Introduction
Nasopharyngeal carcinoma (NPC) is a tumor, which
arises from the epithelial cells of the nasopharynx. NPC can be
classified into three subtypes: Squamous cell carcinoma,
non-keratinizing carcinoma and undifferentiated carcinoma. The
exact etiology of NPC remains to be elucidated, however, it has
been suggested that Epstein-Barr virus may be one of the causes of
NPC, since it has been reported to be associated with epithelial
cell transformation into NPC type 2 and 3 (1,2). In
addition, type 2 (non-keratinizing carcinoma) and 3
(undifferentiated carcinoma) NPCs are associated with increased
titers of the virus (3). Diet,
genetic factors, gender and ethnicity have also been regarded as
possible risk factors of NPC.
NPC is a rare malignancy in the majority of
countries, with an incidence rate of <1 case per 100,000
individuals annually. However, the incidence rate is higher (20–50
cases per 100,000 individuals per year) in southern China and
southeast Asia. A combination of radiation and chemotherapy is the
current predominant therapeutic strategy for the treatment of
patients with locally and regionally confined tumors (4). However, the majority of established
chemotherapies are associated with intolerable side effects,
including dizziness, vomiting and constipation. Furthermore,
certain patients may not respond well to treatment or may develop
resistance following long term usage. Therefore, alternative
medicine, including the use of medicinal plants, is becoming
essential. The use of medicinal plants has been extensively
investigated for several years.
Malaysia is a megadiverse country with a high number
of endemic vascular plant species (5), as well as several types of medicinal
plants and herbs, which have been widely used in traditional
medicine. For example, Agrimony eupatoria has been used to
treat diabetes (6), and
Centella asiatica can be used for wound healing (7). Another well-known medicinal plant is
Strobilanthes crispa Blume, which is termed pecah-kaca or
jinbatu in Malaysia; enyohkelo, kecibeling, kejibeling or ngokilo
in Java; and daun pecahbeling in Jakarta. S. crispa is a
flowering shrub, which belongs to the Acanthaceae family, and is
distributed throughout regions of Madagascar to Indonesia (8). S. crispa can be readily found
in forests, riverbanks and fields.
S. crispa has been used extensively in
various regions as a traditional remedy to heal a wide range of
diseases. Infusion of the dried leaves of S. crispa has been
suggested to possess antidiabetic, diuretic, antilithic and
laxative properties (9). In
addition, fresh leaves may be masticated and swallowed, to benefit
the immune system (10). The water
extract of S. crispa has been reported to inhibit the
proliferation of retroviruses (11). Previously, S. crispa extract
has demonstrated inhibitory properties against breast (12–17),
liver (14,16–20),
prostate (12) and colon (14) carcinomas. These findings have
encouraged further investigation regarding the use of this plant
for its anticancer properties. There are currently no reports
regarding the anticancer effects of S. crispa on NPC.
Therefore, the present study aimed to determine the cytotoxic and
apoptogenic effects of S. crispa on NPC. Furthermore, the
mechanism of action of apoptosis through caspase activation was
investigated.
Materials and methods
Plant extraction procedures
The fresh leaves and stems of S. crispa were
collected from Paka (Terengganu, Malaysia), and oven dried at a
temperature of 40°C until a constant weight was obtained. The
botanical identity of the plant was determined and authenticated by
a taxonomist from the Forest Research Institute Malaysia (Kuala
Lumpur, Malaysia; sample number: PID 040114-04). Water and organic
solvents were used to perform plant extraction. To prepare the
water extract, 10 g dried S. crispa was mixed with 500 ml
distilled water and maintained at 60°C for 3 h. The resulting
suspension was then filtered and freeze-dried. In addition,
stepwise extraction using solvents of increasing polarity (hexane,
chloroform, ethyl acetate and methanol) was performed to prepare
the organic solvent extracts (14). Briefly, 100 g dried S.
crispa was mixed with 500 ml organic solvent and maintained in
the dark for 3 days at room temperature. The samples were
subsequently filtered using Whatman paper (Thermo Fisher
Scientific, Waltham, MA, USA) and the resulting filtrates were
evaporated using a rotary evaporator (Buchi R-2154; Buchi, Flawil,
Switzerland). Finally, 10 different plant extracts were obtained,
as follows: Leaves hexane (LH), leaves chloroform (LC), leaves
ethyl acetate (LEA), leaves methanol (LM), leaves water (LW), stems
hexane (SH), stems chloroform (SC), stems ethyl acetate (SEA),
stems methanol (SM) and stems water (SW).
Cell culture
The CNE-1 human NPC cell line was provided by
Professor Sam Choon Kook (Institute of Biological Sciences,
University Malaya, Kuala Lumpur, Malaysia), and the NRK-52E normal
neonatal rat kidney epithelial cell line was purchased from
American Type Culture Collection (Manassas, VA, USA). The cell
lines were cultured in Dulbecco's modified Eagle's medium (Gibco
Life Technologies, Carlsbad, CA, USA) supplemented with 10% fetal
bovine serum (Gibco Life Technologies), 100 U/ml penicillin and 100
µg/ml streptomycin, at 37°C in an incubator with 5%
CO2.
Cell viability assay
A 3-(4,5-dimethylthiazol-2-yl)-2,5 diphenyl
tetrazolium bromide (MTT) assay was performed to assess the
viability of the cells. Briefly, the CNE-1 or NRK-52E cells were
seeded in 96-well plates at a density of 1×105 cells/ml
and treated with various concentrations of plant extracts (12.5,
25, 50, 100 and 200 µg/ml) at 37°C for 72 h. The cells were
also treated with the known anti-cancer drug, 5-fluorouracil
(Sigma-Aldrich) at the concentrations of 12.5, 25, 50, 100 and 200
µg/ml. The cytotoxicity of the extracts of NPC cells was
compared with the effect of 5-fluorouracil. MTT (10 µl;
Biobasic, Amherst, NY, USA) was then added to each well and
incubated for an additional 4 h prior to removal of the media.
Dimethyl sulfoxide (Friendemann Schmidt, Parkwood, WA, USA) was
subsequently added to dissolve the formazan product. The absorbance
was read at 570 nm wavelength, with the results expressed as a
percentage of cell viability using the following formula: Optical
density (OD) sample/OD control × 100%. A dose-response curve was
plotted, and the half maximal inhibitory concentration
(IC50) of the extracts were determined through
interpolation from the curve. The extracts that exhibited cytotoxic
activity were used in the following assays for the determination of
cell proliferation and apoptosis. The control cells in the present
study represent untreated cells.
The selectivity index (SI) was calculated from the
IC50 ratio of NRK-52E and CNE-1 cells. The SI value
indicates the selectivity of the sample to the cell lines assessed.
Samples with an SI value >3 were considered to have high
selectivity (21).
Morphological assessment of cells
The CNE-1 cells (1×106 cells per well of
6-well plate) were treated with various extracts at their
IC50 concentration for 72 h at 37°C. Morphological
changes of the cells were observed under an inverted microscope
(Nikon Eclipse TS100; Nikon Corporation, Tokyo, Japan) and
micrographs were captured.
Determination of population doubling time
(PDT)
The PDT was calculated to estimate the duration of
the cell cycle and compare the effects of the extracts. The cells
were counted using a manual hemocytometer (Marienfeld,
Lunda-Königshofen, Germany). Briefly, 1×104 CNE-1 cells
were seeded into a 24-well plate and treated with the various
extracts at their IC50 concentration for 72 h at 37°C.
Subsequently, the number of cells were counted following staining
with trypan blue (Gibco Life Technologies). The PDT was calculated
by dividing the total duration (h) by the total number of
generations. The number of generations was calculated by 3.32
(logNN-logN1), where NN is the
number of cells counted and N1 is the number of cells
inoculated (22).
Flow cytometric analysis
The cells (1×106 cells per well of 6-well
plate) were treated with the plant extracts at their respective
IC50 concentrations for 72 h at 37°C were harvested and
fixed in 70% ethanol. Following two washes with cold
phosphate-buffered saline, 500 µl propidium iodide (20
µg/ml; Sigma-Aldrich, St. Louis, MO, USA) and 500 µl
RNase (Sigma-Aldrich) was added to the cells. Flow cytometric
analysis (FACScan; BD Biosciences, Franklin Lakes, NJ, USA) was
performed following 30 min incubation, in order to investigate the
rate of cell apoptosis.
Caspase activity assay
The Apo-ONE® Homogeneous Caspase-3/7
Assay kit was purchased from Promega Corporation (Madison, WI, USA)
and the Caspase-8 and-9 Assay kits were obtained from Calbiochem
(Merck KGaA, Darmstadt, Germany). The CNE-1 cells were treated with
the different plant extracts at their IC50
concentrations for 72 h, following which the activities of caspase
3/7, -8 and -9 were measured using the above-mentioned kits. The
detection of caspase activity was performed, according to the
manufacturer's instructions.
Statistical analysis
The results are presented as the mean ± standard
deviation. The data were subjected to one-way analysis of variance
using GraphPad Instat version 3.0 (GraphPad Software, Inc., La
Jolla, CA, USA). P<0.05 was considered to indicate a
statistically significant difference.
Results
Cytotoxicity of S. crispa extracts
A summary of the IC50 of the extracts on
CNE-1 cells is presented in Table
I. Of all the extracts assessed, LH, LC, LEA, SH, SC and SEA
resulted in CNE-1 cell death. The hexane extract of S.
crispa stems exhibited the most potent cytotoxic effect against
the CNE-1 cells, with an IC50 value as low as 49.4±8.0
µg/ml. Following the hexane extract, the order of
cytotoxicity was as follows: LEA>LH> SC>LC>SEA extracts
of S. crispa. Methanol and water extracts of the S.
crispa leaves and stems caused no significant inhibition of
cell growth of CNE-1 cells.
 | Table IIC50 (µg/ml) and
selectivity index values of extracts of Strobilanthes crispa
on CNE-1 and NRK-52E cells. |
Table I
IC50 (µg/ml) and
selectivity index values of extracts of Strobilanthes crispa
on CNE-1 and NRK-52E cells.
| Plant material |
Extract/treatment | IC50
(µg/ml)
| Selectivity
index |
|---|
| CNE-1 | NRK-52E |
|---|
| Leaves | Hexane | 123.50±37.50 | 84.00±1.41 | 0.68 |
| Chloroform | 161.70±20.20 | 184.50±12.02 | 1.14 |
| Ethyl acetate | 119.00±48.10 | 166.50±2.12 | 1.40 |
| Methanol | N/A | N/A | – |
| Water | N/A | N/A | – |
| Stems | Hexane | 49.40±8.00 | 11.00±2.83 | 0.22 |
| Chloroform | 148.30±23.20 | N/A | >1.35 |
| Ethyl acetate | 163.50±16.30 | 174.00±5.66 | 1.06 |
| Methanol | N/A | N/A | – |
| Water | N/A | N/A | – |
| – | 5-fluorouracil | 3.05±1.15 | 9.60±4.81 | 3.15 |
The cytotoxicity of the extracts on NRK-52E cells
was also assessed. Similar observations were noted in the NRK-52E
cells as the CNE-1 cells. SH exhibited the most potent activity
against the NRK-52E cells, with the lowest IC50 value.
However, it was not possible to determine the IC50 value
of SC. Overall, all of the extracts in the present study exhibited
a low SI, with values ranging between 0.22 and 1.40.
The anticancer drug 5-fluorouracil exhibited potent
activity against CNE-1 cell growth, with a low IC50
value (3.05±1.15 µg/ml) and high SI (3.15).
Antiproliferative effects of S. crispa
extracts
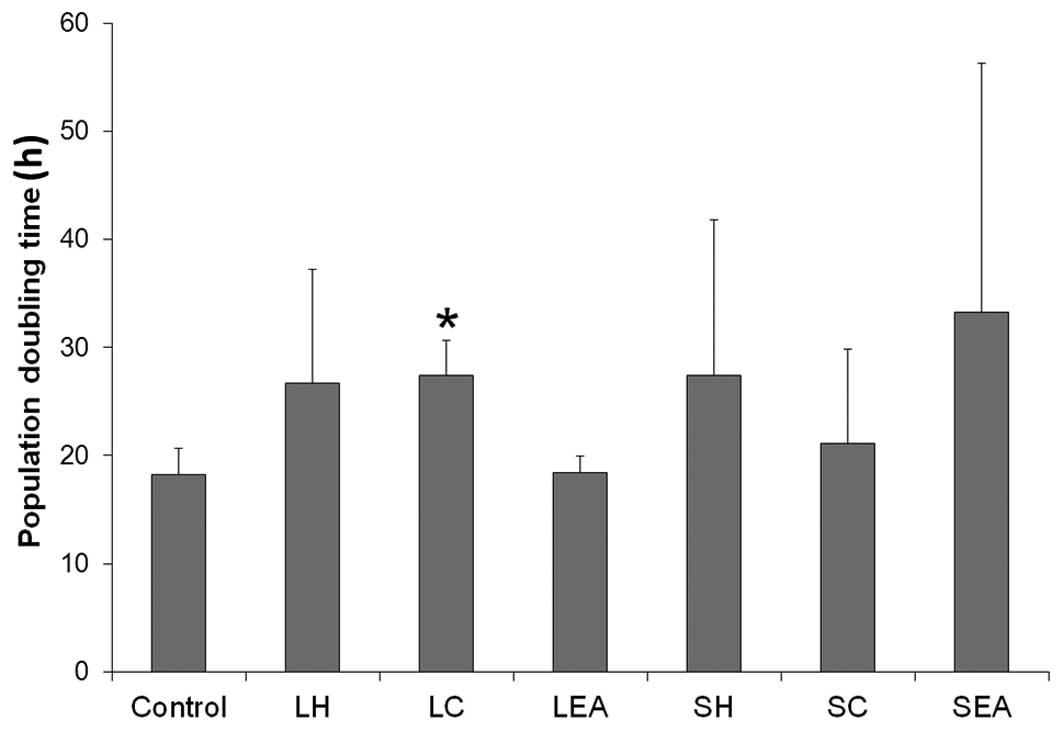
As shown in Fig. 1,
all the plant extracts prolonged the PDT of the cells, with the
exception of LEA. Treatment with SC resulted in marginal increase
in PDT, compared with the control cells, whereas the cells treated
with LH, LC and SH exhibited a ~2-fold increase in PDT. Treatment
with SEA increased PDT by 82.50%; however, of the extracts
assessed, only LC increased the PDT with statistical significance
(P<0.05).
Effects of S. crispa on CNE-1 cell
morphology
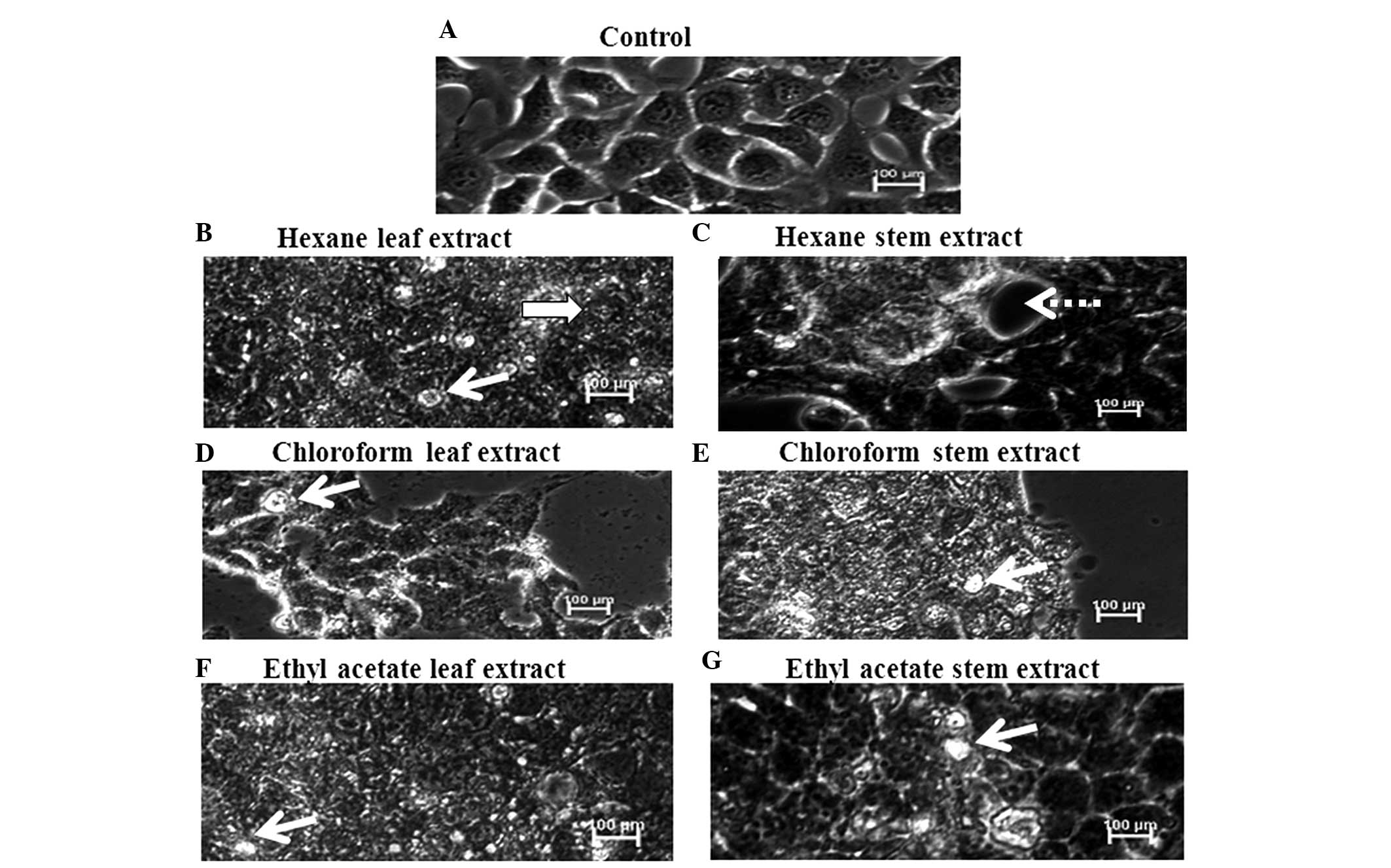
The normal morphology of CNE-1 cells is shown in
Fig. 2A. The cells appeared
attached among each other and well-organized. The cells treated
with the various plant extracts (Figs.
2B–G) exhibited distinct morphological changes, compared with
the control cells; with apoptotic bodies and floating dead cells.
In addition, a reduction in the number of cells was observed in the
extract-treated wells.
Apoptogenic effects of S. crispa on CNE-1
cells
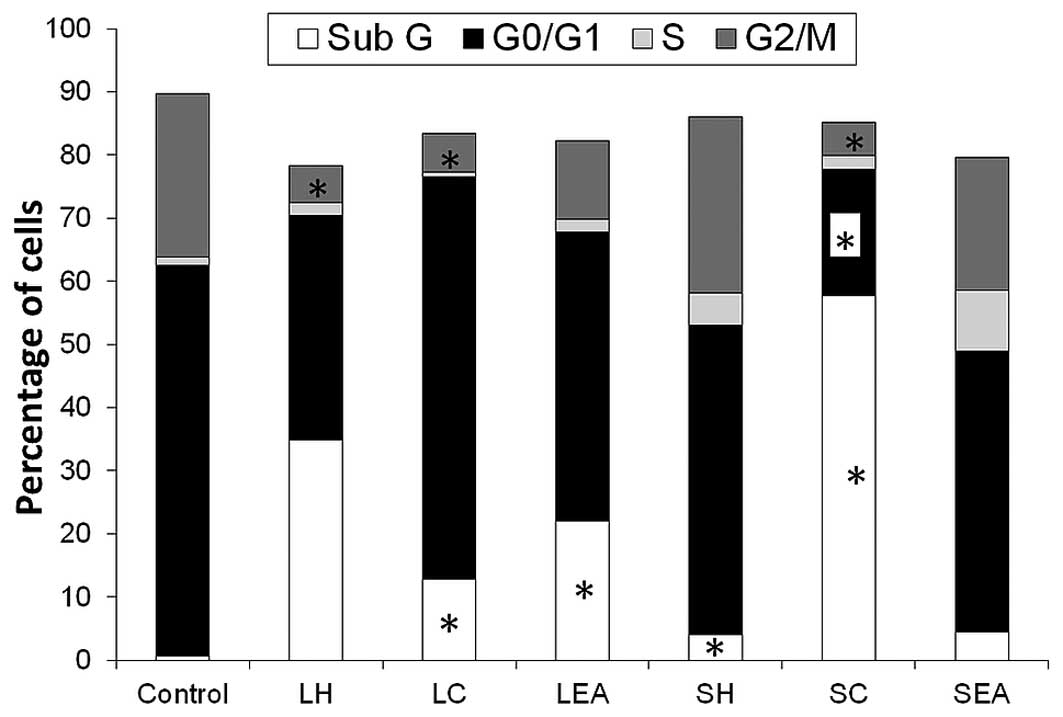
As shown in Fig. 3,
the percentage of control cells in the sub G phase (0.69%)
increased significantly to 12.85 and 22.08% following treatment
with LC and LEA, respectively (P<0.05). The percentage of cells
in the sub G phase was also markedly increased by SC (57.67%) and
moderately increased by SH (4.07%; P<0.05). Treatment with SC
also reduced the percentage of cells in the
G0/G1 phase by 67.55%. A significant
reduction in the number of cells in the G2/M phase was
also observed following treatment of the cells with LH, LC and SC
(P<0.05).
Effects of S. crispa on caspase
activation
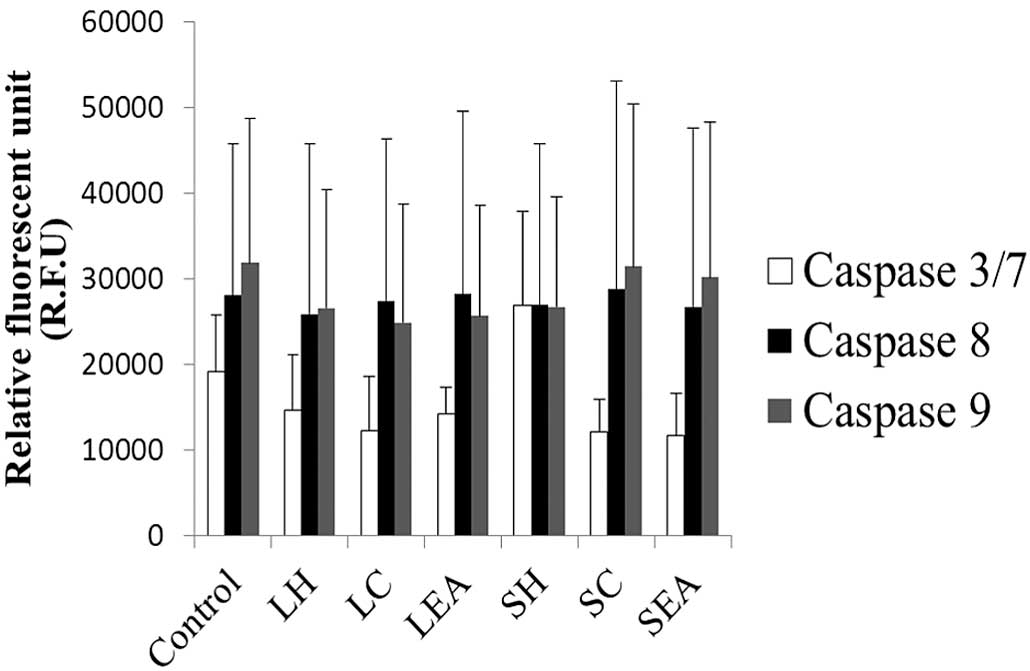
As shown in Fig. 4,
treatment with all the extracts, with the exception of SH,
decreased the levels of caspase 3/7 in the CNE-1 cells. However, no
marked changes were observed in the levels of caspase-8 and -9.
None of these differences observed in caspase activities were
statistically significant.
Discussion
The principle behind the MTT assay is the reduction
of yellow tetrazolium salt to purple formazan by reductase enzyme
of viable cells, and this conversion directly reflects the number
of viable cells. Therefore, an MTT assay was used in the present
study to determine the cytotoxicity of plant extracts of S.
crispa on CNE-1 NPC cells.
The IC50 values of the S. crispa
extracts on CNE-1 cells are summarized in Table I. The cytotoxic effects exhibited
by S. crispa extracts in the present study were concordant
with the findings of a previous study by Rahmat et al
(23). The previous study
demonstrated that bioactive compounds isolated from S.
crispa leaves exhibit anticancer properties towards a certain
cancer cell lines. The bioactive compounds were identified as
β-sitosterol and stigmasterol (23). However, there may be other
bioactive compounds that are able to inhibit cancer growth in S.
crispa, requiring further investigation of the isolation and
purification of the active compounds. Liza et al (24) observed the presence of flavonoids,
(+)-catechin, (−)-epicatechin, rutin, myricetin, luteolin,
apigenin, naringenin and kaempferol, in S. crispa. The
consumption of vegetables and fruits rich in flavonoids has
previously been associated with a decreased cancer risk (25,26);
therefore, these flavonoids may also contribute to the anticancer
properties of S. crispa.
In the present study, the hexane extract of S.
crispa stems exhibited the most potent cytotoxic effect against
the CNE-1 cells. These results suggested that the active compounds
in S. crispa are of low polarity, as cytotoxic effects were
detected in samples extracted with solvents of low polarity,
including hexane. Hexane has been identified as a suitable solvent
for the extraction of non-polar compounds, including oils and fats.
A previous study successfully isolated anthraquinones from the
roots of Cratoxylum formasum as a potent anticancer agent
using hexane (27). In the present
study, the IC50 value of SH extract was markedly lower
than that of the LH extract (Table
I). This many indicate that the active compounds responsible
for the anticancer property of S. crispa are more abundant
in the stems, compared with the leaves.
Methanol and water extracts from S. crispa
leaves and stems did not cause CNE-1 cell death. This result was
consistent with that of a previous study, which demonstrated that
the water extract of S. crispa did not exhibit a cytotoxic
effect toward any of the cancer cell lines assessed (13). These results suggested that the
polar compounds extracted by methanol or water had a less toxic
effect against cancer cells.
Previous studies have indicated that crude extracts
of S. crispa have no cytotoxic effect against Chang liver
cells (12). In the present study,
the cytotoxicity of S. crispa was assessed on NRK-52E normal
kidney epithelial cells, since kidneys serve as the excretory organ
of metabolized drugs. The results of the present study demonstrated
that the extracts that exhibited a cytotoxic effect towards the
CNE-1 cells also inhibited proliferation of the NRK-52E cells, with
the exception of SC. The IC50 value of SC on NRK-52E
cells was not determined, even at the highest concentration (200
µg/ml) of the plant extract. Therefore, SC had a higher SI
index (>1.35), compared with the other extracts, suggesting that
SC is safer to use as an anticancer agent.
PDT is the average duration required for successful
cell division, and is used an indicator of cell proliferation. As
shown in Fig. 1, treatment with LC
significantly increased the PDT. These results suggested that the
effect of LC on CNE-1 cell growth inhibition, detected by the MTT
assay, may be due to the ability of LC to decrease the rate of cell
proliferation.
The stimulation of apoptosis and inhibition of cell
proliferation remain the most effective anticancer treatment
strategies; therefore, the therapeutic induction of apoptosis has
been used as a model for the development of antitumor drugs
(28). In the present study,
apoptotic features of cells were observed under the microscope.
Quantitative measurement of apoptosis was performed by propidium
iodide staining of DNA, analyzed using flow cytometry. The flow
cytometric assay aimed to detect cells in the sub-G1
phase, resulting from DNA fragmentation (29).
As shown in Figs.
2B–G, CNE-1 cell morphology was markedly altered following
treatment with the S. crispa extracts. Apoptotic features,
as well as dead cells, were observed. The cells were subsequently
subjected to cell cycle distribution analysis using flow cytometry.
The results demonstrated that the percentage of cells in the
sub-G1 phase increased, and the percentage of cells in
the S and G2/M phases decreased following treatment with
the S. crispa extracts. These results indicated that the
extracts induced apoptosis and fewer CNE-1 cells entered into the
synthesis phase and mitotic cycle. Reduced proliferation of the
CNE-1 cells was noted in the present study, following treatment
with the S. crispa extracts. There was a positive correlation
between the inhibition of cell proliferation and inhibition of cell
cycle progression, due to the increased proportion of cells in the
sub-G1 phase, together with the decreased proportion of
cells in the G2/M phase (30). Furthermore, distinctive
morphological changes associated with apoptosis, including
apoptotic body formation (31),
were prevalent in the extract-treated CNE-1 cells. These results
suggested that S. crispa may induce CNE-1 cell death through
the apoptotic mechanism. However, further assessments for
apoptosis, including annexin-V staining, a comet assay and a
terminal deoxynucleotidyl transferase dUTP nick-end labeling assay
are required in the future to confirm these results. In addition,
investigation of the apoptosis-mediated mechanisms is required for
better understanding of the effects of S. crispa at the
molecular level, therefore the present study detected caspase
activation.
Caspases belong to a group of enzymes known as
cysteine proteases, which act as important mediators of signal
transduction in apoptosis. Caspase-8 and -9 are regarded as two key
initiators of apoptosis, the death receptor-mediated and
mitochondrion-mediated pathways. By contrast, caspase-3/7 are
responsible as effectors in executing apoptosis by cleaving key
cellular substrates, thus leading to apoptotic cell death (32). As shown in Fig. 4, no significant changes in caspase
levels were observed in the CNE-1 cells treated with the S.
crispa extracts, compared with the untreated cells. These
results suggested that caspase-mediated apoptosis may not be the
sole process of apoptotic cell death induced by S. crispa.
The other possible mechanism of action of apoptosis may be via
downregulation of the expression of c-myc, lysosomal proteases
(33) or granzyme release
(34), which require further
investigation. Several studies have suggested that downregulation
of the expression of c-myc may be necessary for the induction of
apoptosis in leukemia cells (35),
macrophage cells (36), prostate
cancer (37) and lung cancer
(38). Notably, a previous study
demonstrated that chloroform extract of S. crispa suppressed
the expression of c-myc expression in cancer (18), suggesting the important role of
c-myc in mediating the effects of S. crispa.
In conclusion, hexane, chloroform and ethyl acetate
extracts of S. crispa leaves and stems exerted potent
cytotoxic effects against CNE-1 cells. The extracts also induced
apoptosis by increasing the sub-G1 population, and the
effects were independent of caspase-3, -7 and -9 activation. Based
on the observations of the present study, these extracts may serve
as potential anticancer agents. Therefore, isolation of active
compounds from S. crispa extracts and evaluation of their
anti-cancer effects in vivo may be performed in the
future.
Acknowledgments
This study was supported by the International
Medical University (Kuala Lumpur, Malaysia; no. BPharm B0108_
Res082011).
References
|
1
|
Wolf H, zur Hausen H and Becker V: EB
viral genomes in epithelial nasopharyngeal carcinoma cells. Nat New
Biol. 244:245–247. 1973. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lo YM, Chan LY, Lo KW, Leung SF, Zhang J,
Chan AT, Lee JC, Hjelm NM, Johnson PJ and Huang DP: Quantitative
analysis of cell-free Epstein-Barr virus DNA in plasma of patients
with nasopharyngeal carcinoma. Cancer Res. 59:1188–1191.
1999.PubMed/NCBI
|
|
3
|
Neel HB III, Pearson GR and Taylor WF:
Antibodies to Epstein-Barr virus in patients with nasopharyngeal
carcinoma and in comparison groups. Ann Otol Rhinol Laryngol.
93:477–482. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Tabuchi K, Nakayama M, Nishimura B,
Hayashi K and Hara A: Early detection of nasopharyngeal carcinoma.
Int J Otolaryngol. 2011:6380582011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Australian State of the Environment
Committee 2001, Australia State of the Environment 2001,
Independent Report to the Commonwealth Minister for the Environment
and Heritage. CSIRO Publishing on behalf of the Department of the
Environment and Heritage, Canberra.
|
|
6
|
Gray AM and Flatt PR: Actions of the
traditional anti-diabetic plant, Agrimony eupatoria (agrimony):
Effects on hyperglycaemia, cellular glucose metabolism and insulin
secretion. Br J Nutr. 80:109–114. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gohil KJ, Patel JA and Gajjar AK:
Pharmacological review on Centella asiatica: A potential herbal
cure-all. Indian J Pharm Sci. 72:546–556. 2010. View Article : Google Scholar
|
|
8
|
Burkill IH, Birtwistle W, Foxworthy FW,
Scrivenor JB and Watson JG: A Dictionary of the Economic Products
of the Malay Peninsula. 2. (Crown Agents for the Colonies). London:
pp. 2086–2087. 1935
|
|
9
|
Perry LM and Metzger J: Medicinal plants
of east and south east Asia. Attributed properties and uses. MIT
press; Cambridge, MA: pp. 452–617. 1980
|
|
10
|
Samuel AJ, Kalusalingam A, Chellappan DK,
Gopinath R, Radhamani S, Husain HA, Muruganandham V and Promwichit
P: Ethnomedical survey of plants used by the Orang Asli in Kampung
Bawong, Perak, West Malaysia. J Ethnobiol Ethnomed. 6:52010.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kusumoto JT, Shimada I, Kakiuchi N,
Hattori M, Supriyatna S and Namba T: Inhibitory effects of
Indonesian plant extracts on reverse transcriptase of an RNA tumour
virus (I). Phytother Res. 6:241–244. 1992. View Article : Google Scholar
|
|
12
|
Yaacob NS, Hamzah N, Nik Mohamed Kamal NN,
Zainal Abidin SA, Lai CS, Navaratnam V and Norazmi MN: Anticancer
activity of a sub-fraction of dichloromethane extract of
Strobilanthes crispus on human breast and prostate cancer cells in
vitro. BMC Complement Altern Med. 10:422010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Muslim NS, Ng KW, Itam A, Nassa ZD, Ismail
Z and Abdul Majid AMS: Evaluation of cytotoxic, anti-angiogenic and
antioxidant properties of standardized extracts of Strobilanthes
crispus leaves. International Journal of Pharmacology. 6:591–599.
2010. View Article : Google Scholar
|
|
14
|
Rahmat A, Endrini S, Md Akim A, Ismail P,
Taufiq Y, Yun H and Abu Bakar MF: Anticarcinogenic properties of
Strobilanthes crispus extracts and its compounds in vitro. Int J
Cancer Res. 2:47–49. 2006. View Article : Google Scholar
|
|
15
|
Chong HZ, Rahmat A, Yeap SK, Md Akim A,
Alitheen NB, Othman F and Gwendoline-Ee CL: In vitro cytotoxicity
of Strobilanthes crispus ethanol extract on hormone dependent human
breast adenocarcinoma MCF-7 cell. BMC Complement Altern Med.
12:352012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Endrini S, Rahmat S, Ismail P and
Taufiq-Yap YH: Comparing of the cytotoxicity properties and
mechanism of Lausonia inermis and Strobilanthes crispus extract
against several cancer cell lines. J Med Sci. 7:1098–1102. 2007.
View Article : Google Scholar
|
|
17
|
Ng KW, Salhimi SM, Majid AM and Chan KL:
Anti-angiogenic and cytotoxicity studies of some medicinal plants.
Planta Med. 76:935–940. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Endrini S, Suherman, Rahmat A, Ismail P,
Taufiq-Yap YH and Othman F: Effects of Strobilanthes crispus
extract on the apoptotic pathway of human liver carcinoma cell
lines. Jurnal Kedorkteran Yarsi. 15:1–5. 2007.
|
|
19
|
Hanachi P, Othman F and Rahmat A: Lesion
scoring and P450 isoenzyme activity in liver of
hepatocarcinogenesis rats treated with Strobilanthes crispus.
Iranian Journal of Cancer Prevention. 1:12–16. 2008.
|
|
20
|
Suherman J, Rahmat A, Othman F, Ismail P
and Nor Haslinda A: Effect of Strobilanthes crispus on tumour
marker enzymes and glutathione during chemical hepatocarcinogenesis
in the rat. Pakistan J Biol Sci. 7:947–951. 2004. View Article : Google Scholar
|
|
21
|
Mahavorasirikul W, Viyanant V,
Chaijaroenkul W, Itharat A and Na-Bangchang K: Cytotoxic activity
of Thai medicinal plants against human cholangiocarcinoma,
laryngeal and hepatocarcinoma cells in vitro. BMC Complement Altern
Med. 10:552010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Schoene NW and Kamara KS: Population
doubling time, phosphatase activity, and hydrogen peroxide
generation in Jurkat cells. Free Radic Biol Med. 27:364–369. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Rahmat A, Edrini S, Ismail P, Taufiq Y,
Yun H and Abu Bakar MF: Chemical constituents, antioxidant activity
and cytotoxic effects of essential oil from Strobilanthes crispus
and Lawsonia inermis. J Biol Sci. 6:1005–1010. 2006. View Article : Google Scholar
|
|
24
|
Liza MS, Abdul Rahman R, Mandana B, Jinap
S, Rahmat A, Zaidul ISM and Hamid A: Supercritical carbon dioxide
extraction of bioactive flavonoid from Strobilanthes crispus (Pecah
Kaca). Food Bioprod Process. 88:319–326. 2010. View Article : Google Scholar
|
|
25
|
Ferguson PJ, Kurowska E, Freeman DJ,
Chambers AF and Koropatnick DJ: A flavonoid fraction from cranberry
extract inhibits proliferation of human tumor cell lines. J Nutr.
134:1529–1535. 2004.PubMed/NCBI
|
|
26
|
Jain R and Jain SK: Screening of in vitro
cytotoxic activity of some medicinal plants used traditionally to
treat cancer in Chhattisgarh state, India. Asian Pac J Trop Biomed.
S147–S150. 2011. View Article : Google Scholar
|
|
27
|
Boonsri S, Karalai C, Ponglimanont C,
Kanjana-opas A and Chantrapromma K: Antibacterial and cytotoxic
xanthones from the roots of Cratoxylum formosum. Phytochemistry.
67:723–727. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang TS, Chen LJ, Zhang ST, Lin JM and
Wang ZY: Purified alkaloids extract from Scutellaria barbata
inhibits proliferation of nasopharyngeal carcinoma CNE-1 cells by
inducing apoptosis and cell cycle arrest at S phase. J Med Plants
Res. 5:3687–3696. 2011.
|
|
29
|
Tayarani-Najaran Z, Emami SA, Asili J,
Mirzaei A and Mousavi SH: Analyzing cytotoxic and apoptogenic
properties of Scutellaria litwinowii root extract on cancer cell
lines. Evid Complementary Alternat Med. 2011:1606822011.
|
|
30
|
Yi ZC, Wang H, Zhang GY and Xia B:
Downregulation of connexin 43 in nasopharyngeal carcinoma cells is
related to promoter methylation. Oral Oncol. 43:898–904. 2006.
View Article : Google Scholar
|
|
31
|
Hu ZY, Zhu XF, Zhong ZD, Sun J, Wang J,
Yang D and Zeng YX: ApoG2, a novel inhibitor of antiapoptotic Bcl-2
family proteins, induces apoptosis and suppresses tumor growth in
nasopharyngeal carcinoma xenografts. Int J Cancer. 123:2418–2429.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Riedl SJ and Shi Y: Molecular mechanisms
of caspase regulation during apoptosis. Nat Rev Mol Cell Biol.
5:897–907. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Stoka V, Turk B, Schendel SL, Kim TH,
Cirman T, Snipas SJ, Ellerby LM, Bredesen D, Freeze H, Abrahamson
M, et al: Lysosomal protease pathways to apoptosis: Cleavage of
bid, not pro-caspases, is the most likely route. J Biol Chem.
276:3149–3157. 2001. View Article : Google Scholar
|
|
34
|
Barry M, Heibein JA, Pinkoski MJ, Lee SF,
Moyer RW, Green DR and Bleackley RC: Granzyme B short-circuits the
need for caspase 8 activity during granule-mediated cytotoxic
T-lymphocyte killing by directly cleaving Bid. Mol Cell Biol.
20:3781–3794. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Alneri ES, Fernandes TF, Halder S, Croce
CM and Litwack G: Involvement of BCL-2 in glucocorticoid-induced
apoptosis of human pre-B-leukemias. Cancer Res. 52:491–495.
1992.
|
|
36
|
Oritani K, Kaisho T, Nakajima K and Hirano
T: Retinoic acid inhibits interleukin-6-induced macrophage
differentiation and apoptosis in a murine hematopoietic cell line,
Y6. Blood. 80:2298–2305. 1992.PubMed/NCBI
|
|
37
|
Balaji KC, Koul H, Mitra S, Maramag C,
Reddy P, Menon M, Malhotra RK and Laxmanan S: Antiproliferative
effects of c-myc antisense oligonucleoride in prostate cancer
cells: A novel therapy in prostate cancer. Urology. 50:1007–1015.
1997. View Article : Google Scholar
|
|
38
|
Van Waardenburg RC, Meijer C, Burger H,
Nooter K, De Vries EG, Mulder NH and De-Jong S: Effects of an
inducible anti-sense c-myc gene transfer in a drug-resistant human
small-cell-lung-carcinoma cell line. Int J Cancer. 73:544–550.
1997. View Article : Google Scholar : PubMed/NCBI
|