Shimizu S, Tsujimoto Y, Yoshioka K, Masuyama N and Gotoh Y: JNK
promotes Bax translocation to mitochondria through phosphorylation
of 14-3-3 proteins. EMBO J. 23:1889–1899. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Pugazhenthi S, Nesterova A, Sable C,
Heidenreich KA, Boxer LM, Heasley LE and Reusch JE: Akt/protein
kinase B up-regulates Bcl-2 expression through cAMP-response
element-binding protein. J Biol Chem. 275:10761–10766. 2000.
View Article : Google Scholar
|