Introduction
Infection with Helicobacter pylori is one of
the most common types of bacterial infection. Helicobacter
pylori infection mainly occurs in economically underdeveloped
regions, with the infection rates of pediatric patients in China,
Japan and Korea being higher than those in developed countries
(1–5). Helicobacter pylori colonizes
the stomach and duodenum areas, causing chronic inflammation of the
gastric mucosa, and the development of stomach ulcers and neoplasms
(gastric cancer and mucosa-associated lymphoid tissue) (6,7).
However, the mechanism by which Helicobacter pylori
infection causes pathological changes of the gastric mucosa remains
to be fully elucidated.
Inflammatory responses have been known to have a key
role in the pathogenesis of Helicobacter pylori infection
and contribute to chronic gastritis as well as gastric and duodenal
ulcers in the patients (8–10). A growing body of evidence has
demonstrated that elevated levels of inflammatory factors,
including tumor necrosis factor (TNF)-α, interferon-γ, interleukin
(IL)-6, IL-8 and IL-32 were associated with more serious
pathogenesis of patients infected with Helicobacter pylori
(10–15). However, the underlying regulatory
mechanisms of the production of inflammatory factors have largely
remained elusive.
Nuclear factor (NF)-κB has an important role in the
regulation of inflammatory responses in mammals (16,17).
It has been demonstrated that Helicobacter pylori infection
activates NF-κB and its target genes, particularly inflammatory
factors in epithelial cells, which is thought to be critical for
Helicobacter pylori-initiated chronic inflammation (9,18–21).
The virulence factor cytotoxin-associated gene A (CagA) encoded by
the Helicobacter pylori Cag pathogenicity island, has an
important role in the pathogenicity of Helicobacter pylori,
including Helicobacter pylori-induced activation of NF-κB
and expression of NF-κB target genes (22,23).
However, the exact function of CagA in the activation of NF-κB and
the NF-κB-dependent inflammatory response has not been well
characterized.
Emerging evidence indicated that post-translational
modification, such as acetylation, has important roles in various
biological events, including inflammation (24–26).
It has been reported that the acetylation of the NF-κB p65 sub-unit
has an important role in its activation (27–29).
In addition, recent studies showed that P300/CBP-associated factor
(PCAF), also frequently referred to as lysine (K) acetyltransferase
2B, is a transcription co-activator that contains several nuclear
receptor-interacting domains and can function as an acetyl
transferase (30–33). PCAF has also been reported to be
associated with infection and inflammation (34–37).
Therefore, there is a requirement to explore the roles of PCAF in
mediating p65 activation and the production of TNF-α and IL-6 in
gastric adenocarcinoma cells.
The present study aimed to investigate the
implication of Helicobacter pylori CagA in the inflammatory
response of human gastric adenocarcinoma AGS cells in vitro,
as well as PCAF-mediated p65 acetylation and its role in regulating
the production of TNF-α and IL-6 as two important inflammatory
factors in AGS cells.
Materials and methods
Reagents
Monoclonal rabbit antibodies against human total p65
(cat. no. 8242), phospho (p-)p65 (cat. no. 3031), as well as
monoclonal mouse antibodies against acetylated-Lysine (cat. no.
9681) and human β-actin (cat. no. 3700) were from Cell Signaling
Technology (Danvers, MA, USA). Mouse monoclonal antibodies against
human PCAF (cat. no. sc-13124) and Hemagglutinin (HA)-tag (cat. no.
sc-7392) were purchased from Santa Cruz Biotechnology (Dallas, TX,
USA). For western blot analysis, horseradish peroxidase-conjugated
anti-mouse immunoglobulin G (IgG) antibody (cat. no. 7076) and
anti-rabbit IgG antibody (cat. no. 7074) were purchased from Cell
Signaling Technology. Enhanced chemiluminescence (ECL) Western
Blotting Substrate, co-immunoprecipitation (co-IP) assay buffer,
radioimmunoprecipitation assay (RIPA) lysis buffer, a bicinchoninic
acid (BCA) protein assay kit and protein G-Sepharose beads were
purchased from Thermo Fisher Scientific (Waltham, MA, USA).
Polyvinylidene difluoride (PVDF) membranes were obtained from Roche
(Basel, Switzerland). TRIzol reagent was purchased from Invitrogen
Life Technologies (Carlsbad, CA, USA). Moloney Murine Leukemia
Virus reverse transcrip-tase and oligo (dT) 15 primer were
purchased from Promega (Madison, WI, USA). TaqMan® Fast
Advanced Master Mix was from Applied Biosystems (Thermo Fisher
Scientific). The plasmids pGCsi-U6/neo/green fluorescence protein
(GFP) were obtained from Shanghai Genkan Biotechnology Co., Ltd
(Shanghai, China). The pcDNA3.1 vector was purchased from
Invitrogen Life Technologies. The incision enzymes HindIII
and XhoI as well as T4 DNA ligase were purchased from Takara
Bio Inc. (Tokyo, Japan). The Escherichia coli strain DH5α
was purchased from Molecular Cloning Laboratories (San Francisco,
CA, USA). QIAprep spin miniprep kit was obtained from Qiagen
(Hilden, Germany). PDTC was purchased from Abcam (Cambridge,
UK).
Cell culture and treatment
The human AGS cells were obtained from the American
Type Culture Collection (Manassas, VA, USA). Cells was cultured in
the medium of RPMI-1640 (Gibco-BRL; Invitrogen Life Technologies)
supplemented with 10% fetal bovine serum (FBS; Gibco-BRL) and
antibiotics (50 U/ml penicillin and 100 μg/ml streptomycin)
at 37°C in a 5%-CO2 incubator. AGS cells were incubated
with Helicobacter pylori CagA protein at 37°C for different
time points. For certain studies, the cells were incubated with 10
μM PDTC at 30 min before treatment with Helicobacter
pylori CagA protein.
Construction of overexpression
plasmids
The open reading frame of human PCAF mRNA (National
Center of Biotechnology Information reference sequence,
NM_003884.4) containing a HA tag was amplified by polymerase chain
reaction (PCR) from cDNA of human AGS cells. The PCR products and
pcDNA3.1 vector were further digested with the two restriction
enzymes of HindIII and XhoI, and then ligated with
each other by using T4 DNA ligase. The recombinant plasmids were
amplified in the Escherichia coli strain DH5α at 37°C for 16
h. Subsequently, the plasmids were extracted by QIAprep spin
miniprep kit according to the manufacturer's instructions. Finally,
the constructed plasmids (pcDNA3.1/PCAF-HA) were sequenced to
confirm the nucleotide sequence by GenScript (Nanjing, China).
Construction of small hairpin (sh)RNA
expression plasmids
Various shRNA sequences targeting human PCAF mRNA
(NM_003884.4) were designed to silence the PCAF gene in human AGS
cells. The various DNA segments for the expression of PCAF shRNA
were synthesized and inserted into the shRNA pGCsi-U6/neo/GFP
plasmids by Shanghai Genkan Biotechnology Co., Ltd. The most
effective shRNA expression plasmid to silence human PCAF gene was
selected to be used in subsequent experiments. The PCAF shRNA
sequence was as follows: AAG ATG GCC GTG TTA TTG GTG and the
Scrambled shRNA sequence was as follows: AAT GAC GGG CTT GTT ATG
GGT.
Cellular transfection
Plasmids were transfected into AGS cells by using
Lipofectamine 2000 according to the manufacturer's instructions.
For transfection, 4 μg plasmid was mixed with 250 μl
serum-free RPMI-1640. At the same time, 10 μl Lipofectamine
2000 was mixed with 250 μl serum-free RPMI-1640. The
plasmids and Lipofectamine 2000 were further incubated with each
other for 20 min at room temperature. Finally, the 500-μl
mixture was added into each well in a 6-well plate containing
4×105 AGS cells. The medium was replaced with serum
containing RPMI-1640 at 5 h after transfection, and the cells were
incubated sequentially.
Production of the CagA protein
Helicobacter pylori CagA-His-tag was
constructed in the pET21a plasmid from Novagen (Madison, WI, USA).
The plasmid was then transformed into BL21(DE3) Singles™ Competent
Cells (Novagen) for protein expression and purification according
to the manufacturer's instructions. The protein was extracted by
B-PER® bacterial protein extraction reagent (Thermo
Fisher Scientific) from 50 ml bacteria according to the
manufacturer's instructions. Subsequently, HisPur™ Cobalt Spin
Columns (Thermo Fisher Scientific) were used for His-tag
purification of the Helicobacter pylori CagA protein
according to the manufacturer's instructions.
co-IP experiment
AGS cells were lysed in co-IP assay buffer at 4°C
for 30 min. Cell lysates were then centrifuged at 15,000 ×g for 20
min at 4°C to remove any insoluble material. 300 μg cell
lysate was incubated with 40 μl protein G-Sepharose beads in
co-IP assay buffer at 4°C for 2.5 h under constant agitation and
then centrifuged at 1,000 × g for 2 min at 4°C. The recovered
supernatant was then incubated with the antibody against human
total p65 (1.5 μg for each sample) at 4°C overnight under
constant agitation. The sample was then incubated with 40 μl
protein G-Sepharose beads at 4°C for 2.5 h under constant
agitation. Protein G-protein complex was precipitated by
centrifugation and finally re-suspended in 40 μl SDS lysis
buffer (Beyotime Institute of Biotechnology, Haimen, China). The
samples were then analyzed by SDS-PAGE followed by staining with
specific antibodies against total p65 (1:1,000), acetylated-Lysine
(1:1,000), PCAF (1:200) and HA (1:500), respectively. At the same
time, 40-μg aliquots of whole-cell extract from the AGS
cells were used to detect the protein expression as an input
control.
RNA isolation and reverse transcription
quantitative (RT-q)PCR
Cells were collected at the indicated time-points
and RNA was extracted using TRIzol reagent. A total of 1 μg
RNA was used for the first-strand cDNA synthesis using MMLV and
oligo (dT) 15 primer according to the manufacturer's instructions.
The cDNA was amplified by using TaqMan® Fast Advanced
Master Mix to detect the expression of human PCAF gene using
primers purchased from GenScript (forward 5′-TAC CTC GGT ACG AAA
CCACA-3′ and reverse 5′-TCC TGT CTT GCT TGT TCCAG-3′;
Fam/Tamra-labeled probe, 5′-CGA GCG AAG CAA TGT TCT CCCA-3′). The
human β-actin gene was used as an internal control using primers
purchased from GenScript (forward 5′-TGG ACT TCG AGC AAG AGATG-3′
and reverse 5′-GAA GGA AGG CTG GAA GAGTG-3′; Fam/Tamra-labeled
probe, 5′-CGG CTG CTT CCA GCT CCTCC-3′). The 7500 Real-time PCR
system (Applied Biosystems) was used to perform qPCR. The reaction
program included an initial step for denaturation at 95°C for 10
min, and 40 cycles of denaturation at 95°C for 15 sec and annealing
at 60°C for 60 sec. Each sample was assayed in triplicate. The
relative levels of the gene expression were obtained using the
2−ΔΔCt method as described previously (38).
Western blot analysis
Cells were lysed in RIPA lysis buffer at 4°C for 30
min. The cellular lysates were then centrifuged at 15,000 x g for
20 min at 4°C to remove any insoluble material. The concentration
of the protein was determined by a BCA protein assay kit according
to the manufacturer's instructions. The protein (40 μg/well)
was subjected to 10–12% SDS-PAGE and transferred onto PVDF
membranes using the Mini-Protean System (Bio-Rad Laboratories,
Inc., Hercules, CA USA). The PVDF membranes were incubated in
blocking buffer [5% skimmed milk in Tris-buffered saline containing
Tween 20 (TBS-T)] at room temperature for 1 h and then incubated
with the specific antibodies (anti-total p65, 1:1,000; anti-p-p65,
1:1,000; anti-acetylated-Lysine, 1:1,000; anti-PCAF, 1:200;
anti-HA; 1:500) at 4°C overnight. β-actin expression in each sample
was used as an internal standard (anti-β-actin, 1:1,000). After
five washes with TBS-T, the PVDF membranes were further incubated
with horseradish peroxidase-conjugated anti-mouse IgG (1:2,000) or
anti-rabbit IgG (1:2,000) at 37°C for 1 h. The bands were
visualized using X-ray film (Life Technologies, Carlsbad, CA, USA)
and the ECL detection system after washing the PVDF membranes five
times. Finally, the density of the radiographic bands on the PVDF
membranes was analyzed using the Quantity One software v44 (Bio-Rad
Laboratories, Inc.).
ELISA
The levels of TNF-α and IL-6 in the cell media were
determined using commercial human TNF-α and IL-6 ELISA kits (cat.
no. 555212 and 550799) from BD Biosciences (Franklin Lakes, NJ,
USA). ELISA was performed according to the manufacturer's
instructions.
Statistical analysis
All statistical analyses were performed using SPSS
12 software (SPSS, Inc., Chicago, IL, USA). Values are expressed as
the mean ± standard deviation. The statistical significance of
differences between groups was evaluated by one-way analysis of
variance with simultaneous multiple comparisons between groups by
the Bonferroni method. P<0.05 was considered to indicate
statistically significant differences.
Results
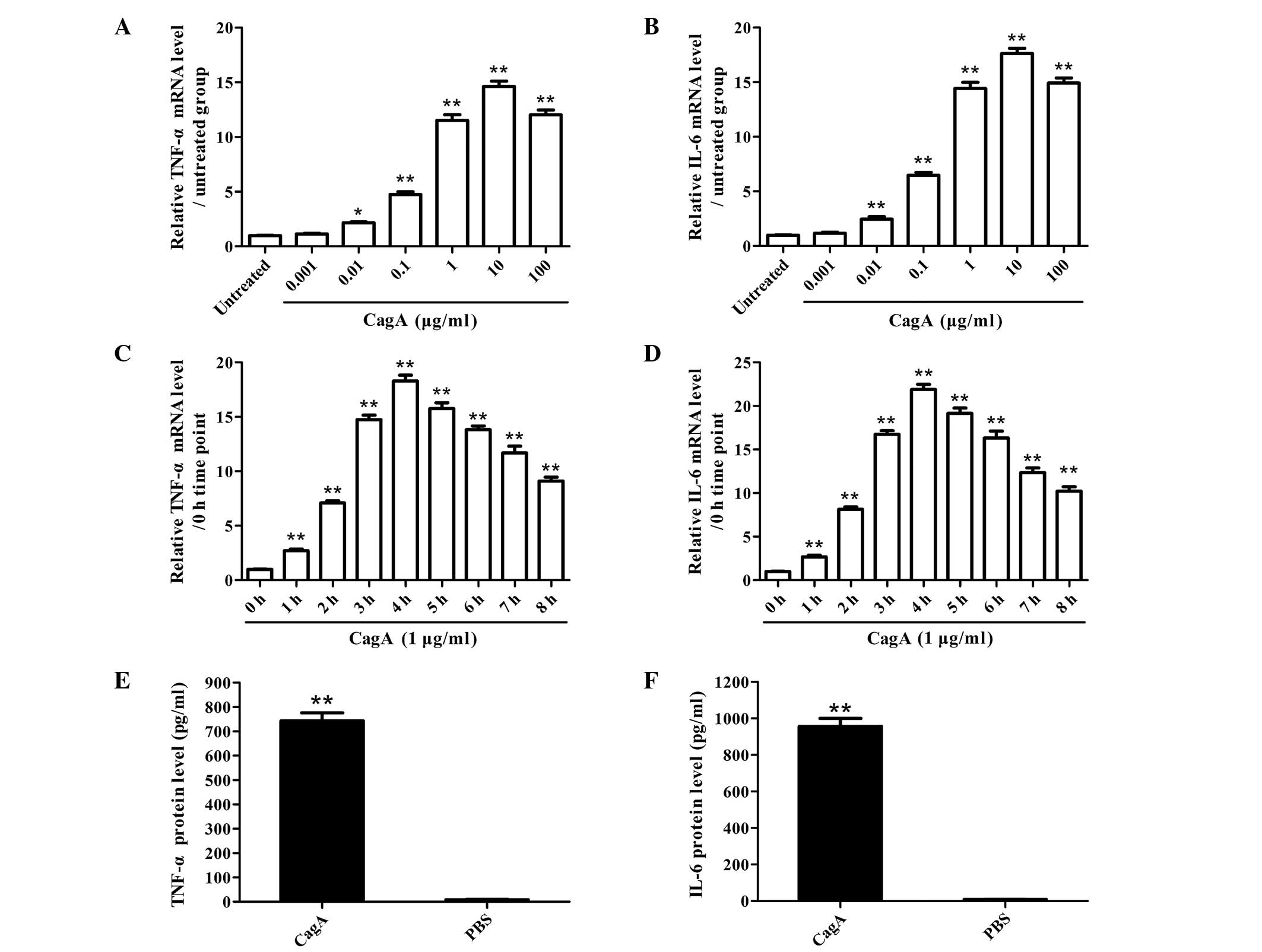
CagA promotes the activation of NF-κB and
the production of TNF-α and IL-6 in human AGS cells
Human AGS cells were cultured with purified
Helicobacter pylori CagA and the production of TNF-α and
IL-6 was subsequently observed. The results showed that CagA
enhanced the production of TNF-α and IL-6 in human AGS cells in
vitro in a dose- and time-dependent manner (Fig. 1). Furthermore, CagA was able to
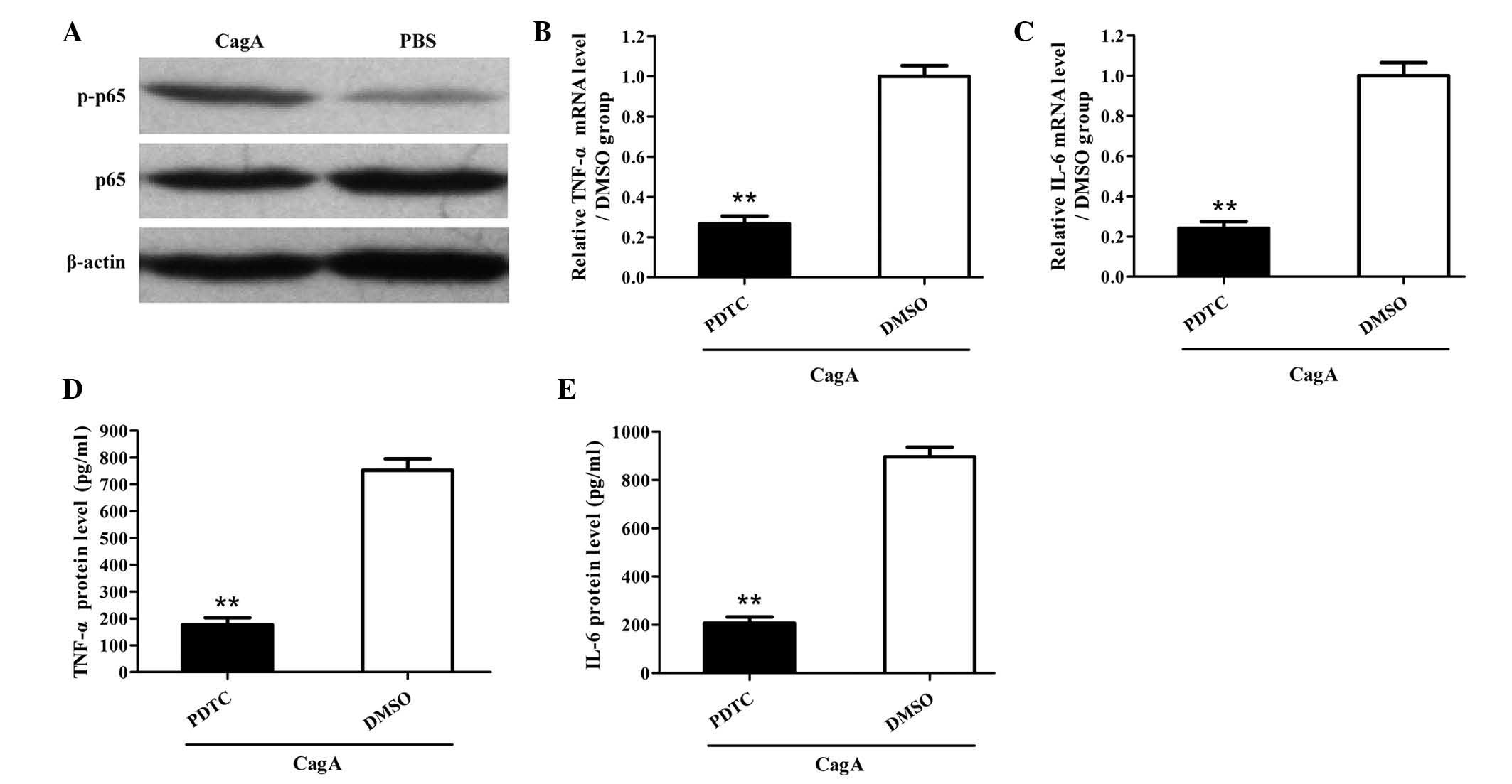
induce the activation of NF-κB in human AGS cells in vitro
(Fig. 2A). Of note, inhibition of
NF-κB significantly reduced the production of TNF-α and IL-6 in AGS
cells induced by CagA (Fig. 2B-E).
These findings indicated that CagA promoted the production of TNF-α
and IL-6 by human AGS cells in vitro via activation of
NF-κB.
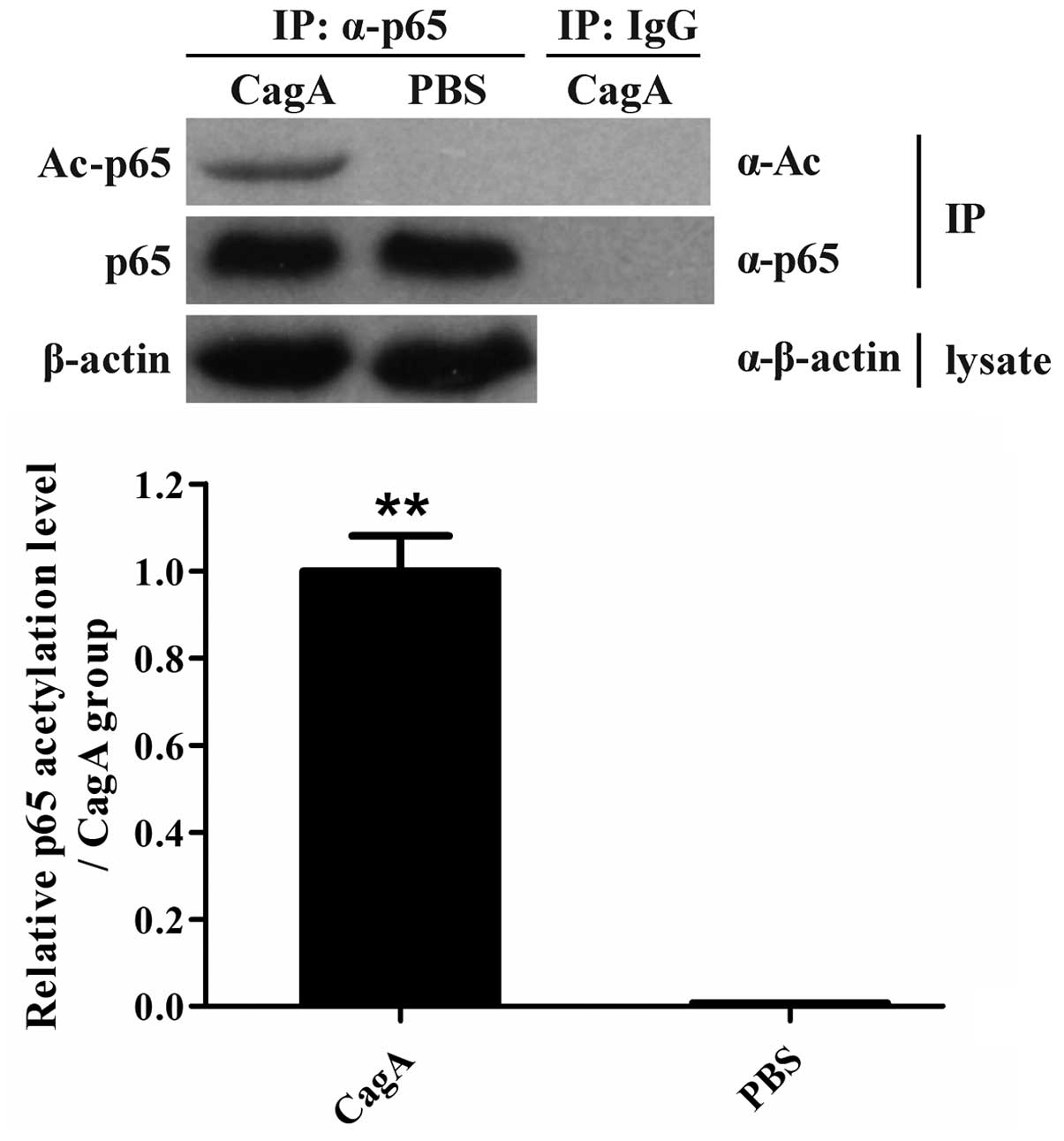
CagA enhances p65 acetylation in human
AGS cells
Since Helicobacter pylori CagA was found to
have the ability to stimulate the activation of NF-κB and the
production of TNF-α and IL-6 in human AGS cells (Figs. 1 and 2), the effects of CagA stimulation on the
acetylation of p65 were further assessed in AGS cells. The level of
p65 acetylation was found to be significantly enhanced in human AGS
cells exposed to CagA in vitro (Fig. 3), suggesting that p65 acetylation
may have an important role in promoting p65 activation in human AGS
cells induced by CagA.
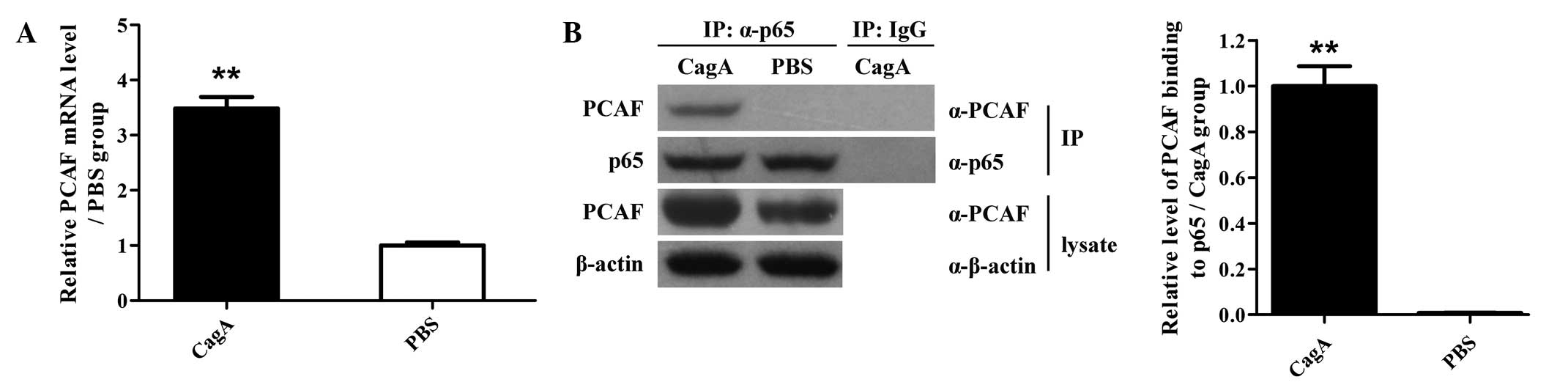
PCAF and p65 association is increased in
human AGS cells exposed to CagA
The expression of PCAF at the mRNA and protein level
was detected in human AGS cells induced by CagA. It was found that
incubation with CagA significantly increased the mRNA and protein
expression levels of PCAF in human AGS cells (Fig. 4A and B). Furthermore, the
interaction of PCAF with p65 was assessed at the protein level by
IP. The results showed that the molecular interaction of PCAF with
p65 at the protein level was markedly enhanced in human AGS cells
upon stimulation with CagA (Fig.
4B). These findings indicated that stimulation with CagA
induced the expression of PCAF and further enhanced the molecular
interaction of PCAF with p65 at the protein level in human AGS
cells, suggesting a potential ability of PCAF to acetylate p65.
PCAF induction contributes to p65
acetylation in AGS cells stimulated by CagA
To further determine the role of PCAF expression in
p65 acetylation and phosphorylation, AGS cells were treated with
PCAF shRNA expression vector for 36 h followed by CagA stimulation
for 1.5 h. A Co-IP assay showed that PCAF knockdown decreased
CagA-induced p65 acetylation and phosphorylation in AGS cells
(Fig. 5A). Conversely,
overexpression of PCAF triggered p65 acetylation and
phosphorylation in AGS cells (Fig.
5B). These results suggested that PCAF is necessary for p65
acetylation and phosphorylation in CagA-induced AGS cells.
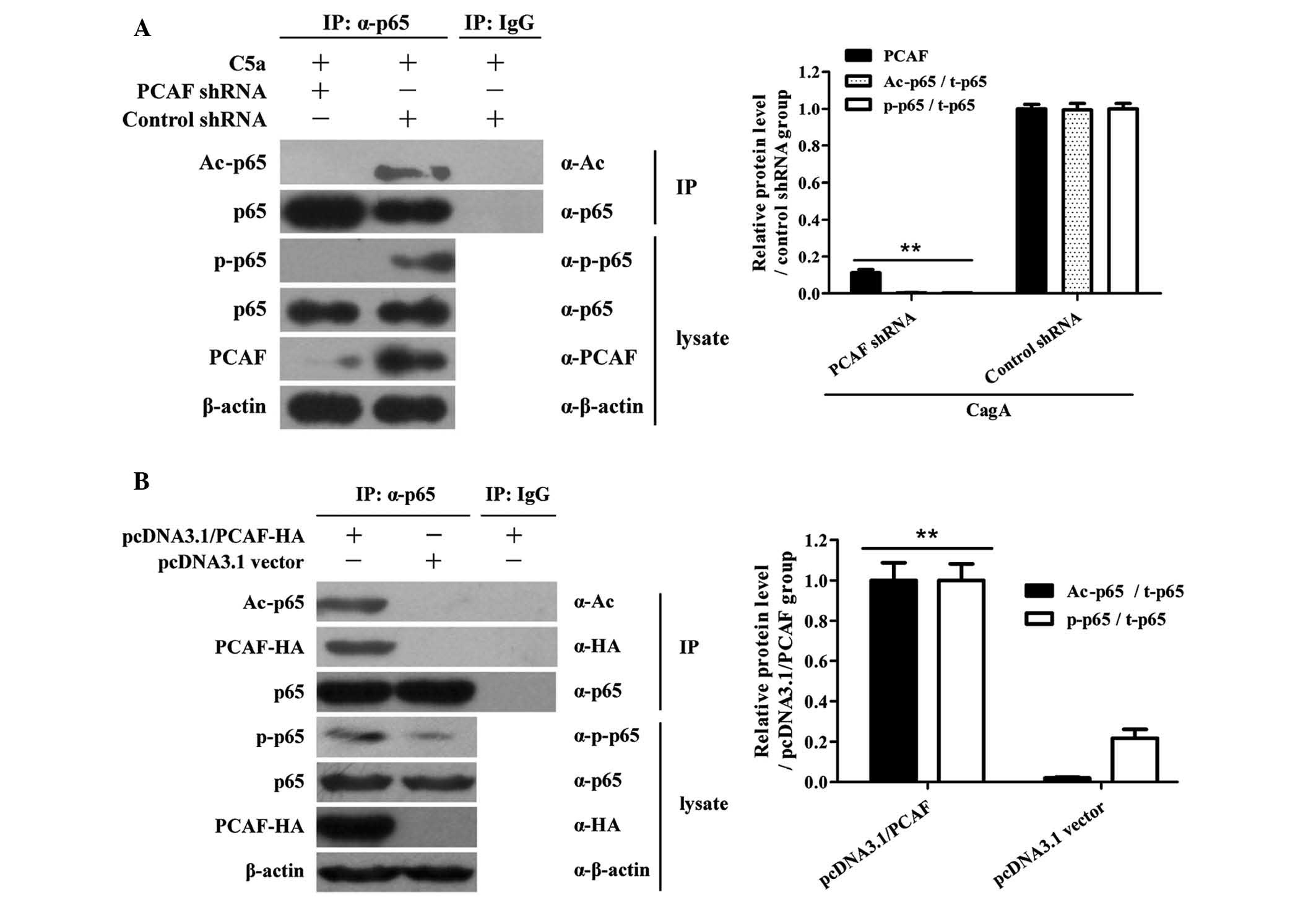 | Figure 5PCAF induction promotes p65
acetylation in AGS cells stimulated by CagA. (A) AGS cells were
transfected with PCAF shRNA for 36 h and then stimulated with 10
μg/ml CagA for 1.5 h. Cell lysates were then subjected to
immunoprecipitation with anti-p65 antibodies. Western blot analysis
was then used to detect the levels of Ac-p65 and total p65 in the
immunoprecipitated complex with anti-A-p65c and anti-p65
antibodies. In addition, western blot analysis was used to detect
p-p65, p65, PCAF and β-actin levels in cell lysates without
immunoprecipitation. The results showed that PCAF shRNA decreased
CagA-induced p65 acetylation and phosphorylation in AGS cells.
**P<0.01 compared to control shRNA + CagA group. (B)
AGS cells were treated with pcDNA3.1 PCAF-HA overexpression vector
for 48 h. Cell lysates were then subjected to immunoprecipitation
with anti-p65 antibodies. Subsequently, the levels of PCAF-HA,
Ac-p65 and total p65 in the immunoprecipitated complexes were
detected by western blot analysis with anti-HA, anti-Ac-p65 and
anti-p65 antibodies. In addition, the levels of p-p65, p65, PCAF-HA
and β-actin were detected by western blot in cell lysates without
immunoprecipitation. The results showed that overexpression of PCAF
triggered p65 acetylation and phosphorylation in AGS cells.
**P<0.01 compared to pcDNA3.1 vector group. Results
shown are from one experiment, representative of three independent
experiments. Values are expressed as the mean ± standard deviation
(n=3 for each group). CagA, cytotoxin-associated gene A; PCAF,
P300/CBP-associated factor; PBS, phosphate-buffered saline; IP,
immunoprecipitation; IgG, immunoglobulin G; shRNA, small hairpin
RNA; Ac, acetylated; p, phosphorylated; HA, hemagglutinin tag. |
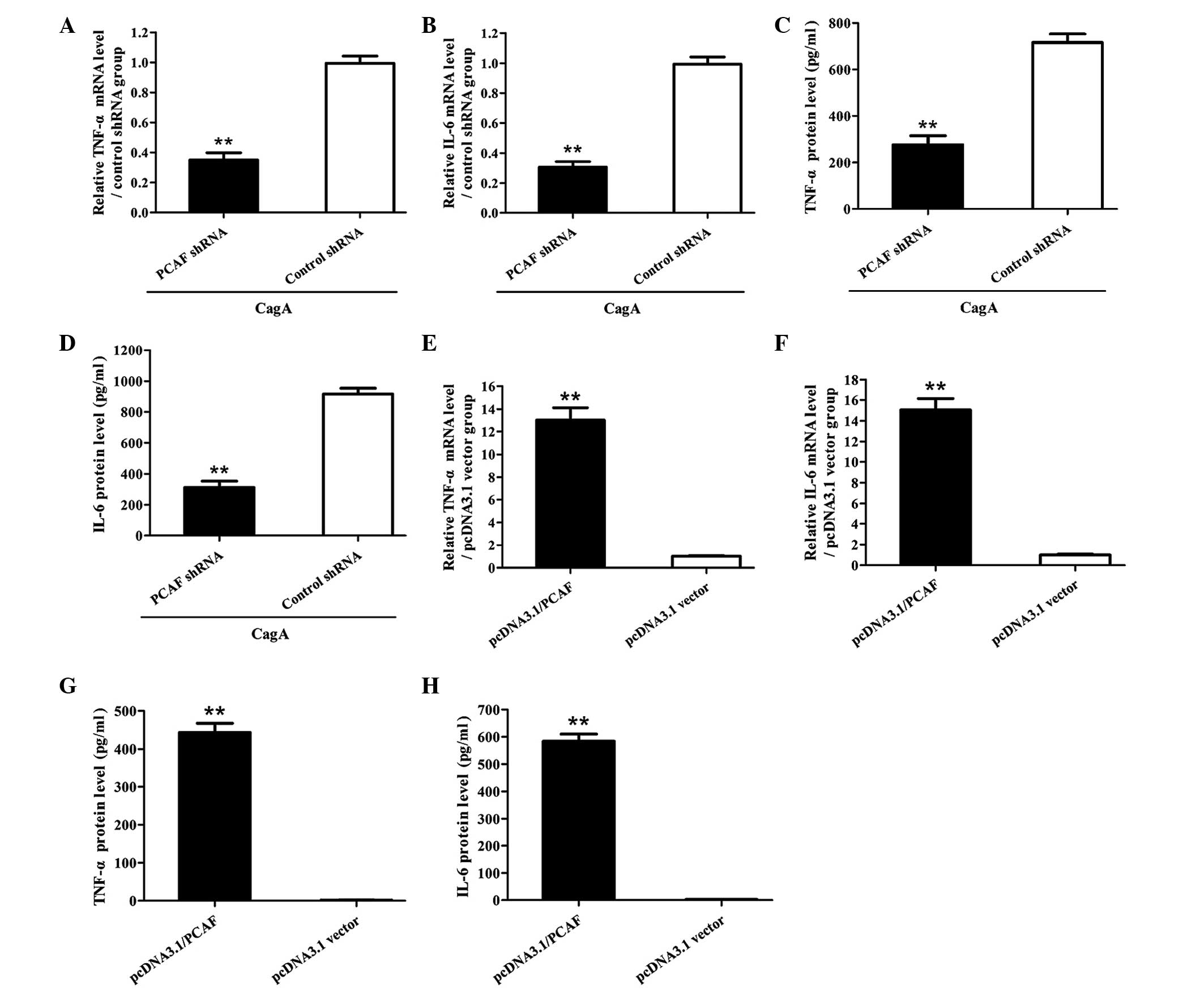
PCAF-mediated p65 acetylation is required
for CagA-induced production of TNF-α and IL-6 in AGS cells
Since PCAF was shown to be required for p65
acetylation in AGS cells stimulated by CagA (Fig. 5), the present study further
explored the role of PCAF-mediated p65 acetylation in the
production of TNF-α and IL-6 in AGS cells induced by CagA. The
results showed that suppression of PCAF markedly reduced the
production of TNF-α and IL-6 in AGS cells stimulated by CagA
(Fig. 6A-D). Conversely, AGS cells
overexpressing PCAF exhibited enhanced production of TNF-α and IL-6
(Fig. 6E-H). These results
indicated that PCAF-mediated p65 acetylation is involved in the
production of TNF-α and IL-6 in AGS cells stimulated with CagA.
Discussion
Helicobacter pylori is a Gram-negative
microaerophilic bacterium that selectively colonizes in human
gastric and duodenal mucosa. It is generally known that persistent
Helicobacter pylori infection leads to chronic gastritis and
severe gastric pathologies, including peptic ulcers and even
gastric cancer (39–41). It has been reported that infection
induces not only innate immune responses but also adaptive immune
responses against microorganisms; however in most cases, this fails
to eradicate the microorganisms. By contrast, permanent infection
ultimately triggers chronic inflammation and further leads to the
damage of the body (42,43).
It is well accepted that Helicobacter pylori
infection triggers production of various cytokines, including
IL-1β, IL-6, IL-8, IL-18, IL-32, TNF-α and IL-6, which are known to
have important roles in chronic inflammation and ultimate disease
outcome (10,14,44–46).
Among these inflammatory cytokines, TNF-α and IL-6 are considered
to be two important inflammatory factors that can also be produced
in other chronic inflammatory diseases, including Crohn's disease,
rheumatoid arthritis and atherosclerosis (47–49).
These studies indicated that overproduction of TNF-α and IL-6 may
have a key role in mediating the inflammatory response in the
process of Helicobacter pylori infection. In addition, CagA
is considered to be an important virulence factor for
Helicobacter pylori (46,50).
Therefore, the present study aimed to investigate the implication
of Helicobacter pylori CagA in the inflammatory response of
human AGS cells in vitro. The results revealed that CagA
markedly stimulated the production of TNF-α and IL-6 in human AGS
cells in vitro, which is in accordance with the findings of
previous studies (50,51).
NF-κB is a major transcriptional factor that
participates in the regulation of numerous cellular functions
including inflammation (52–54).
It has been demonstrated that Helicobacter pylori infection
activates NF-κB and its target genes in gastric epithelial cells,
which is thought to be critical for Helicobacter
pylori-initiated chronic inflammation (9,21).
The virulence factor CagA, encoded by the Helicobacter
pylori Cag pathogenicity island, has an important role in the
pathogenicity of Helicobacter pylori, including
Helicobacter pylori-induced activation of NF-κB and
expression of NF-κB target genes (22,23).
However, the exact function of CagA in the activation of NF-κB and
the NF-κB-dependent inflammatory response have not been well
characterized. Emerging evidence indicated that post-translational
modification, including ubiquitination and acetylation, has
important roles in various biological events, including
inflammation (55–60). Among these, the acetylation of the
NF-κB p65 sub-unit has been shown to have an important role in its
activation (27–29). The present study showed that the
acetylation level of the NF-κB p65 sub-unit was significantly
enhanced in human AGS cells incubated with CagA in vitro.
Therefore, the present study aimed to investigate the regulatory
mechanism of p65 acetylation in CagA-induced gastric adenocarcinoma
cells.
Recent studies showed that PCAF functions as an
acetyl transferase (30–33); furthermore, PCAF has been reported
to be linked to infection and inflammation (34,35,61).
However, the roles of PCAF in mediating p65 activation and the
production of TNF-α and IL-6 in CagA-induced AGS cells has remained
largely elusive. The present study therefore investigated the
expression of PCAF in human AGS cells induced by CagA, and showed
that CagA increased the mRNA and protein expression levels of PCAF
in human AGS cells. Furthermore, the results of the present study
demonstrated that PCAF was required for p65 acetylation and
phosphorylation in human AGS cells stimulated with CagA, suggesting
that p65 acetylation contributed to p65 phosphorylation. Of note,
PCAF was demonstrated to have an important role in p65 acetylation,
although PCAF expression was not strongly increased. This result
indicated that, in addition to the expression of PCAF, the
activation of PCAF may have had a predominant role in the
regulation of p65 acetylation in human AGS cells stimulated with
CagA.
In the present study, it was also demonstrated that
PCAF-mediated p65 acetylation functionally contributed to the
production of TNF-α and IL-6 in human AGS cells stimulated by CagA,
indicating that PCAF may have an important role in promoting the
production of inflammatory factors. Kiernan et al (62) revealed that acetylation of p65 may
have a key role in inhibitor of NF-κB-mediated attenuation of NF-κB
transcription. The present study showed that p65 acetylation
promoted its activation as a transcription factor to activate TNF-α
and IL-6 gene expression. This difference may be due to different
acetylated sites of p65. Therefore, further studies are required to
identify the differential sites of PCAF-induced acetylation in AGS
cells stimulated by CagA.
In conclusion, the results of the present study
suggested that Helicobacter pylori CagA promoted the
production of TNF-α and IL-6 in AGS cells through PCAF-mediated p65
acetylation. The present study therefore provided novel insight
into the pathomechanism of Helicobacter pylori
infection.
References
|
1
|
Ueda J, Gosho M, Inui Y, Matsuda T,
Sakakibara M, Mabe K, Nakajima S, Shimoyama T, Yasuda M, Kawai T,
et al: Prevalence of Helicobacter pylori infection by birth year
and geographic area in Japan. Helicobacter. 19:105–110. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Shiota S, Murakami K, Okimoto T, Kodama M
and Yamaoka Y: Serum Helicobacter pylori CagA antibody titer as a
useful marker for advanced inflammation in the stomach in Japan. J
Gastroenterol Hepatol. 29:67–73. 2014. View Article : Google Scholar
|
|
3
|
Bhuiyan TR, Islam MM, Uddin T, Chowdhury
MI, Janzon A, Adamsson J, Lundin SB, Qadri F and Lundgren A: Th1
and Th17 responses to Helicobacter pylori in Bangladeshi infants,
children and adults. PLoS One. 9:e939432014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Guo S, He L, Zhang J, Zhou L, Ding Z,
Zhang J, Yu F, Wang G, Zhou J, Guan D, et al: Antibiotic resistance
of Helicobacter pylori in children and macrolide-resistant
genotypes in Helicobacter pylori. Zhonghua Yi Xue Za Zhi.
94:563–566. 2014.In Chinese. PubMed/NCBI
|
|
5
|
Yu Y, Su L, Wang X, Wang X and Xu C:
Association between Helicobacter pylori infection and pathological
changes in the gastric mucosa in Chinese children. Intern Med.
53:83–88. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Basiri Z, Safaralizadeh R, Bonyadi MJ,
Somi MH, Mahdavi M and Latifi-Navid S: Helicobacter pylori vacA d1
Genotype predicts risk of gastric adenocarcinoma and peptic ulcers
in Northwestern Iran. Asian Pac J Cancer Prev. 15:1575–1579. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhang L, Sung JJ, Yu J, Ng SC, Wong SH,
Cho CH, Ng SS, Chan FK and Wu WK: Xenophagy in Helicobacter pylori-
and Epstein-Barr virus-induced gastric cancer. J Pathol.
233:103–112. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sokolova O, Maubach G and Naumann M: MEKK3
and TAK1 synergize to activate IKK complex in Helicobacter pylori
infection. Biochim Biophys Acta. 1843:715–724. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Devi S, Ansari SA, Vadivelu J, Mégraud F,
Tenguria S and Ahmed N: Helicobacter pylori antigen HP0986 (TieA)
interacts with cultured gastric epithelial cells and induces IL8
secretion via NF-κB mediated pathway. Helicobacter. 19:26–36. 2014.
View Article : Google Scholar
|
|
10
|
Peng LS, Zhuang Y, Li WH, Zhou YY, Wang
TT, Chen N, Cheng P, Li BS, Guo H, Yang SM, et al: Elevated
interleukin-32 expression is associated with Helicobacter
pylori-related gastritis. PLoS One. 9:e882702014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhao Y, Wang JW, Tanaka T, Hosono A, Ando
R, Tokudome S, Soeripto, Triningsih FX, Triono T, Sumoharjo S, et
al: Association between TNF-α and IL-1β genotypes vs Helicobacter
pylori infection in Indonesia. World J Gastroenterol. 19:8758–8763.
2013. View Article : Google Scholar :
|
|
12
|
Allison CC, Ferrand J, McLeod L, Hassan M,
Kaparakis-Liaskos M, Grubman A, Bhathal PS, Dev A, Sievert W,
Jenkins BJ and Ferrero RL: Nucleotide oligomerization domain 1
enhances IFN-γ signaling in gastric epithelial cells during
Helicobacter pylori infection and exacerbates disease severity. J
Immunol. 190:3706–3715. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Abdollahi H, Shams S, Zahedi MJ, Darvish
Moghadam S, Hayatbakhsh MM and Jafarzadeh A: IL-10, TNF-α and IFN-γ
levels in serum and stomach mucosa of Helicobacter pylori-infected
patients. Iran J Allergy Asthma Immunol. 10:267–271.
2011.PubMed/NCBI
|
|
14
|
Nakagawa H, Tamura T, Mitsuda Y, Goto Y,
Kamiya Y, Kondo T, Wakai K and Hamajima N: Significant association
between serum interleukin-6 and Helicobacter pylori antibody levels
among H. pylori-positive Japanese adults. Mediators Inflamm.
2013:1423582013. View Article : Google Scholar
|
|
15
|
Zhang X, Yang Y, Zhu R, Bai J, Tian Y, Li
X, Peng Z, He Y, Chen L, Fang D, et al: H. pylori induces the
expression of Hath1 in gastric epithelial cells via
interleukin-8/STAT3 phosphorylation while suppressing Hes1. J Cell
Biochem. 113:3740–3751. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhang MS, Zhang KJ, Zhang J, Jiao XL, Chen
D and Zhang DL: Phospholipases A-II (PLA2-II) induces acute
pancreatitis through activation of the transcription factor
NF-kappaB. Eur Rev Med Pharmacol Sci. 18:1163–1169. 2014.PubMed/NCBI
|
|
17
|
Cuadrado A, Martin-Moldes Z, Ye J and
Lastres-Becker I: Transcription factors NRF2 and NF-κB are
coordinated effectors of the Rho family, GTP-binding protein RAC1
during inflammation. J Biol Chem. 289:15244–15258. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Saravanan S, Islam VI, Babu NP, Pandikumar
P, Thirugnanasambantham K, Chellappandian M, Raj CS, Paulraj MG and
Ignacimuthu S: Swertiamarin attenuates inflammation mediators via
modulating NF-κB/IκB and JAK2/STAT3 transcription factors in
adjuvant induced arthritis. Eur J Pharm Sci. 56:70–86. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kim GD, Oh J, Park HJ, Bae K and Lee SK:
Magnolol inhibits angiogenesis by regulating ROS-mediated apoptosis
and the PI3K/AKT/mTOR signaling pathway in mES/EB-derived
endothelial-like cells. Int J Oncol. 43:600–610. 2013.PubMed/NCBI
|
|
20
|
Tan GK and Tabata Y: Chondroitin-6-sulfate
attenuates inflammatory responses in murine macrophages via
suppression of NF-κB nuclear translocation. Acta Biomater.
10:2684–2692. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chen JP, Wu MS, Kuo SH and Liao F: IL-22
negatively regulates Helicobacter pylori-induced CCL20 expression
in gastric epithelial cells. PLoS One. 9:e973502014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kang DW, Hwang WC, Park MH, Ko GH, Ha WS,
Kim KS, Lee YC, Choi KY and Min DS: Rebamipide abolishes
Helicobacter pylori CagA-induced phospholipase D1 expression via
inhibition of NFκB and suppresses invasion of gastric cancer cells.
Oncogene. 32:3531–3542. 2013. View Article : Google Scholar
|
|
23
|
Lamb A, Yang XD, Tsang YH, Li JD, Higashi
H, Hatakeyama M, Peek RM, Blanke SR and Chen LF: Helicobacter
pylori CagA activates NF-kappaB by targeting TAK1 for
TRAF6-mediated Lys 63 ubiquitination. EMBO Rep. 10:1242–1249. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chen H, Li J, Jiao L, Petersen RB, Li J,
Peng A, Zheng L and Huang K: Apelin inhibits the development of
diabetic nephropathy by regulating histone acetylation in Akita
mouse. J Physiol. 592:505–521. 2014. View Article : Google Scholar :
|
|
25
|
Yang H, Lee SM, Gao B, Zhang J and Fang D:
Histone deacetylase sirtuin 1 deacetylates IRF1 protein and
programs dendritic cells to control Th17 protein differentiation
during autoimmune inflammation. J Biol Chem. 288:37256–37266. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Qiu W, Zhou J, Zhu G, Zhao D, He F, Zhang
J, Lu Y, Yu T, Liu L and Wang Y: Sublytic C5b-9 triggers glomerular
mesangial cell apoptosis via XAF1 gene activation mediated by
p300-dependent IRF-1 acetylation. Cell Death Dis. 5:e11762014.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hah YS, Cheon YH, Lim HS, Cho HY, Park BH,
Ka SO, Lee YR, Jeong DW, Kim HO, Han MK and Lee SI: Myeloid
deletion of SIRT1 aggravates serum transfer arthritis in mice via
nuclear factor-κB activation. PLoS One. 9:e877332014. View Article : Google Scholar
|
|
28
|
Hwang YJ, Lee EW, Song J, Kim HR, Jun YC
and Hwang KA: MafK positively regulates NF-κB activity by enhancing
CBP-mediated p65 acetylation. Sci Rep. 3:32422013. View Article : Google Scholar
|
|
29
|
Cuccurazzu B, Bortolotto V, Valente MM,
Ubezio F, Koverech A, Canonico PL and Grilli M: Upregulation of
mGlu2 receptors via NF-κB p65 acetylation is involved in the
Proneurogenic and antidepressant effects of acetyl-L-carnitine.
Neuropsychopharmacology. 38:2220–2230. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Puttagunta R, Tedeschi A, Sória MG,
Hervera A, Lindner R, Rathore KI, Gaub P, Joshi Y, Nguyen T,
Sehmandke A, et al: PCAF-dependent epigenetic changes promote
axonal regeneration in the central nervous system. Nat Commun.
5:35272014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hu P, Wang X, Zhang B, Zhang S, Wang Q and
Wang Z: Fluorescence polarization for the evaluation of
small-molecule inhibitors of PCAF BRD/Tat-AcK50 association. Chem
Med Chem. 9:928–931. 2014. View Article : Google Scholar
|
|
32
|
Zhu LH, Sun LH, Hu YL, Jiang Y, Liu HY,
Shen XY, Jin XY, Zhen X, Sun HX and Yan GJ: PCAF impairs
endometrial receptivity and embryo implantation by down-regulating
β3-integrin expression via HOXA10 acetylation. J Clin Endocrinol
Metab. 98:4417–4428. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zheng X, Gai X, Ding F, Lu Z, Tu K, Yao Y
and Liu Q: Histone acetyltransferase PCAF up-regulated cell
apoptosis in hepatocellular carcinoma via acetylating histone H4
and inactivating AKT signaling. Mol Cancer. 12:962013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhao J, Gong AY, Zhou R, Liu J, Eischeid
AN and Chen XM: Downregulation of PCAF by miR-181a/b provides
feedback regulation to TNF-α-induced transcription of
proinflammatory genes in liver epithelial cells. J Immunol.
188:1266–1274. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Bastiaansen AJ, Ewing MM, de Boer HC, van
der Pouw Kraan TC, de Vries MR, Peters EA, Welten SM, Arens R,
Moore SM, Faber JE, et al: Lysine acetyltransferase PCAF is a key
regulator of arteriogenesis. Arterioscler Thromb Vasc Biol.
33:1902–1910. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ong SP, Lee LM, Leong YF, Ng ML and Chu
JJ: Dengue virus infection mediates HMGB1 release from monocytes
involving PCAF acetylase complex and induces vascular leakage in
endothelial cells. PLoS One. 7:e419322012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Li W, Cun W, Liu L, Hong M, Wang L, Wang
L, Dong C and Li Q: The transactivating effect of HSV-1 ICP0 is
enhanced by its interaction with the PCAF component of histone
acetyltransferase. Arch Virol. 154:1755–1764. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Hou F, Wang L, Wang H, Gu J, Li M, Zhang
J, Ling X, Gao X and Luo C: Elevated gene expression of S100A12 is
correlated with the predominant clinical inflammatory factors in
patients with bacterial pneumonia. Mol Med Rep. 11:4345–4352.
2015.PubMed/NCBI
|
|
39
|
Talaiezadeh A, Hajiani E and Tarshizi MA:
The relative frequency of the Helicobacter pylori Infection in
proximal gastric cancers. Pol Przegl Chir. 85:657–662. 2013.
|
|
40
|
Zhu Y, Zhou X, Wu J, Su J and Zhang G:
Risk factors and prevalence of Helicobacter pylori infection in
persistent high incidence area of gastric carcinoma in Yangzhong
City. Gastroenterol Res Pract. 2014:4813652014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Repetto O, Zanussi S, Casarotto M,
Canzonieri V, De Paoli P, Cannizzaro R and De Re V: Differential
proteomics of Helicobacter pylori associated with autoimmune
atrophic gastritis. Mol Med. 20:57–71. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Satkunanathan S, Kumar N, Bajorek M,
Purbhoo MA and Culley FJ: Respiratory syncytial virus infection,
TLR3 ligands and proinflammatory cytokines induce CD161 ligand LLT1
expression on the respiratory epithelium. J Virol. 88:2366–2373.
2014. View Article : Google Scholar :
|
|
43
|
Mota A, Areias J and Cardoso MF: Chronic
liver disease and cirrhosis among patients with hepatitis B virus
infection in northern Portugal with reference to the viral
genotypes. J Med Virol. 83:71–77. 2011. View Article : Google Scholar
|
|
44
|
Srivastava R, Kashyap A, Kumar M, Nath G
and Jain AK: Mucosal IgA & IL-1β in Helicobacter pylori
infection. Indian J Clin Biochem. 28:19–23. 2013. View Article : Google Scholar :
|
|
45
|
Kim DJ, Park JH, Franchi L, Backert S and
Núñez G: The Cag pathogenicity island and interaction between
TLR2/NOD2 and NLRP3 regulate IL-1β production in Helicobacter
pylori infected dendritic cells. Eur J Immunol. 43:2650–2658. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Bartchewsky W Jr, Martini MR, Masiero M,
Squassoni AC, Alvarez MC, Ladeira MS, Salvatore D, Trevisan M,
Pedrazzoli J Jr and Ribeiro ML: Effect of Helicobacter pylori
infection on IL-8, IL-1beta and COX-2 expression in patients with
chronic gastritis and gastric cancer. Scand J Gastroenterol.
44:153–161. 2009. View Article : Google Scholar
|
|
47
|
Augustine MV, Leonard MB, Thayu M,
Baldassano RN, de Boer IH, Shults J, Denson LA, DeBoer MD,
Herskovitz R and Denburg MR: Changes in vitamin D-related mineral
metabolism after induction with anti-tumor necrosis factor-α
therapy in crohn's disease. J Clin Endocrinol Metab. 99:E991–E998.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Lv Q, Yin Y, Li X, Shan G2, Wu X, Liang D,
Li Y and Zhang X: The status of rheumatoid factor and anti-cyclic
citrullinated peptide antibody are not associated with the effect
of anti-TNF α agent treatment in patients with rheumatoid
arthritis: A meta-analysis. PLoS One. 9:e894422014. View Article : Google Scholar
|
|
49
|
Schuett H, Oestreich R, Waetzig GH, Annema
W, Luchtefeld M, Hillmer A, Bavendiek U, von Felden J, Divchev D,
Kempf T, et al: Transsignaling of interleukin-6 crucially
contributes to atherosclerosis in mice. Arterioscler Thromb Vasc
Biol. 32:281–290. 2012. View Article : Google Scholar
|
|
50
|
Abbas Z, Yakoob J, Usman MW, Shakir T,
Hamid S and Jafri W: Effect of Helicobacter pylori and its
virulence factors on portal hypertensive gastropathy and
interleukin (IL)-8, IL-10 and tumor necrosis factor-alpha levels.
Saudi J Gastroenterol. 20:120–127. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Figura N, Palazzuoli A, Vaira D, Campagna
M, Moretti E, Iacoponi F, Giordano N, Clemente S, Nuti R and
Ponzetto A: Cross-sectional study: CagA-positive Helicobacter
pylori infection, acute coronary artery disease and systemic levels
of B-type natriuretic peptide. J Clin Pathol. 67:251–257. 2014.
View Article : Google Scholar
|
|
52
|
Peng Q, Liu H, Shi S and Li M: Lycium
ruthenicum polysaccharide attenuates inflammation through
inhibiting TLR4/NF-κB signaling pathway. Int J Biol Macromol.
67:330–335. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Luo JG, Zhao XL, Xu WC, Zhao XJ, Wang JN,
Lin XW, Sun T and Fu ZJ: Activation of spinal NF-κB/p65 contributes
to peripheral inflammation and hyperalgesia in rat adjuvant-induced
arthritis. Arthritis Rheumatol. 66:896–906. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Heymann MC, Winkler S, Luksch H, Flecks S,
Franke M, Ruß S, Ozen S, Yilmaz E, Klein C, Kallinich T, et al:
Human procaspase-1 variants with decreased enzymatic activity are
associated with febrile episodes and may contribute to inflammation
via RIP2 and NF-κB signaling. J Immunol. 192:4379–4385. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Corn JE and Vucic D: Ubiquitin in
inflammation: The right linkage makes all the difference. Nat
Struct Mol Biol. 21:297–300. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Liu Z, Qin T, Zhou J, Taylor A, Sparrow JR
and Shang F: Impairment of the ubiquitin-proteasome pathway in RPE
alters the expression of inflammation related genes. Adv Exp Med
Biol. 801:237–250. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Khan YM, Kirkham P, Barnes PJ and Adcock
IM: Brd4 Is essential for IL-1β-induced inflammation in human
airway epithelial cells. PLoS One. 9:e950512014. View Article : Google Scholar
|
|
58
|
Herbert C, Shadie AM, Bunting MM, Tedla N,
Garthwaite L, Freeman A, Yoo H, Park SH and Kumar RK:
Anti-inflammatory and anti-remodelling effects of ISU201, a
modified form of the extracellular domain of human BST2, in
experimental models of asthma: Association with inhibition of
histone acetylation. PLoS One. 9:e904362014. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Qiao G, Ying H, Zhao Y, Liang Y, Guo H,
Shen H, Li Z, Solway J, Tao E, Chiang YJ, et al: E3 ubiquitin
ligase Cbl-b suppresses proallergic T cell development and allergic
airway inflammation. Cell Rep. 6:709–723. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Soe NN, Sowden M, Baskaran P, Kim Y, Nigro
P, Smolock EM and Berk BC: Acetylation of cyclophilin A is required
for its secretion and vascular cell activation. Cardiovasc Res.
101:444–453. 2014. View Article : Google Scholar :
|
|
61
|
Chan C, Wang Y, Chow PK, Chung AY, Ooi LL
and Lee CG: Altered binding site selection of p53 transcription
cassettes by hepatitis B virus X protein. Mol Cell Biol.
33:485–497. 2013. View Article : Google Scholar :
|
|
62
|
Kiernan R, Brès V, Ng RW, Coudart MP, El
Messaoudi S, Sardet C, Jin DY, Emiliani S and Benkirane M:
Post-activation turn-off of NF-kappa B-dependent transcription is
regulated by acetylation of p65. J Biol Chem. 278:2758–2766. 2003.
View Article : Google Scholar
|