Introduction
Pancreatic cancer is one of the most life
threatening and aggressive types of malignancy and has a global
annual incidence rate of ~8/100,000 individuals (1). Due to recurrence, the 5-year survival
rate is ~3–5% (2,3). According to its histogenetic origin,
pancreatic cancer can be classified into several types (4), and adenocarcinoma is the most common
histological type (5). Generally,
genetic and environmental factors are involved in the development
of pancreatic cancer (6,7).
Numerous studies have focused on the molecular
mechanisms of pancreatic cancer. Genetic alterations in several
genes, including KRAS, TP53, p16/CDKN2A, SMAD4, BRCA2, STK11, PRSS1
and BRAF30 have been demonstrated to be involved in the development
of pancreatic cancer (8–12). Several pathways, including the
Raf/extracellular signal-regulated kinase pathway, the
phosphoinositide 3-kinase pathway and the Ral guanine nucleotide
dissociation stimulator pathway have also been identified to
contribute to the initiation and progression of pancreatic cancer
(13,14). Furthermore, the above-mentioned
genes and pathways are involved in the proliferation, invasion,
metastasis and apoptosis of pancreatic cancer (15).
Toll-like receptors (TLRs) have been demonstrated to
be involved in the pathogenesis of the pancreatic cancer (16,17).
Ochi et al (18) reported a
novel role of TLR7 in the development and progression of pancreatic
cancer through interference with cell cycle regulation and
activation of multiple cell signaling pathways.
TLR7 is one of the pattern recognition receptors,
which has been identified as a primary detector of bacterial and
viral components (19). TLR7 can
trigger a series of signaling pathways by binding to
single-stranded RNA or small molecular chemicals, including
imiquimod and its derivatives (20). The effect of TLR7 agonists on
cancer cells remain contradictory (21,22).
In renal cells and bladder cancer cells, TLR7 agonists exhibit
anti-proliferative and apoptosis-inducing effects by suppressing
the expression of c-Myc and directly affecting cell growth and
tumorigenesis (23,24). By contrast, activation of TLR7 has
been reported to promote the survival, proliferation and
invasiveness of tumor cells, including B-cell chronic lymphocytic
leukemia cells (25–27). Thus, the effects of TLR agonists on
tumor cells require further investigation (28).
In the present study, the effects of the TLR7
agonist, gardiquimod, on the proliferation, the migration and the
apoptosis of the BxPC-3 pancreatic cancer cell line were
investigated.
Materials and methods
Reagents
The BxPC-3 human pancreatic cancer cell line cells
were purchased from the Shanghai Cell Bank of Chinese Academy of
Sciences (Shanghai, China). Gardiquimod was obtained from InvivoGen
(San Diego, CA, USA).
3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxyme-thoxy-phenyl)-2-(4-sulfophenyl)-2H-tetrazolium,
inner salt (MTS) was obtained from Promega Corporation (Madison,
WI, USA). Antibodies against β-actin (cat. no. sc-8432; mouse
monoclonal IgG1), cyclin E (cat. no. sc-247; mouse monoclonal
IgG2b), cyclin B1 (cat. no. sc-594; rabbit polyclonal IgG), B-cell
lymphoma (Bcl)-2 (cat. no. sc-7382; mouse monoclonal IgG1) and
B-cell-associated X protein (Bax; cat. no. sc-493, rabbit
polyclonal IgG) were purchased from Santa Cruz Biotechnology, Inc.
(Santa Cruz, CA, USA). Fluorescein isothiocyanate (FITC)-Annexin V
and propidium iodide (PI) were obtained from BioSharp (Hefei,
China). Primers (Table I) were
purchased from Sangon Biotech Co. Ltd. (Shanghai, China). Trypsin,
Triton X-100, RNase A, radioimmunoprecipitation (RIPA) assay
buffer, phenylmethylsulfonyl fluoride (PMSF), bicinchoninic acid
(BCA) assay kit and SDS-polyacrylamide gel electrophoresis (PAGE)
gels were obtained from Sigma-Aldrich (St. Louis, MO, USA) and
Tris-buffered saline with Tween-20 (TBST) was purchased from
Sunshine Biotechnology Co., Ltd. (Nanjing, China). Fetal bovine
serum (FBS), streptomycin and penicillin were obtained from
Invitrogen Life Technologies (Shanghai, China).
 | Table IPrimers used for reverse
transcription-quantitative polymerase chain reaction analysis. |
Table I
Primers used for reverse
transcription-quantitative polymerase chain reaction analysis.
| Gene | Primer
sequence |
|---|
| GAPDH | Forward
5′-AGATCATCAGCAATGCCTCCTG-3′ |
| Reverse
5′-ATGGCATGGACTGTGGTCATG-3′ |
| TLR7 | Forward
5′-TAGGATCACTCCATGCCATCAA-3′ |
| Reverse
5′-CAGTGTCCACATTGGAAACACC-3′ |
Peripheral blood monouclear cell (PBMC)
preparation
EDTA anticoagulated venous blood samples (2 ml) were
collected from healthy volunteers and PBMCs were isolated using
lymphocyte separation medium (Tianjin Haoyang Biological
Manufacture Co., Ltd., Tianjin, China). The expression of TLR7 in
PBMCs was analyzed and served as a positive control. Written
informed consent was obtained from the volunteers.
Cell culture
The BxPC-3 cells were cultured for 2 days in
RPMI-1640 medium supplemented with 10% FBS, 100 U/ml streptomycin
and 100 U/ml penicillin at 37°C in a 5% CO2 atmosphere.
Upon reaching 70% confluence, the cells were passaged for use in
the following experiments.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
BxPC-3 cells were seeded (3×103
cells/well) in 24-well plates and cultured in FBS-free media for 3
days. Total cellular RNA was extracted from the cells using TRIzol
(Invitrogen Life Technologies, Carlsbad, CA, USA). RT-qPCR analysis
of TLR7 and GAPDH were performed, as previously described (29). Briefly, samples were amplified in
an Applied Biosystem 7500 Real-Time PCR System (Applied Biosystems
Life Technologies, Foster City, CA, USA) for 40 cycles with the
following conditions: Denaturation at 95°C for 15 sec, and
annealing and extension at 60°C for 40 sec. The primer sequences
used are presented in Table I and
the relative expression of TLR7 was calculated using
2−ΔΔCt.
MTS assay
The BxPC-3 cells were seeded (3×103
cells/well) in 96-well plates and cultured in FBS-free RPMI-1640
for 1 day. The cells were continually cultured at 37°C in a 5%
CO2 atmosphere and treated with different concentrations
(0, 1.5, 3 and 5 µg/ml) of gardiquimod for various
time-periods (0, 12 and 30 min, 1, 3, 6, 12, 24 and 48 h),
following which 20 µl MTS reagent was added. The cells were
incubated for 2 h and absorbance was measured at 490 nm using a
Spectrophotometer Multiskan GO (Thermo Fisher, Vantaa, Finland).
The experiment was repeated a minimum of three times.
Flow cytometric analysis
The BxPC-3 cells were seeded in 12-well plates and
treated with 3 µg/ml gardiquimod for different time-periods,
between 1 and 5 days, in FBS-free RPMI-1640 at 37°C in a 5%
CO2 atmosphere. The cells were detached with trypsin and
washed with phosphate-buffered saline (PBS), and were then BxPC-3
cells were suspended at a density of 3×105/ml and fixed
in 70% alcohol for 30 min. Subsequent to washing with PBS, 5
µl Triton X-100, 3 µl RNase A (20 ug/ml) and 1
µl PI dye (50 ug/ml) were added to the cells. Following
mixing at room temperature and maintenance in the dark for 30 min,
flow cytometric analysis was performed. For analysis of apoptosis
in the cells, 5 µl FITC-Annexin V and 5 µl PI were
added to the resuspended cells at room temperature for 15 min. Then
the cells were detected by flow cytometry using a BD FACSVerse™
flow cytometer (BD Biosciences, San Jose, CA, USA). The cells in
the lower left, lower right, upper right and upper left quadrant
signified viable, early apoptotic, late apoptotic or necrotic
cells, respectively. The experiment was repeated three times.
Western blot analysis
The BxPC-3 cells were cultured in FBS-free RPMI-1640
at 37°C in a 5% CO2 atmosphere, following which
gardiquimod (3 ug/ml) was added and the cells were cultured for 0,
12 and 30 min, 1, 3, 6, 12, 24 and 48 h. Subsequent to washing with
cold PBS, the cells were incubated in RIPA assay buffer containing
1 mM PMSF fluoride for 30 min in an ice-water bath. Subsequent to
protein quantification using a BCA method, 100 µg proteins
were loaded onto 12% SDS-PAGE gels and the separated proteins were
transferred onto polyvinylidene difluoride membranes (EMD
Millipore, Billerica, MA, USA). The membrane was blocked in TBST
containing 5% non-fat milk for 2 h at room temperature with
agitation. The membranes were then incubated with primary
antibodies against β-actin, cyclin B1, cyclin E, Bcl-2 and Bax
(1:500) overnight at 4°C. Subsequent to washing with TBST, the
membrane was incubated in horseradish peroxidase-linked secondary
antibody/TBST solution for 2 h at room temperature. The bound
secondary antibodies were detected using chemiluminescence
substrate (Pierce Biotechnology, Inc., Woburn, MA, USA) and were
visualized by exposure to X-ray film. The band intensity was
determined using a Gel Image Analysis system and were normalized to
β-actin.
Scratch/migration assay
The BxPC-3 cells were cultured in 24-well dishes
(5×104 cells/well) for 24 h at 37°C in a 5%
CO2 atmosphere. A 10-µl pipette tip was used to
scratch three perpendicular lines on the bottom of dishes.
Following washing with PBS, gardiquimod (3 µg/ml) was added
to the cells for different time periods (1–5 days) at 37°C in a 5%
CO2 atmosphere. The distance that the cells migrated
into the scratched region was quantified using the ocular
micrometer of a Nikon Ecolipse TS100 microscope (Nikon Corporation,
Tokyo, Japan; 10X objective lens, 1 unit = 10 mm). The assessment
was repeated three times.
Statistical analysis
All experiments were repeated a minimum of three
times and the representative results are presented. The data are
presented as means ± standard deviation. Student's t-test, analysis
of variance and the χ2 analysis were conducted using
SPSS software, version 13.0 (SPSS, Inc., Chicago, IL, USA) and
P<0.05 was considered to indicate a statistically significant
difference.
Results
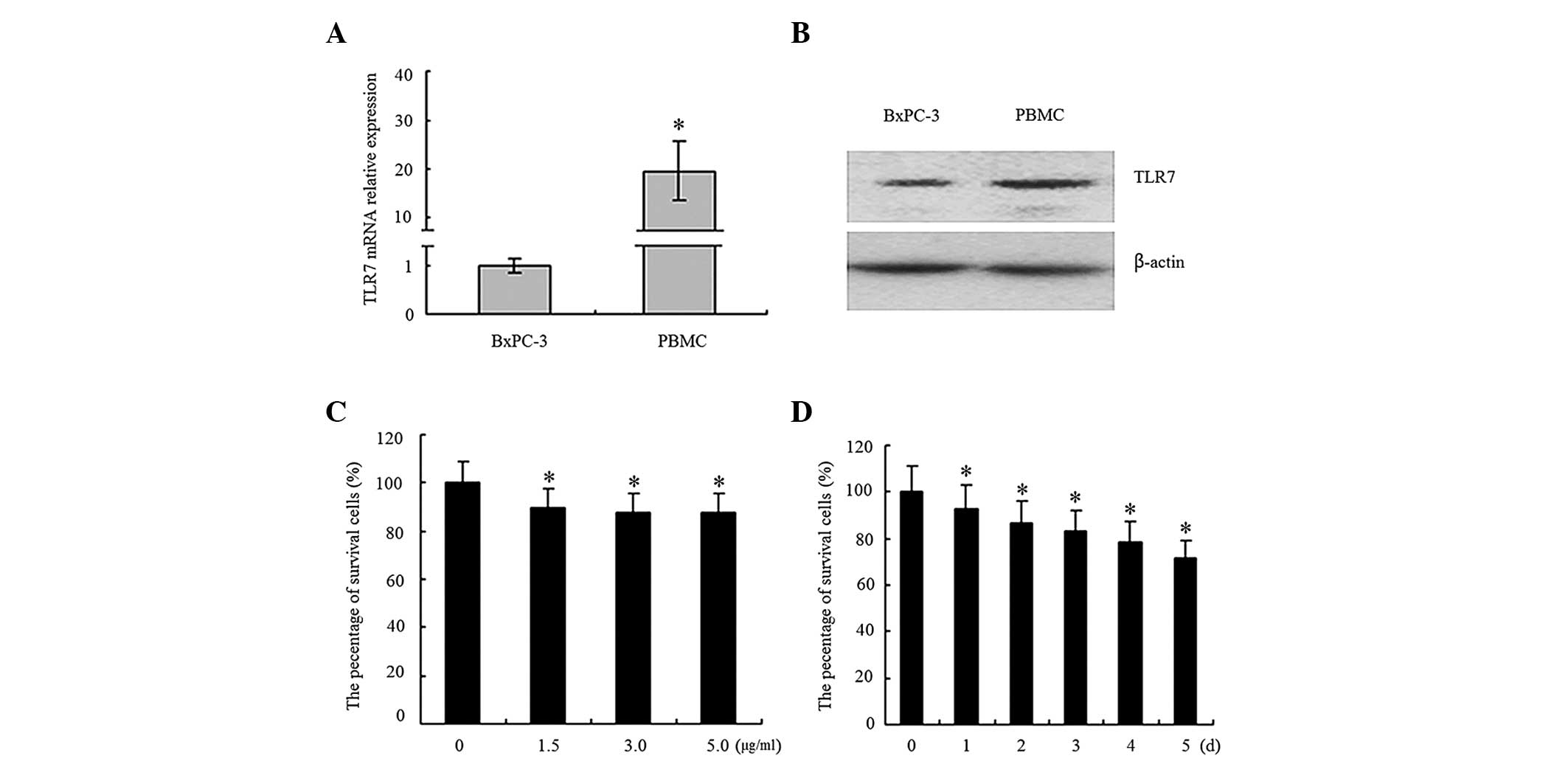
TLR7 is expressed in BxPC-3 cells
In the present study, total RNA and protein were
extracted from the BxPC-3 cells, following which RT-qPCR and
western blot analysis were used to analyze the mRNA and protein
expression levels of TLR, respectively. Peripheral blood
mononuclear cells (PBMCs) were used as a positive control sample.
Compared with the PBMCs, the expression of TLR7 in the BxPC-3 cells
was reduced at the mRNA (Fig. 1A)
and protein (Fig. 1B) levels.
Gardiquimod inhibits the proliferation of
BxPC-3 cells in a dose- and time-dependent manner
Treatment of gardiquimod inhibited the proliferation
of the BxPC3 cells. The inhibition ratio was 10.41% at a low
concentration (1.5 µg/ml) and 20.12% at a high concentration
(5 µg/ml; P<0.05; Fig.
1C). The time course analysis indicated that the inhibition
ratio gradually increased between 7.01 and 28.55% between days 1
and 5 following the addition of 3 µg/ml gardiquimod
(P<0.05; Fig. 1D).
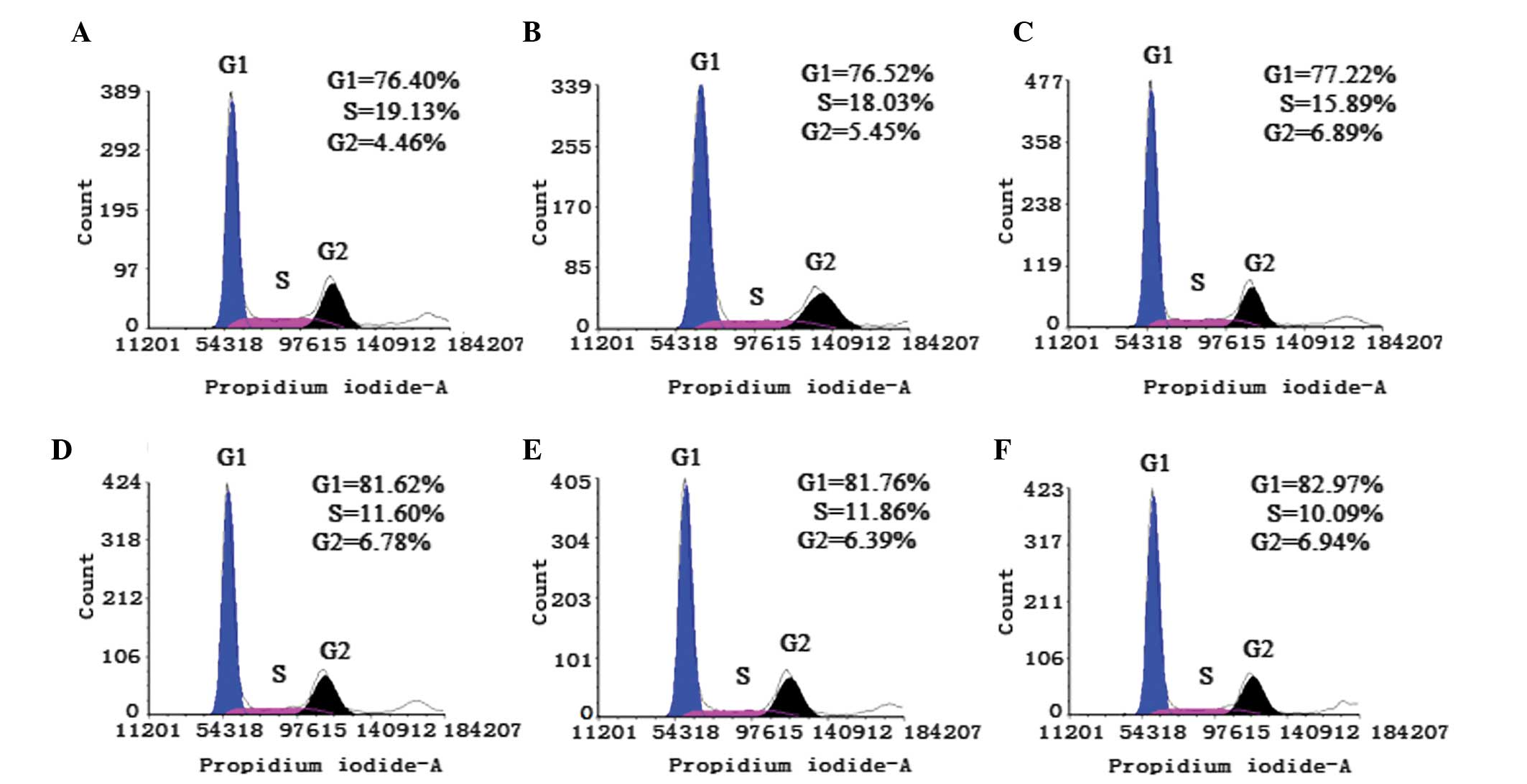
Analysis of the cell cycle analysis using flow
cytometry demonstrated that the percentage of BxPC-3 cells treated
with gardiquimod in the G1 phase gradually increased
between 76.41 and 82.97% (Fig. 2).
In addition, the percentage of cells in the S phase reduced between
19.13 and 10.09% as the duration of treatment increased (Fig. 2).
Gardiquimod induces the apoptosis of
BxPC-3 cells
The effect of gardiquimod on the apoptosis of BxPC-3
cells was also examined (Fig. 3).
The percentage of cells in the early apoptotic phase increased
between 11.5% at day 1 and 19.32% at day 5. There was a
statistically significant difference between the control group
(Fig. 3A) and the groups treated
with gardiquimod for >3 days (P<0.05; Fig. 3D–F).
Gardiquimod reduces the expression levels
of cyclin B1, cyclin E and Bcl-2, and increases the protein
expression levels of Bax
Based on the results of the MTS and flow cytometry,
gardiquimod was found to inhibit the proliferation and induce the
apoptosis of the BxPC-3 cells. In order to investigate the
underlying mechanism, western blot analysis was performed to
analyze changes in the expression levels of cyclin B1, cyclin E,
Bcl-2 and Bax in the BxPC-3 cells treated with gardiquimod. As
shown in Fig. 4A, treatment with
gardiquimod reduced the expression levels of cyclin B1, cyclin E
and Bcl-2 in the BxPC-3 cells and, following treatment for 12 h,
the expression levels of these three proteins markedly reduced. By
contrast, gardiquimod induced the expression of Bax.
 | Figure 4Gardiquimod reduces the expression
levels of cyclin B1, cyclin E and Bcl-2, increases the expression
of Bax at the protein level and inhibits the migration of BxPC-3.
(A) BxPC-3 cells were treated with gardiquimod (3 µg/ml) for
various time-periods and the expression levels of cyclin B1, cyclin
E, Bcl-2 and Bax were determined using western blot analysis. The
results represent one of three independent experiments. (B) BxPC-3
cells were treated with gardiquimod (3 µg/ml) for 1 day,
following which the surface of the cell layer was scratched and
continually cultured for 6 days, monitoring the scratched space
daily under a microscope (magnification, ×100). Bcl, B-cell
lymphoma; Bax, B-cell-associated X protein; G, gariquimod-treated;
Con, untreated. |
Gardiquimod inhibits the migration of
BxPC-3 cells
Cancer cells can migrate from their original sites
and invade into adjacent normal tissues or into the blood or lymph
vessels, and form additional neoplasms. In the present study, a
scratch/migration assay was used to detect the effects of
gardiquimod on the migration of BxPC-3 cells. In the control group,
without gardiquimod treatment, the BxPC-3 cells occupied almost the
entire scratched space at day 5, however, in the groups treated
with gardiquimod, ~60% of the space present at day 0 remained
unoccupied at day 5 (Fig. 4B).
Discussion
Pancreatic cancer is one of the five most common
causes of cancer-associated mortality (30). In the present study, gardiquimod
was used to treat BxPC-3 pancreatic cancer cells, and the results
demonstrated that gardiquimod inhibited the proliferation and
migration, and induced the apoptosis of the BxPC-3 cells. The
effect of gardiquimod on the proliferation of the BxPC-3 cells may
be involved in the regulation of the expression levels of cyclin
B1, cyclin E, Bcl-2 and Bax.
Previous studies have demonstrated that TLR7
signaling pathways have different roles in different types of
cancer. TLR7 agonists have been suggested to be antiproliferative
and induce apoptosis in renal cancer cells and bladder cancer
cells, including normal T cells (23,24).
By contrast, they can promote the survival, proliferation and
migration of tumor cells, including B-cell chronic lymphocytic
leukemia cells (25–27). Ochi et al (18) previously demonstrated that the TLR7
agonist, ssRNA40, stimulates the proliferation of pancreatic cancer
cells, but had no effect on the pancreatic cells of wild mice
(18). In addition, the activation
of TLR7 was demonstrated to regulate the expression levels of a
series of genes, including phosphatase and tensin homolog, p16,
p21, p27, p53, Rb, c-Myc, transforming growth factor-β and SHPTP1
(18). The results of the present
study appear to be in contradiction with the observations of Ochi
et al (18), however, the
present study used BxPC-3 cells as a model, unlike the pancreatic
cancer cells from p48Cre; KrasG12D mice,
which were used as the model in the study by Ochi et al. The
Kras gene was sequenced in the present study, and no
KrasG12D mutation was identified (data not shown). This
suggested that cell lines with different genetic backgrounds may
have entirely different responses to treatment with gardiquimod.
These paradoxical results suggest that the TLR7 agonists have
opposite effects, even in different cell lines from the same type
of cancer. However, the molecular mechanism of this phenomena
requires further investigation, which may assist in the development
of novel therapeutic methods for cancer.
The expression of cyclin is a key element in the
control of the cell cycle. In the present study, gardiquimod was
observed to reduce the expression levels of cyclin B1 and cyclin E,
which suggested that gardiquimod may exert its antiproliferative
effects at the transition of the G1-S phase. It has been
previously reported that gardiquimod exerts its antiproliferative
activity in urothelial cell carcinoma via downregulating the
expression of cyclin D2 (24, therefore, the antiproliferative
mechanism of gardiquimod appears similar between BxPC-3 cells and
urothelial cell carcinoma (24).
On analyzing apoptosis in the present study, gardiquimod was found
to induce apoptosis; which was consistent with its effect on renal
cells (31). It is well known that
the apoptosis-associated proteins, Bcl-2 and Bax (32), are important in the process of
apoptosis in cells (33,34). In the present study, gardiquimod
was observed to regulate the expression of these two genes. All
these results suggested that gardiquimod may exert its
antiproliferative activity and induce apoptosis on the BxPC-3 cells
via the regulation of proliferation and the expression levels of
apoptosis-associated genes. These results also provide insight into
the pathogenesis of pancreatic cancer, potentially and may assist
in the development of novel treatment strategies. However, there
are several details, which remain to be fully elucidated, thus, the
effects of TLR agonists on tumor cells require careful further
investigation prior to their application in immunotherapy. The
present study also assessed the effects of gardiquimod on the
migration of the BxPC-3 cells and demonstrated that, following
wounding, the cells treated with gardiquimod healed more slowly,
compared with those in the control group. The molecular mechanism
underlying this phenomenon remains to be elucidated and requires
investigation in subsequent investigations.
In conclusion, the results of the present study
suggested that gardiquimod inhibited the proliferation and
migration, and induced the apoptosis of BxPC-3 pancreatic cancer
cells, which may assist in elucidating the involvement of TLR7 in
the development of pancreatic cancer.
Acknowledgments
This study was supported by grants from the General
Program of the National Natural Science Foundation of China (grant.
no. 81271748) and the Foundation for Doctors, Anhui Medical
University (Hefei, China; grant no. 0108016101).
References
|
1
|
Raimondi S, Maisonneuve P and Lowenfels
AB: Epidemiology of pancreatic cancer: An overview. Nat Rev
Gastroenterol Hepatol. 6:699–708. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Steele CW, Jamieson NB, Evans TR, McKay
CJ, Sansom OJ, Morton JP and Carter CR: Exploiting inflammation for
therapeutic gain in pancreatic cancer. Br J Cancer. 108:997–1003.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Jemal A, Siegel R, Ward E, Hao Y, Xu J and
Thun MJ: Cancer statistics. CA Cancer J Clin. 59:225–249. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Cooper CL, O'Toole SA and Kench JG:
Classification, morphology and molecular pathology of premalignant
lesions of the pancreas. Pathology. 45:286–304. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Alexakis N, Halloran C, Raraty M, Ghaneh
P, Sutton R and Neoptolemos JP: Current standards of surgery for
pancreatic cancer. Br J Surg. 91:1410–1427. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lin Y, Yagyu K, Egawa N, Ueno M, Mori M,
Nakao H, Ishii H, Nakamura K, Wakai K, Hosono S, et al: An overview
of genetic polymorphisms and pancreatic cancer risk in molecular
epide-miologic studies. J Epidemiol. 21:2–12. 2011. View Article : Google Scholar
|
|
7
|
Nitsche C, Simon P, Weiss FU, Fluhr G,
Weber E, Gärtner S, Behn CO, Kraft M, Ringel J, Aghdassi A, et al:
Environmental risk factors for chronic pancreatitis and pancreatic
cancer. Dig Dis. 29:235–242. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Iacobuzio-Donahue CA, Velculescu VE,
Wolfgang CL and Hruban RH: The genetic basis of pancreas cancer
development and progression: Insights from whole-exome and
whole-genome sequencing. Clin Cancer Res. 18:4257–4265. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Jones S, Zhang X, Parsons DW, Lin JC,
Leary RJ, Angenendt P, Mankoo P, Carter H, Kamiyama H, Jimeno A, et
al: Core signaling pathways in human pancreatic cancers revealed by
global genomic analyses. Science. 321:1801–1806. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Petersen GM, de Andrade M, Goggins M,
Hruban RH, Bondy M, Korczak JF, Gallinger S, Lynch HT, Syngal S,
Rabe KG, et al: Pancreatic cancer genetic epidemiology consortium.
Cancer Epidemiol Biomarkers Prev. 15:704–710. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Klein AP, Brune KA, Petersen GM, Hruban
RH, Bondy M, Korczak JF, Gallinger S, Lynch HT, Syngal S, Rabe KG,
et al: Prospective risk of pancreatic cancer in familial pancreatic
cancer kindreds. Cancer Res. 64:2634–2638. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Shi C, Hruban RH and Klein AP: Familial
pancreatic cancer. Arch Pathol Lab Med. 133:365–374.
2009.PubMed/NCBI
|
|
13
|
Siveke JT, Einwächter H, Sipos B,
Lubeseder-Martellato C, Klöppel G and Schmid RM: Concomitant
pancreatic activation of Kras (G12D) and Tgfa results in cystic
papillary neoplasms reminiscent of human IPMN. Cancer Cell.
12:266–279. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ma J, Sawai H, Ochi N, Matsuo Y, Xu D,
Yasuda A, Takahashi H, Wakasugi T and Takeyama H: PTEN regulates
angiogenesis through PI3K/Akt/VEGF signaling pathway in human
pancreatic cancer cells. Mol Cell Biochem. 331:161–171. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Mihaljevic AL, Michalski CW, Friess H and
Kleeff J: Molecular mechanism of pancreatic cancer-understanding
proliferation, invasion and metastasis. Langenbecks Arch Surg.
395:295–308. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ikebe M, Kitaura Y, Nakamura M, Tanaka H,
Yamasaki A, Nagai S, Wada J, Yanai K, Koga K, Sato N, et al:
Lipopolysaccharide (LPS) increases the invasive Aability of
pancreatic cancer cells through the TLR4/MyD88 signaling pathway. J
Surg Oncol. 100:725–731. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Mai CW, Kang YB and Pichika MR: Should a
Toll-like receptor 4 (TLR-4) agonist or antagonist be designed to
treat cancer? TLR-4: its expression and effects in the ten most
common cancers. Onco Targets Ther. 6:1573–1587. 2013.PubMed/NCBI
|
|
18
|
Ochi A, Graffeo CS, Zambirinis CP,
Zambirinis CP, Rehman A, Hackman M, Fallon N, Barilla RM, Henning
JR, Jamal M, et al: Toll-like receptor 7 regulates pancreatic
carcinogenesis in mice and humans. J Clin Invest. 122:4118–4129.
2012. View
Article : Google Scholar : PubMed/NCBI
|
|
19
|
Olivier M: Host-pathogen interaction:
Culprit within a culprit. Nature. 471:173–174. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhu J, Lai K, Brownile R, Babiuk LA and
Mutwiri GK: Porcine TLR8 and TLR7 are both activated by a selective
TLR7 ligand, imiquimod. Mol Immunol. 45:3238–3243. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Shojaei H, Oberg HH, Juricke M, Marischen
L, Kunz M, Mundhenke C, Gieseler F, Kabelitz D and Wesch D:
Toll-like Receptors 3 and 7 Agonists Enhance Tumor Cell T Cells
Lysis by Human. Cancer Res. 69:8710–8717. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kutikhin AG: Association of polymorphisms
in TLR genes and in genes of the Toll-like receptor signaling
pathway with cancer risk. Hum Immunol. 72:1095–1116. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Schwartz MJ, Liu H, Hwang DH, Kawamoto H
and Scherr DS: Antitumor effects of an imidazoquinoline in renal
cell carcinoma. Urology. 73:1156–1162. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liu H, Schwartz MJ, Hwang DH and Scherr
DS: Tumour growth inhibition by an imidazoquinoline is associated
with c-Myc down-regulation in urothelial cell carcinoma. BJU Int.
101:894–901. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Funderburg N, Luciano AA, Jiang W,
Rodriguez B, Sieg SF and Lederman MM: Toll-like receptor ligands
induce human T cell activation and death, a model for HIV
pathogenesis. PLoS One. 3:e19152008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
François S, El Benna J, Dang PM, Pedruzzi
E, Gougerot-Pocidalo MA and Elbim C: Inhibition of neutrophil
apoptosis by TLR agonists in whole blood: involvement of the
phosphoinositide3-kinase/Akt and NF-kappaB signaling pathways,
leading to increased levels of Mcl-1, A1 and phosphorylated Bad. J
Immunol. 174:3633–3642. 2005. View Article : Google Scholar
|
|
27
|
Hammadi A, Billard C, Faussat AM and Kolb
JP: Stimulation of iNOS expression and apoptosis resistance in
B-cell chronic lymphocytic leukemia (B-CLL) cells through
engagement of Toll-like receptor 7 (TLR-7) and NF-kappaB
activation. Nitric Oxide. 19:138–145. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zambirinis CP and Miller G: Signaling via
MYD88 in the pancreatic tumor microenvironment. Oncoimmunology.
2:e225672013. View Article : Google Scholar
|
|
29
|
Li L, Cheng FW, Wang F, Jia B, Luo X and
Zhang SQ: The activation of TLR7 regulates the expression of VEGF,
TIMP1, MMP2, IL-6 and IL-15 in Hela cells. Mol Cell Biochem.
389:43–49. 2014. View Article : Google Scholar
|
|
30
|
Bosetti C, Bertuccio P, Negri E, La
Vecchia C, Zeegers MP and Boffetta P: Pancreatic cancer: overview
of descriptive epidemiology. Mol Carcinog. 51:3–13. 2012.
View Article : Google Scholar
|
|
31
|
Schwartz MJ, Liu H, Hwang DH, Kawamoto H
and Scherr DS: Antitumor effects of an imidazoquinoline in renal
cell carcinoma. Urology. 73:1156–1162. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Sasi N, Hwang M, Jaboin J, Csiki I and Lu
B: L Regulated cell death pathways: new twists in modulation of
BCL2 family function. Mol Cancer Ther. 8:1421–1429. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Pietrantonio F, Biondani P, Ciurlia E,
Fanetti G, Tessari A, Bertarelli G, Bossi I, Musella V, Melotti F,
Di Bartolomeo M, et al: Role of BAX for outcome prediction in
gastrontestinal malignancies. Med Oncol. 30:6102013. View Article : Google Scholar
|
|
34
|
Rubenstein M, Hollowell CM and Guinan P:
Differentiated prostatic antigen expression in cells following
treatment with bispecific antisense oligonucleotides directed
against BCL-2 and EGFR. Med Oncol. 29:835–841. 2012. View Article : Google Scholar
|