Introduction
Podocytes are critical in maintaining the integrity
of the glomerular filtration barrier. Acquired proteinuric
glomerulopathies, including minimal change disease and focal
segmental glomerulosclerosis, commonly exhibit abnormalities in
podocytic phenotypes, including effacement and apoptosis (1,2).
Glucocorticoids, which are the primary therapy for the treatment of
proteinuric glomerulopathies, exert profound effects through their
interaction with specific glucorticoid receptors (GRs) (3). GRs are abundantly expressed in human
and mouse podocytes (4,5), and exist in different isoforms as a
result of alternative splicing. GRα is the functionally active
isoform located in the cytoplasm. Glucocorticoids are reported to
induce recovery in podocyte injuries predominantly via two
GRα-mediated mechanisms. Genetic regulation occurs following the
binding of glucocorticoids at low doses (0.5 mg/kg/day), resulting
in the activation and translocation of GRα into the nucleus.
Subsequently, activated GRα binds at the glucocorticoid response
elements within the genome to mediate the direct regulation of
target genes leading to anti-inflammatory and immunosuppressive
effects (5). The alternative
mechanism involves the direct effect of glucocorticoids on
podocyte-expressed GRα (4), which
reduces podocytic apoptosis and promotes actin aggregation
(6). Therefore, GRα regulation is
a crucial determinant of steroid efficacy.
Our previous study demonstrated a reduced expression
of GRα in podocytes from patients with steroid-resistant idiopathic
nephrotic syndrome compared with steroid-sensitive patients
(7). Therefore, upregulation of
the GRα may be a promising target for the improvement of steroid
sensitivity. miRNAs are post-transcriptional regulators of gene
expression, which bind to complementary sequences in the target
mRNA to induce gene repression (8,9).
Previous data indicates that GRα-specific miRNAs represent
potential therapeutic targets in T-cell acute lymphoblastic
leukemia and multiple myeloma (10,11).
However, the regulation of GRα by miRNAs in podocytes remains to be
determined. In the present study, the miRNA expression profile in
normal murine podocytes and injured podocytes induced by puromycin
aminonucleoside (PAN) was compared, and demonstrated the existence
of an association between miR-30a and the expression of GR, based
on bioinformatic analysis. Subsequently, the present study observed
that miR-30a downregulated GRα in injured podocytes. Furthermore,
inhibition of miR-30a reduced podocytic apoptosis by upregulating
GRα. These findings indicated that inhibition of miR-30a may reduce
podocytic apoptosis via a GRα-mediated mechanism.
Materials and methods
miRNA microarray
miRNA microarray analysis was performed using miRNA
microarrays (RiboBio Co., Ltd., Guangzhou, China), according to the
manufacturer's instructions. Briefly, 600 ng total RNA was isolated
from differentiated murine podocytes and injured podocytes induced
by PAN, and was dephosphorylated with a calf intestine alkaline
phosphatase treatment for 30 min at 37°C prior to labeling. The
samples were diluted with dimethylsulfoxide, denatured for 10 min
at 100°C and were subsequently labeled with pCp-Cy3 (Biorbyt,
Cambridge, UK) in T4 RNA ligation buffer (BestBio, Shanghai,
China). The labeled RNA was hybridized using 2X hybridization
buffer (Agilent Technologies, Palo Alto, CA, USA), washed with 6X
SSC (Solarbio Science & Technology Co., Ltd., Beijing,
China)/0.005% Triton X-102 (DOW, Midland, MI, USA), stained with
horseradish peroxidase-streptavidin (RiboBio Co., Ltd.) and scanned
with an Agilent 2200 Bioanalyzer (Agilent Technologies). Microarray
data analysis was performed using an Illumina HiSeq™2500 (Illumina
Inc., San Diego, CA, USA).
Prediction of murine GR targeting
miRNAs
The following target prediction algorithms were used
to predict murine GR targeting miRNAs: Pictar (http://pictar.mdc-berlin.de/), miRanda (http://www.microrna.org/) and TargetScan/TargetScanS
(http://www.targetscan.org/). The highest
threshold value was used for the prediction using miRanda.
Podocytes culture
The conditionally immortalized mouse MPC5 podocyte
cell line was cultured, as previously described (6). Briefly, undifferentiated podocytes
were grown at 33°C with a relative humidity of 100% and 5%
CO2-containing atmosphere in RPMI-1640 medium (Hyclone,
Logan, UT, USA), supplemented with 10% fetal bovine serum (FBS;
Gibco Life Technologies, Grand Island, NY, USA), 100 U/ml
penicillin, 100 µg/ml streptomycin (Proteintech Group, Inc.,
Chicago, IL, USA) and 10 U/ml mouse interferon-γ (IFN-γ; Sigma
Chemical Co., St. Louis, MO, USA). Podocyte differentiation was
induced by culturing at 37°C for 10–14 days in the presence of rat
tail collagen type I (Sigma Chemical Co.) without IFN-γ.
Podocyte treatment
Podocyte injury was induced in vitro with PAN
(Sigma Chemical Co.) using the following procedure: Differentiated
podocytes (day 12) were incubated in RPMI-1640 medium, supplemented
with charcoal-stripped FBS for 24 h. Drug treatments were performed
24 h following transfection. The podocytes were exposed to PAN at a
concentration of 45 ng/ml and were incubated for 48 h. Podocyte
recovery was induced using dexamethasone (Dex; Sigma Chemcial Co.).
The cells were pretreated for 1 h with the GR agonist, Dex (1
µM) prior to PAN treatment. Correspondingly, the cells were
pretreated with the GR antagonist, RU486 (20 µM; Sigma
Chemical Co.), for 30 min prior Dex treatment.
Transfection
Using the methods described by the relevant
manufacturers, the podocytes (day 12) were transiently transfected
with 50 nM synthetic miR-30a mimics, 100 nM inhibitor and the
corresponding scrambled control oligonucleotides (RiboBio Co.,
Ltd.). At 48 h post-transfection, the cells were harvested and
prepared for analysis.
Total RNA isolation and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
On the 12th day of podocyte differentiation (day
12), the total RNA was isolated using TRIzol reagent (Invitrogen
Life Technologies, Carlsbad, CA, USA), according to the
manufacturer's instructions. The RNA concentration and purity was
assessed by measuring the absorbance at 260 and 280 nm using a
Nanodrop spectrophotometer 2000 (Thermo Fisher Scientific, Waltham,
MA, USA).
The primers for assay of miR-30a were purchased from
ABI (Applied Biosystems, Foster City, CA, USA). RT reactions,
containing the total RNA (5 µl), 50 nM stem-loop RT primer
(3 µl), 10X RT buffer (1.5 µl), 100 mM dNTPs (0.15
µl), multiScribe reverse transcriptase (1 µl) and
RNase inhibitor (0.19 µl), were performed using a GeneAmp
PCR system 9700 (Applied Biosystems) for 30 min at 16°C, 30 min at
42°C and 5 min at 85°C. PCR amplification was performed using a
standard TaqMan PCR kit (Applied Biosystems) with the RT product
(1.33 µl), 2X TaqMan Universal PCR Master mix (14.5
µl), 20X TaqMan MicroRNA assay (1.5 µl). The PCR
amplification conditions were as follows: 95°C for 10 min, followed
by 40 cycles of 95°C for 15 sec and 60°C for 1 min. The
intra-experimental variation was assessed based on the average of
the mean ΔΔCt values following normalization against U6. All
samples were assessed in triplicate.
The gene expression levels were analyzed by RT-qPCR
on a LightCycler (Roche, Mannheim, Germany) using the following
primers (Takara Bio, Inc., Otsu, Japan): GRα, sense:
5′-AAAGAGCTAGGAAAAGCCATTGTC-3′ and antisense:
5′-TCAGCTAACATCTCTGGGAATTCA-3′; GAPDH, sense:
5′-GAAGGTGAAGGTCGGAGT-3′ and antisense: 5′-GAAGATGGTGATGGGATTTC-3′.
The total RNA (2 µl) was used for RT using the
SYBR® PrimerScriptTM RT-PCR kit (Takara Bio,
Inc.), according to the manufacturer's instructions. The resulting
data were analyzed using the comparative ΔΔCt method for
relative gene expression quantification against GAPDH (12).
Analysis of the expression of GRα by
western blotting
The protein concentration of the cell lysates (100
µg) was determined using a bicinchoninic acid protein assay
kit (Pierce Biotechnology, Rockford, IL, USA). MPC5 cells grown in
six-pore plates (Costar Corning, Corning, NY, USA) were washed
twice with PBS, then treated with RIPA cell lysis solution
(Solarbio Science & Technology Co., Ltd.) on ice for 30 min,
then centrifuged at 12,000 × g for 30 min at 4°C to attain the
supernatant. The protein samples were heated to 100°C for 5 min
prior to 20 µl aliquots being separated by SDS-PAGE (Beijing
Solarbio Science & Technology Co., Ltd.) and transferred onto
polyvinylidene membranes (Beijing Solarbio Science & Technology
Co., Ltd.). The membranes were incubated with a polyclonal
rabbit-anti-mouse GRα primary antibody (cat. no. sc-37642; 1:250;
Santa Cruz Biotechnology, Inc., CA, USA) in 10 ml dilution buffer
(1X Tris-buffered saline, 0.1% Tween-20 and 5% BSA), with agitation
overnight at 4°C. The membranes were subsequently incubated with
the appropriate horseradish peroxidase-conjugated anti-rabbit
immunoglobulin G (cat. no. zf-0311; 1:2,000; Zhongshan
Biotechnology Co., Ltd., Beijing, China) in 10 ml dilution buffer
with agitation for 1 h at 25°C. The expression of GRα was
normalized against GAPDH, as an internal control. The bands were
detected using an enhanced chemiluminescence kit (Amersham,
Pittsburgh, PA, USA) and the protein expression was quantified
using Labworks™ Image Acquisition and Analysis software version 4.6
(UVP LLC, Upland, CA, USA).
Flow cytometric analysis of podocyte
apoptosis
The apoptosis rate in the transfected podocytes
exposed to Dex and PAN was assessed using a fluorescein
isothiocyanate (FITC)-conjugated annexin V apoptosis detection kit
(Molecular Probes, Eugene, Oregon, USA). Briefly, the podocytes
were gently trypsinized and pelleted by centrifugation at 900 × g
for 5 min at room temperature. The pellet was washed with
phosphate-buffered saline (PBS) and resuspended in binding buffer.
The cells were subsequently stained with FITC-annexin V and
propidium iodide (PI) for 5 min on ice in the dark. The stained
cell data was acquired on a FACScan flow cytometer (Becton
Dickinson, San Jose, CA, USA) and analyzed using CellQuest Pro
software 5.1 (BD Biosciences, San Jose, CA, USA). At least
1×106/ml cells were assessed per sample and the
apoptotic cells were defined as annexin V positive and PI
negative.
Podocyte viability assays
Podocytes transfected with the miR-30a mimics and
inhibitor were pretreated with Dex followed by PAN. The viability
was determined by fluorescence microscopy (BX51TF; Olympus, Tokyo,
Japan) following staining with an EdU incorporation kit (RiboBio
Co. Ltd.), according to the manufacturer's instructions.
Cloning of pmiR-RB-REPORT™ construct
Prediction of the 3′-untranslated region (UTR) and
coding sequence (CDS) was performed with miRanda v1.0b and RNA22,
respectively. The GRα 3′-UTR (1-2021 bp) fragment, bearing the
predicted miR-30a binding site, was isolated from murine cDNA by
PCR. The PCR product was isolated by agarose gel electrophoresis,
purified and cloned into the pmiR-RB-REPORT™ vector (RiboBio Co.,
Ltd.). The resulting construct was designated 3′-UTR (1-2021 bp)
WT. The predicted miR-30a binding site (GTTTGTG) was mutated
(CTATCTC), inserted into another vector and designated 3′-UTR
(1-2021 bp) MT. The GRα 3′-UTR (2020-3953 bp) and CDS fragments
were cloned into another two vectors, and were designated 3′-UTR
(2020-3953 bp) WT and CDS WT, respectively. The primer sequences
(RiboBio Co., Ltd.) were as follows: 3′-UTR (1-2021 bp) WT and
3′-UTR (1-2021 bp) MT, sense:
5′-CCGCTCGAGCTGCCTTACTAAGAAAGGCTGCCTTAAAG-3′ and antisense:
5′-GAATGCGGCCGCAGCCTCTGTTTCTGTTAACC-3′; 3′-UTR (2020-3953 bp) WT,
sense: 5′-CCGCTCGAGACAAGAGTCCATTTCCAAGTAAG-3′ and antisense:
5′-GAATGCGGCCGCAGTCACATAATGAAAAAGCAG-3′; CDS WT, sense:
5′-CCGCTCGAGATGGACTCCAAAGAATCCTTAGC-3′ and antisense:
5′-GAATGCGGCCGCTCATTTCTGATGAAACAGAAGC-3′. The plasmid constructs
were confirmed by automated DNA sequencing.
Luciferase assays
NIH3T3 murine fibroblast cells were seeded into a
96-well plate (4,000 cells/well) and were co-transfected with a
reporter construct (100 ng/ml) and synthetic miR-30a (50 nM),
according to the manufacturer's instructions (RiboBio Co. Ltd.). At
24 h post-transfection, the cells were harvested in 100 µl
Passive Lysis Buffer (Promega, Madison, WI, USA) and the luciferase
activity was measured in the lysates (35 µl) using a
luciferase reporter assay kit (Promega).
Statistical analysis
SPSS version 18.0 (IBM SPSS, Chicago, IL, USA) was
used for statistical analyses. For microarray analysis, the raw
expression signals were log transformed, normalized and filtered,
according to the median corrected signal of all the miRNAs with an
intensity >100. The miRNAs displaying differential expression
between differentiated murine podocytes and injured podocytes
induced by PAN were detected using a two-sample t-test. The data
are presented as the mean ± standard deviation and interpreted by
analysis of variance with Tukey's test. P<0.05 was considered to
indicate a statistically significant difference.
Results
Identification of differentially
expressed miRNA in differentiated murine podocytes and injured
podocytes induced by PAN
Of the 1,908 murine miRNAs represented on the
microarrays, 187 were expressed in differentiated podocytes and 172
in injured podocytes induced by PAN. A total of 19 miRNAs were
identified to be significantly upregulated and 35 were
significantly downregulated (Table
I). Following the application of a fold change threshold >2
(false discovery rate <0.01), 10 miRNAs, including miR-30a,
miR-27a-3p, miR-30d, miR-100, miR181c, miR-5099, miR-3535,
miR-140-3p, miR-148-3p and miR-103-3p were significantly
upregulated.
 | Table IIdentification of differentially
expressed miRNA in differentiated murine podocytes and injured
podocytes induced by PAN. |
Table I
Identification of differentially
expressed miRNA in differentiated murine podocytes and injured
podocytes induced by PAN.
| miRNA ID | Differentiated
podocytes | Podocytes induced
by PAN | P-value |
|---|
| mmu-miR-30a-5p | 8928.94 | 33333.19 | <0.01 |
| mmu-miR-27a-3p | 5597.11 | 18809.07 | <0.01 |
| mmu-miR-30d-5p | 4084.54 | 10617.27 | <0.01 |
| mmu-miR-100-5p | 576.49 | 2146.32 | <0.01 |
| mmu-miR-140-3p | 472.84 | 1605.43 | <0.01 |
| mmu-miR-103-3p | 343.27 | 3171.62 | <0.01 |
|
mmu-miR-148a-3p | 228.49 | 1193.20 | <0.01 |
|
mmu-miR-181c-5p | 23.17 | 2069.71 | <0.01 |
| mmu-miR-5099 | 0.00 | 28.84 | <0.01 |
| mmu-miR-3535 | 0.00 | 12.11 | <0.01 |
| mmu-miR-10b-5p | 9607.71 | 23206.82 | <0.05 |
| mmu-miR-10a-5p | 8993.11 | 22036.14 | <0.05 |
|
mmu-miR-106b-5p | 359.89 | 881.80 | <0.05 |
| mmu-miR-28a-3p | 89.02 | 252.70 | <0.05 |
| mmu-miR-542-3p | 31.25 | 98.28 | <0.05 |
|
mmu-miR-195a-5p | 3.81 | 22.69 | <0.05 |
| mmu-miR-429-3p | 3.35 | 20.99 | <0.05 |
| mmu-miR-6236 | 0.00 | 7.00 | <0.05 |
| mmu-miR-26a-5p | 5669.51 | 376.40 | <0.01 |
| mmu-miR-107-3p | 1315.63 | 81.05 | <0.01 |
| mmu-miR-378d | 639.75 | 0.00 | <0.01 |
| mmu-miR-128-3p | 574.66 | 72.69 | <0.01 |
|
mmu-miR-181b-5p | 478.94 | 215.67 | <0.01 |
| mmu-miR-425-5p | 458.21 | 126.43 | <0.01 |
| mmu-miR-22-3p | 408.67 | 95.38 | <0.01 |
| mmu-miR-191-5p | 403.18 | 144.69 | <0.01 |
|
mmu-miR-3074-2-3p | 323.61 | 142.13 | <0.01 |
| mmu-miR-29b-3p | 172.70 | 77.29 | <0.01 |
| mmu-miR-423-3p | 124.54 | 58.70 | <0.01 |
| mmu-miR-615-3p | 115.54 | 39.59 | <0.01 |
|
mmu-miR-16-1-3p | 87.19 | 11.77 | <0.01 |
| mmu-miR-25-5p | 74.39 | 7.85 | <0.01 |
| mmu-miR-362-5p | 64.94 | 2.39 | <0.01 |
| mmu-miR-671-5p | 51.52 | 5.29 | <0.01 |
|
mmu-miR-450a-5p | 33.69 | 16.72 | <0.01 |
| mmu-miR-296-3p | 29.57 | 13.31 | <0.01 |
| mmu-miR-24-3p | 26.98 | 0.00 | <0.01 |
| mmu-miR-340-3p | 23.78 | 5.12 | <0.01 |
|
mmu-miR-24-2-5p | 17.83 | 6.48 | <0.01 |
|
mmu-miR-30c-2-3p | 14.94 | 5.97 | <0.01 |
| mmu-miR-152-5p | 14.63 | 3.92 | <0.01 |
| mmu-let-7d-3p | 14.48 | 5.46 | <0.01 |
| mmu-miR-350-5p | 14.33 | 3.58 | <0.01 |
|
mmu-miR-126a-3p | 10.21 | 3.58 | <0.01 |
|
mmu-miR-199b-3p | 10.21 | 0.00 | <0.01 |
|
mmu-miR-466e-3p | 10.06 | 0.00 | <0.01 |
| mmu-miR-132-5p | 6.25 | 0.00 | <0.01 |
|
mmu-miR-301a-3p | 14.18 | 7.00 | <0.05 |
|
mmu-miR-1306-3p | 7.77 | 2.05 | <0.05 |
| mmu-miR-182-3p | 6.25 | 2.05 | <0.05 |
|
mmu-miR-181b-1-3p | 6.10 | 0.00 | <0.05 |
|
mmu-miR-3102-3p.2-3p | 5.79 | 0.00 | <0.05 |
|
mmu-miR-1964-3p | 5.03 | 0.00 | <0.05 |
GR is a putative target of miR-30a in
murine podocytes
To identify miRNAs with the potential to modify GR
activity by binding to the 3′-UTR in the 10 significantly
upregulated miRNAs, an in silico search was performed using
Pictar, miRanda and TargetScan/TargetScanS databases. Based on the
maximum likelihood of binding conservation and accessibility, all
three databases predicted that GR was a putative target of miR-30a
in the mouse.
miR-30a negatively regulates GRα in
normal and injured podocytes induced by PAN
Based on the results of the bioinformatics
investigations, the expression levels of GRα and miR-30a were
analyzed in undifferentiated and differentiated podocytes by
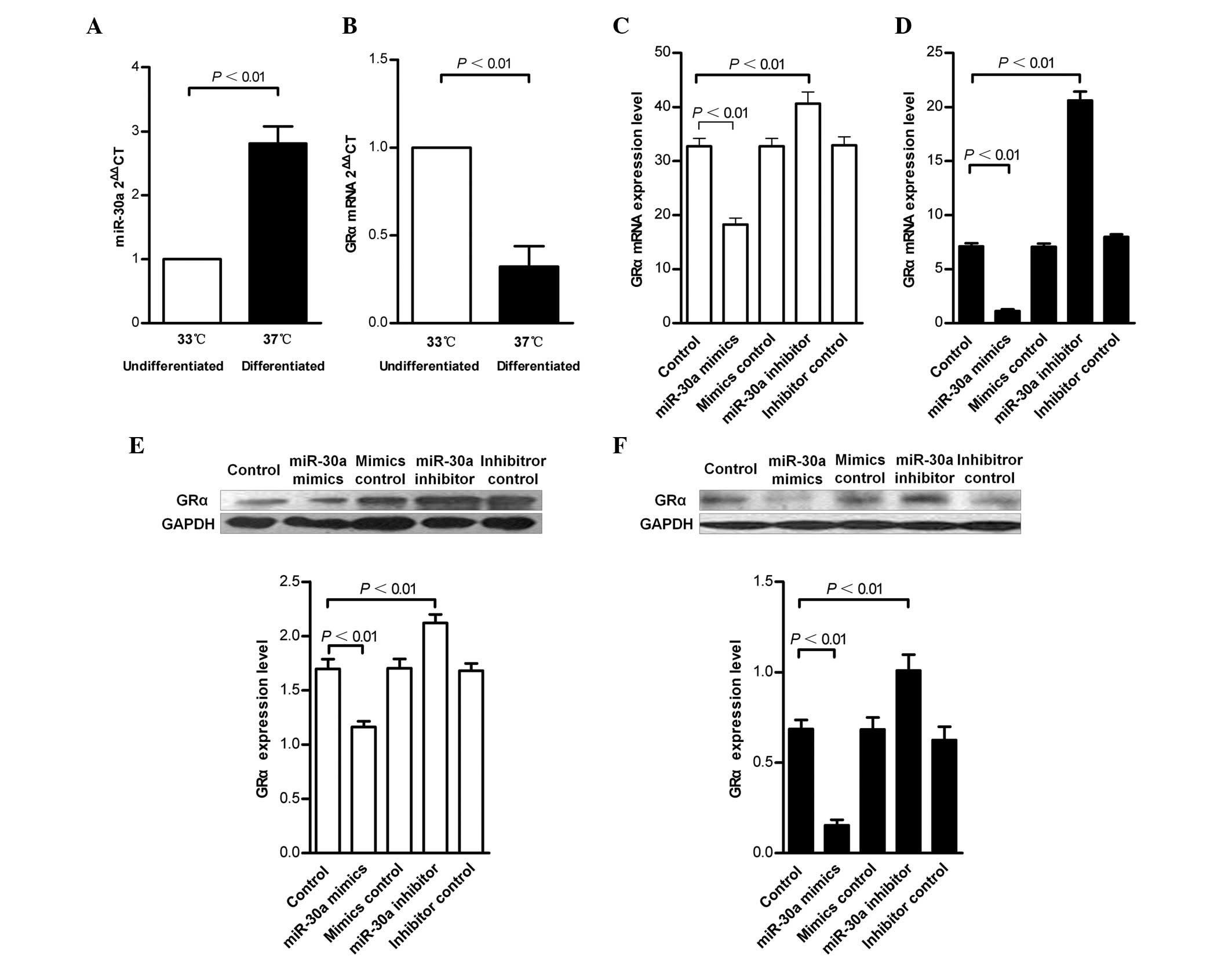
RT-qPCR. While the expression of miR-30a increased, the expression
of GRα decreased during the process of podocytic maturation
(Fig. 1A and B). Subsequently, it
was revealed that transfection of undifferentiated and
differentiated podocytes with the miR-30a mimics caused a reduction
in the mRNA and protein expression levels of GRα (25–60 and 25–40%,
respectively). By contrast, treatment with the miR-30a inhibitor
resulted in increased mRNA and protein expression levels of GRα
(25–35 and 10–25%, respectively; Fig.
2C–F).
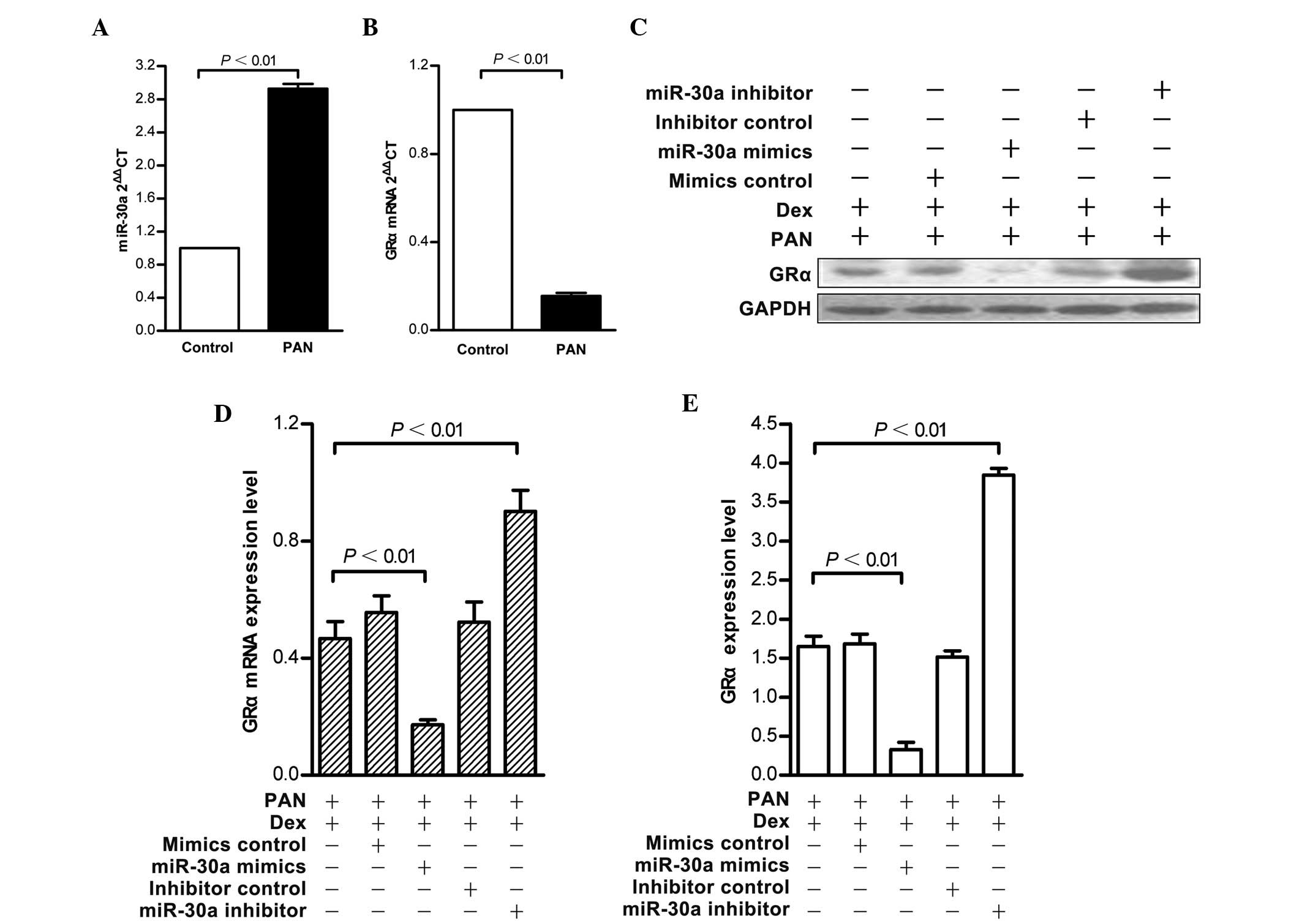
An in vitro model of podocyte injury induced
by PAN was established to further confirm the effect of miR-30a on
GRα. Compared with the normal group, the mRNA expression of GRα
decreased in injured podocytes and the expression of miR-30a
increased, which confirmed the microarray results (Fig. 2A and B). An identical pattern was
observed in response to PAN-injury following miR-30a mimic or
inhibitor transfection of the podocytes. Transfection with the
miR-30a mimics caused a 25% reduction in the mRNA and protein
expression levels of GRα, while miR-30a inhibitor treatment
resulted in a 25% increase in GRα RNA, and a 10% increase in GRα
protein (Fig. 2C–E).
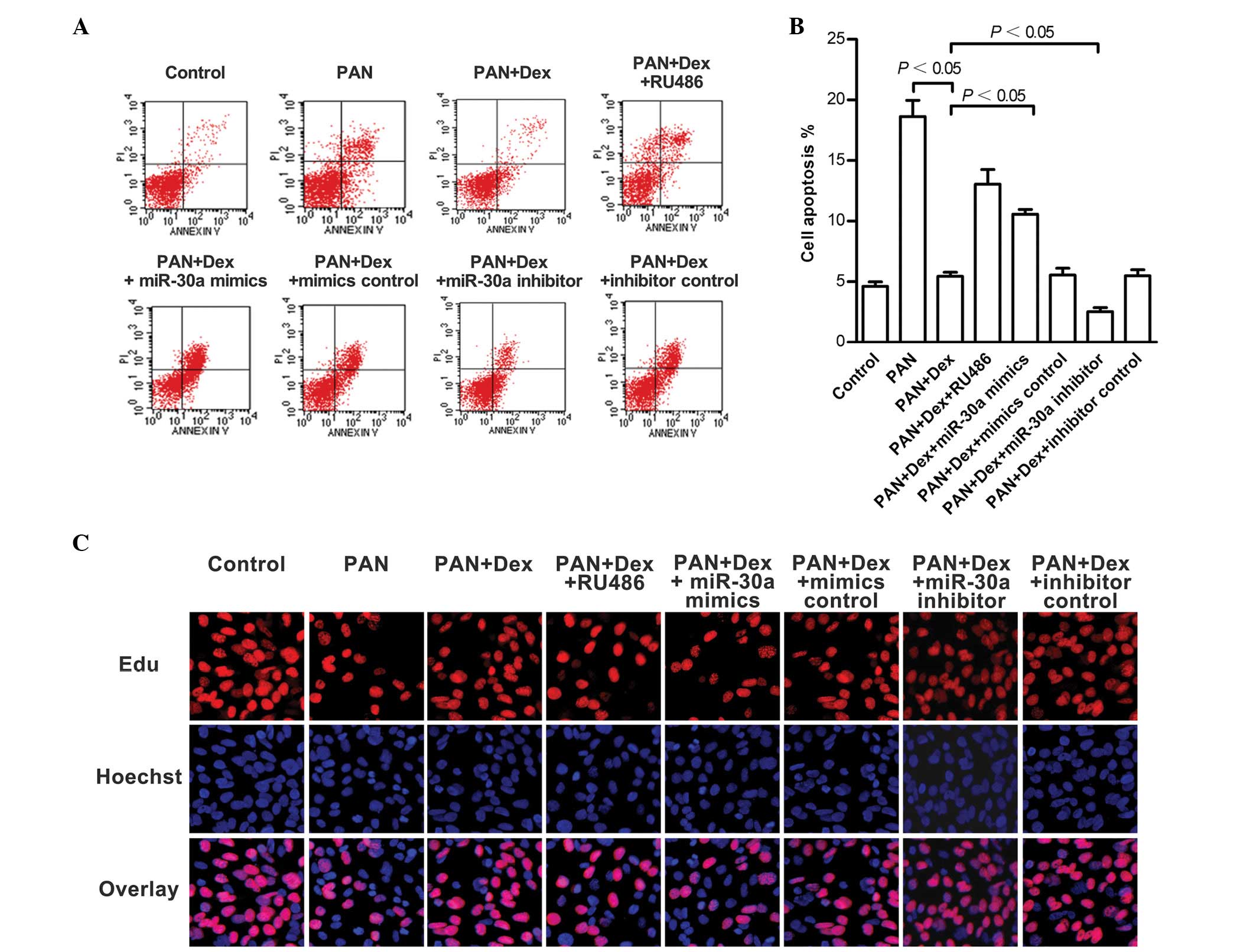
Inhibition of miR-30a prevents
PAN-induced podocyte apoptosis
Apoptosis is the predominant characteristic of
PAN-induced podocytic injury. Flow cytometric analysis revealed
that podocyte apoptosis increased following PAN treatment and was
ameliorated by Dex (P<0.05). However, this amelioration was
inhibited by the GR inhibitor, RU486 (P>0.05). Transfection of
PAN and Dex pretreated podocytes with miR-30a mimics or the miR-30a
inhibitor resulted in increased and decreased levels of apoptosis,
respectively (P<0.05; Fig. 3A and
B).
Furthermore, EdU incorporation analysis of the
injured podocytes demonstrated that the proliferation rate was
increased by Dex and that this increase was inhibited by RU486
(P<0.05). Transfection of PAN and Dex pretreated podocytes with
miR-30a mimics or the miR-30a inhibitor resulted in decreased
(P<0.05) and increased proliferation, respectively (Fig. 3C).
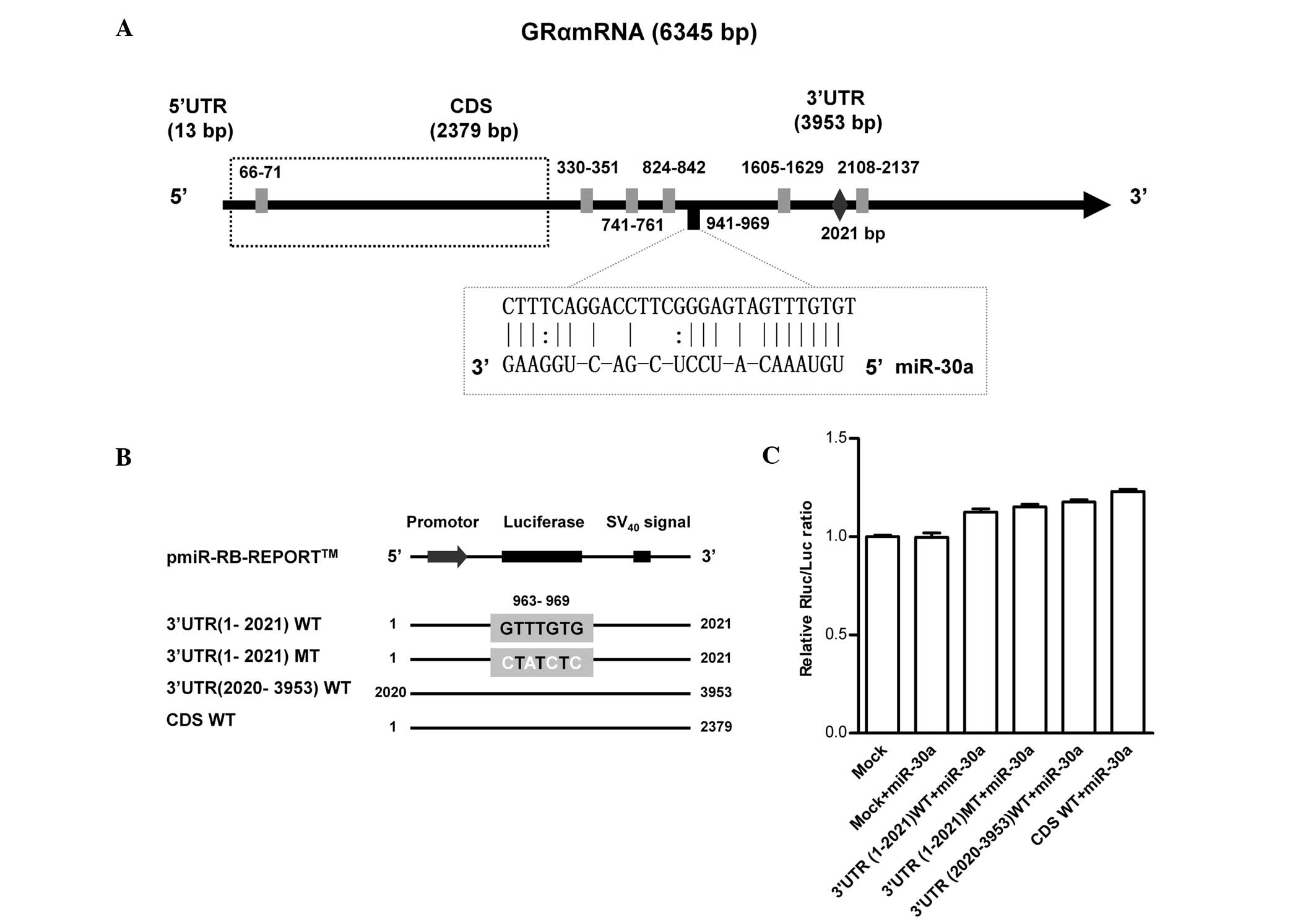
Expression of GRα is downregulated by
miR-30a interference, however, the 3′-UTR and CDS of GRα are
indirect miR-30a targets
To further confirm the association between miR-30a
and GRα, the targeted binding site was examined using a
dual-luciferase reporter system. Firstly, the consensus sequences
for miR-30a in the most common targeted binding site, the GRα
3′-UTR, were predicted. The full-length GRα 3′-UTR (3,953 bp) was
revealed to contain six predicted binding sites for miR-30a:
330–351, 741–761, 824–842, 941–969, 1605–1629 and 2108–2137 bp
(Fig. 4A). The site located at
941–969 bp was identified as the most likely binding site,
according to the continuity of seed sequences, the lowest free
energy and conservation between species. To confirm the predicted
consensus sequences and to determine whether these miR-30a seed
sequences directly contributed to the negative regulation of GRα
expression, substitution mutants (at positions 963, 965, 967 and
969) of the GRα 3′-UTR fragment were cloned into vectors containing
the WT miR-30a targeting sequences. Furthermore, analysis of other
possible candidates, including the GRα-5′-UTR and CDS, indicated
that nucleotides 66–71 bp of the CDS region is also a predicted
miR-30a binding consensus sequence (Fig. 4B). However, co-transfection of
murine cells with 50 nM miR-30a mimics revealed no change in
luciferase activity of the vectors, therefore, demonstrating that
miR-30a binds to either the 3′-UTR or CDS of GRα (Fig. 4C).
Discussion
GRα is a well-known receptor isoform, which mediates
gluco-corticoid functions. Investigation of the miRNA-regulation of
GRα in podocytes may shed new light on the mechanisms underlying
steroid-resistance in podocyte-associated nephrology. By comparing
a microarray miRNA expression profile in normal and injured murine
podocytes, the present study identified 10 candidate miRNAs
significantly upregulated in injured podocytes, along with the
downregulation of their potential target gene, GRα.
Since a certain regularity exists in the interaction
of miRNAs and target genes, computational algorithms, which are
fundamental to our understanding of the role of miRNAs in gene
regulation, have proved to be highly effective in predicting
candidate miRNAs, which can target an mRNA of interest (13,14).
However, this predicted correlation requires biological
confirmation in the context of several influencing factors
(15). The three most common
bioinformatics target prediction tools were used in the present
study and all indicated that GR was putative target gene of
miR-30a. However, the interaction between miR-30a and the GRα
isoform of GR remains to be elucidated experimentally. As classical
negative regulators in mammals, miRNAs were initially hypothesized
to mediate the repression of target proteins at the translational
level (16). Later,
high-throughput studies confirmed that miRNA-mediated changes occur
at the mRNA and protein levels, and are predominantly mediated by
target mRNA destabilization (17,18).
In the present study, it was identified that the marked reduction
in the mRNA expression of GRα mediated by miR-30a was in contrast
to the more moderate influence on the corresponding protein,
indicating that miR-30a regulates GRα expression at the
translational and transcriptional levels.
The podocyte is a terminally differentiated cell
with limited proliferative capacity and podocyte number is a
hallmark of progressive glomerulosclerosis (19,20).
Therefore, podocytic apoptosis is a critical determinant of the
progression of glomerulosclerosis and protection against podocytic
apoptosis is a vital means of slowing down glomerulosclerosis
(21). Additionally, previous
studies have suggested that miR-30a is involved in kidney
development, including podocyte differentiation (22–24).
Therefore, the present study aimed to determine whether miR-30a was
involved in podocytic apoptosis. Besides the conventional
anti-inflammatory or immunosuppressive actions, Dex causes modest
upregulation and increased nuclear localization of GR in podocytes,
and also enhances and accelerates podocytic maturation. PAN
treatment causes podocytic apoptosis, resulting in the development
of massive proteinuria resembling human podocytopathies. This
method has provided insights into the mechanisms of
podocyte-induced injury in vitro. Glucocorticoids can
protect and enhance recovery from PAN-induced injury in cultured
murine podocytes via actin filament stabilization (7). The protective effects of Dex on
PAN-induced podocyte apoptosis are associated with decreased
expression of p53, increased expression of Bcl-xl, inhibition of
apoptosis-inducing factor translocation and ERK phosphorylation
(25–27). This also remains controversial as
certain previous studies have demonstrated that Dex significantly
enhances podocyte number owing to increased survival of individual
cells as opposed to a reduction of apoptosis (4). To confirm the microarray results,
quantitative detection of miR-30a and the expression was higher in
the injured podocytes induced by PAN, confirming the microarray
results. However, Wu et al (28) previously documented that the
expression of the miR-30 family in human podocytopathies were
downregulated. The present study hypothesized that this may be
associated with the tissue and temporal specificity of miRNA.
Therefore, the present study investigated whether GRα was indeed
regulated by miR-30a and whether the observed reduction in podocyte
apoptosis, and upregulation in the expression of GRα following
PAN-induced injury, were mediated by miR-30a. These findings
demonstrated that transfection of miR-30a exerted a
negative-regulatory effect on the expression of GRα in PAN-injured
podocytes. In the present study, Dex was used at 1 µM, which
is consistent with clinical doses of glucocorticoid used in the
treatment of glomerular diseases and mimics the in vivo
therapeutic levels (4,5). Dex reduced PAN-induced podocytic
apoptosis and this effect was inhibited by the GR antagonist,
RU486. Therefore, it was suggested that the therapeutic effect of
Dex was GR-dependent. In addition, inhibition of miR-30a in
podocytes upregulated the expression of GRα, reduced apoptosis and
promoted proliferation, therefore, improving podocytic sensitivity
to Dex and favoring the therapeutic effect of steroids on podocyte
injury.
To further elucidate the direct interaction between
miR-30 and GRα, computational methods were used to predict the
miR-30a binding sites within the GRα mRNA sequence. Previous
studies have demonstrated that miRNAs modulate their target mRNAs
in mammalian cells by base pairing with complementary sites
commonly located within their 3′-UTR (29,30).
Previously, it was demonstrated that miRNA binding sites also exist
within the CDS and 5′-UTR (31,32).
Computational analysis predicted six potential binding sites for
miR-30a within the GRα 3′-UTR sequence, of which the site at
941–969 bp was the most likely based on complementary base pairing,
thermodynamic stability or conservation of seed matches. In
addition, the programs predicted a potential binding site within
the CDS. Luciferase reporters containing the full-length GRα CDS
and 3′-UTR, which included the predicted miR-30 binding sites and
harboring mutations at 963–969 bp, were generated. However, the
luciferase reporter assay data suggested an indirect effect on GRα
transcriptional activity since miR-30a failed to repress luciferase
activity. Since animal miRNAs have limited sequence complementarity
to their targets, it is a challenging task to predict miRNA targets
with high precision. In order to reduce the proportion of false
positives obtained through in silico prediction, the use of
>3 of the prediction programs is recommended for this purpose,
although Selbach et al (33,34)
reported that this approach is associated with precision levels of
only about 60%. Several novel computational target prediction
programs were identified to achieve precision reaching ~93%
(35,36). Accordingly, false positives
obtained through in silico prediction may account for the
negative result observed in the luciferase reporter assay in the
present study. However, the correlation between miR-30a and GRα at
the functional level was demonstrated in this study. A similar
effect was reported by Vreugdenhil et al (37) and revealed that miR-18 was
predicted to bind GR mRNA in silico and that miR-18
overexpression reduced GR protein levels. However, miR-18 was
unable to bind to the predicted seed region in the GR 3′-UTR. It
was suggested that the predicted miRNA binding sites were
unavailable for miRNA binding due to differences in the tertiary
RNA folding of the target gene sequence in the predicted and in
vivo situations (37).
Therefore, these data fail to completely preclude a potential
miR-30a effect on the gene expression of GRα. The functions of
miRNAs are particularly complicated. While a single miRNA is
thought to target numerous genes, multiple miRNAs are co-expressed
and are likely to mediate coordinated regulation of identical mRNAs
(28,38), resulting in corresponding multiplex
associations. Furthermore, synergy and antagonism also exist in the
interaction of miRNAs and represent an additional layer of
diversity to the function of miRNAs (39). The mechanisms by which miRNAs
mediate gene regulation are still open to discussion, and further
investigations are required to improve our understanding of the
function of each miRNA. Only in this way will the network of
interactions, which exist between miRNAs and target genes, be
better elucidated.
The major finding of the present study was that
silencing of miR-30a may have a potential effect on improving
steroid sensitivity in murine podocyte injury. However, although
miR-30a downregulates GRα at the functional level, the mechanism
requires further investigation.
Acknowledgments
This study was supported by a grant from the General
Program of National Natural Science Foundation of China (grant no.
81200522).
References
|
1
|
Pollak MR: Inherited podocytopathies: FSGS
and nephrotic syndrome from a genetic viewpoint. J Am Soc Nephrol.
13:3016–3023. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chuang PY and He JC: Signaling in
regulation of podocyte phenotypes. Nephron Physiol. 111:p9–p15.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Rhen T and Cidlowski JA: Anti-inflammatory
action of glucocorticoids-new mechanisms for old drugs. N Engl J
Med. 353:1711–1723. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Xing CY, Saleem MA, Coward RJ, Ni L,
Witherden IR and Mathieson PW: Direct effects of dexamethasone on
human podocytes. Kidney Int. 70:1038–1045. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Guess A, Agrawal S, Wei CC, Ransom RF,
Benndorf R and Smoyer WE: Dose-and time-dependent glucocorticoid
receptor signaling in podocytes. Am J Physiol Renal Physiol.
14:F845–F853. 2010. View Article : Google Scholar
|
|
6
|
Ransom RF, Lam NG, Hallett MA, Atkinson SJ
and Smoyer WE: Glucocorticoids protect and enhance recovery of
cultured murine podocytes via actin filament stabilization. Kidney
Int. 68:2473–2483. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Xie H, Lin HL, Chen JL, et al: Correlation
of expression of glucocorticoid receptorα and β in podocytes with
sensitivity of glucocorticoid in primary nephrotic syndrome. China
J Mod Med. 36:35–39. 2012.
|
|
8
|
Lin SL, Chang D, Wu DY and Ying SY: A
novel RNA splicing-mediated gene silencing mechanism potential for
genome evolution. Biochem Biophys Res Commun. 10:754–760. 2003.
View Article : Google Scholar
|
|
9
|
Shruti K, Shrey K and Vibha R: Micro RNAs:
Tiny sequences with enormous potential. Biochem Biophys Res Commun.
407:445–449. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lv M, Zhang X, Jia H, Li D, Zhang B, Zhang
H, Hong M, Jiang T, Jiang Q, Lu J, et al: An oncogenic role of
miR-142-3p in human T-cell acute lymphoblastic leukemia (T-ALL) by
targeting glucocorticoid receptor-α and cAMP/PKA pathways.
Leukemia. 26:769–777. 2012. View Article : Google Scholar
|
|
11
|
Tessel MA, Benham AL, Krett NL, Rosen ST
and Gunaratne PH: Role for microRNAs in regulating glucocorticoid
response and resistance in multiple myeloma. Horm Cancer.
2:182–189. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the (−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
13
|
Sethupathy P, Megraw M and Hatzigeorgiou
A: A guide through present computational approaches for the
identification of mammalian microRNA targets. Nat Methods.
3:881–886. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wang X: Computational prediction of
microRNA targets. Methods Mol Biol. 667:283–295. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ørom UA and Lund AH: Experimental
identification of microRNA targets. Gene. 451:1–5. 2010. View Article : Google Scholar
|
|
16
|
Humphreys DT, Westman BJ, Martin DI and
Preiss T: MicroRNAs control translation initiation by inhibiting
eukaryotic initiation factor 4E/cap and poly(A) tail function. Proc
Natl Acad Sci USA. 102:16961–16966. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Guo H, Ingolia NT, Weissman JS and Bartel
DP: Mammalian microRNAs predominantly act to decrease target mRNA
levels. Nature. 466:835–840. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Baek D, Villén J, Shin C, Camargo FD, Gygi
SP and Bartel DP: The impact of microRNAs on protein output.
Nature. 455:64–71. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kriz W, Elger M, Nagata M, Kretzler M,
Uiker S, Koeppen-Hageman I, Tenschert S and Lemley KV: The role of
podocytes in the development of glomerular sclerosis. Kidney Int.
45(Suppl): S64–S72. 1994.
|
|
20
|
Kriz W, Gretz N and Lemley KV: Progression
of glomerular diseases: Is the podocyte the culprit? Kidney Int.
54:687–697. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kim YH, Goyal M, Kurnit D, Wharram B,
Wiggins J, Holzman L, Kershaw D and Wiggins R: Podocyte depletion
and glomerulosclerosis have a direct relationship in the
PAN-treated rat. Kidney Int. 60:957–968. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Nagalakshmi VK, Ren Q, Pugh MM, Valerius
MT, McMahon AP and Yu J: Dicer regulates the development of
nephrogenic and ureteric compartments in the mammalian kidney.
Kidney Int. 385:1–14. 2010.
|
|
23
|
Agrawal R, Tran U and Wessely O: The
miR-30 miRNA family regulates Xenopus pronephros development and
targets the transcription factor Xlim1/Lhx1. Development.
136:3927–3936. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Harvey SJ, Jarad G, Cunningham J, Goldberg
S, Schermer B, Harfe BD, McManus MT, Benzing T and Miner JH:
Podocyte-specific deletion of dicer alters cytoskeletal dynamics
and causes glomerular disease. J Am Soc Nephrol. 19:2150–2158.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wada T, Pippin JW, Nangaku M and Shankland
SJ: Dexamethasone's prosurvival benefits in podocytes require
extracellular signal-regulated kinase phosphorylation. Nephron Exp
Nephrol. 109:e8–e19. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Koshikawa M, Mukoyama M, Mori K, Suganami
T, Sawai K, Yoshioka T, Nagae T, Yokoi H, Kawachi H, Shimizu F, et
al: Role of p38 mitogen-activated protein kinase activation in
podocyte injury and proteinuria in experimental nephrotic syndrome.
J Am Soc Nephrol. 16:2690–2701. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wada T, Pippin JW, Marshall CB, Griffin SV
and Shankland SJ: Dexamethasone prevents podocyte apoptosis induced
by puromycin aminonucleoside: Role of p53 and Bcl-2-related family
proteins. J Am Soc Nephrol. 16:2615–2625. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wu J, Zheng C, Fan Y, Zeng C, Chen Z, Qin
W, Zhang C, Zhang W, Wang X, Zhu X, et al: Downregulation of
microRNA-30 facilitates podocyte injury and is prevented by
glucocorticoids. J Am Soc Nephrol. 25:92–104. 2014. View Article : Google Scholar :
|
|
29
|
Carthew RW and Sontheimer EJ: Origins and
Mechanisms of miRNAs and siRNAs. Cell. 136:642–655. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Fabian MR, Sonenberg N and Filipowicz W:
Regulation of mRNA translation and stability by microRNAs. Annu Rev
Biochem. 79:351–379. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Roberts AP, Lewis AP and Jopling CL:
miR-122 activates hepatitis C virus translation by a specialized
mechanism requiring particular RNA components. Nucleic Acids Res.
39:7716–7729. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Schnall-Levin M, Zhao Y, Perrimon N and
Berger B: Conserved microRNA targeting in Drosophilia is as
widespread in coding regions as in 3′-UTRs. Proc Natl Acad Sci USA.
107:15751–15756. 2010. View Article : Google Scholar
|
|
33
|
Selbach M, Schwanhäusser B, Thierfelder N,
Fang Z, Khanin R and Rajewsky N: Widespread changes in protein
synthesis induced by microRNAs. Nature. 455:58–63. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Maragkakis M, Alexiou P, Papadopoulos GL,
Reczko M, Dalamagas T, Giannopoulos G, Goumas G, Koukis E, Kourtis
K, Simossis VA, et al: Accurate microRNA target prediction
correlates with protein repression levels. BMC Bioinformatics.
10:2952009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Reyes-Herrera PH, Ficarra E, Acquaviva A
and Macii E: miREE: miRNA recognition elements ensemble. BMC
Bioinformatics. 12:4542011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Mapleson D, Moxon S, Dalmay T and Moulton
V: MirPlex: A tool for identifying miRNAs in high-throughput sRNA
datasets without a genome. J Exp Zool B Mol Dev Evol. 320:47–56.
2013. View Article : Google Scholar
|
|
37
|
Vreugdenhil E, Verissimo CS, Mariman R,
Kamphorst JT, Barbosa JS, Zweers T, Champagne DL, Schouten T,
Meijer OC, de Kloet ER, et al: MicroRNA 18 and 124a down-regulate
the glucocorticoid receptor: Implications for glucocorticoid
responsiveness in the brain. Endocrinology. 150:2220–2228. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Bartel DP: MicroRNAs: Target recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Breving K and Esquela-Kerscher A: The
complexities of microRNA regulation: mirandering around the rules.
Int J Biochem Cell Biol. 42:1316–1329. 2010. View Article : Google Scholar
|