Introduction
Fulminant hepatitis has been reported to result from
a variety of causes, such as viral infection and drug poisoning,
and is commonly associated with a poor prognosis (1,2). In
the early stages of fulminant hepatitis, the excessive inflammatory
reactions serve a critical role in mediating the rapid loss of
hepatocytes (3,4). Therapies targeting the
pro-inflammatory mediators are suggested to provide protective
benefits in experimental studies (5,6). In
addition to the direct detrimental effects of pro-inflammatory
mediators, inflammation may also activate the coagulation system
(7,8). The inflammation-induced activation of
coagulation cascades markedly potentiates liver damage via
interrupting the blood supply and propagating further inflammation
(9,10).
It has been well documented that inflammation and
inflammation-induced coagulation are crucial pathological
mechanisms underling the progression of critical disorders,
including sepsis and multiple organ dysfunction syndrome (11,12).
The rapid induction of tissue factor (TF) under inflammatory
circumstances is the pivotal step associating inflammation with
coagulation (13). TF is the
essential initiator of the extrinsic pathway of blood coagulation
(14). The inflammatory stimuli
and inflammatory cytokines may markedly induce the expression of TF
and activate the extrinsic coagulation pathway (15). In addition to enhanced activation
of coagulation cascades, inflammation may induce the dysregulation
of fibrin removal via the upregulation of the plasminogen activator
inhibitor-1 (PAI-1) (16,17).
A previous study demonstrated that the antidiabetic
drug, metformin, attenuated lipopolysaccharide/D-galactosamine
(LPS/D- Gal)-induced fulm inant hepatitis (18). LPS/D-Gal-induced fulminant liver
damage predominantly depends on the induction of the
pro-inflammatory cytokine, tumor necrosis factor α (TNF-α), which
has been proposed as a key factor propagating the progression of
hepatitis induced by toxic insults (4,19).
In a previous study, metformin significantly suppressed the
expression of TNF-α, alleviating the liver lesions and improving
the survival rate of LPS/D-Gal-exposed mice (18).
Due to evidence indicating that the coagulation
system is involved in the pathogenesis of LPS/D-Gal-induced
fulminant hepatitis (20,21), the current study investigated
whether metformin is able to modulate the activation of the
coagulation response. In addition, the potential effects of
metformin on LPS/D-Gal-induced upregulation of TF and PAI-1 and the
subsequent metabolic disturbances were investigated.
Materials and methods
Materials
Metformin, LPS (from Escherichia coli,
055:B5) and D-Gal were purchased from Sigma-Aldrich (St. Louis, MO,
USA). The lactic acid (LA) assay kit was purchased from Nanjing
Jiancheng Bioengineering Institute (Nanjing, China). The Total
Protein Extraction kit was purchased from Beyotime Institute of
Biotechnology (Jiangsu, China). The mouse interleukin 6 (IL-6)
Enzyme-Linked Immunosorbent Assay (ELISA) kit was obtained from
NeoBioscience Technology Company (Shenzhen, China). The rabbit
anti-mouse monoclonal nuclear factor κB (NF-κB) p65 antibody
(D14E12; 1:1,000; cat. no. 8242), rabbit anti-mouse hypoxia
inducible factor 1α (HIF-1α) monoclonal antibody (D2U3T; 1:1,000;
cat. no. 14179), rabbit anti-mouse PAI-1 monoclonal antibody (D9C4;
1:1,000; cat. no. 11907), rabbit anti-mouse β-actin monoclonal
antibody (D6A8; 1:1,000; cat. no. 8457) and Histone 3: rabbit
anti-mouse Histone 3 monoclonal antibody (D1H2; 1:500; cat. no.
4499) were purchased from Cell Signaling Technology, Inc. (Danvers,
MA, USA). The rabbit anti-mouse TF monoclonal antibody (EPR8986;
1:1,000; cat. no. ab151748) was purchased from Abcam (Cambridge,
UK). The bicinchoninic acid (BCA) protein assay kit, horseradish
peroxidase-conjugated goat anti-rabbit polyclonal secondary
antibody (1:5,000; cat. no. G21234) and enhanced chemiluminescence
(ECL) reagents were obtained from Pierce Biotechnology, Inc.
(Rockford, IL, USA).
Animals
A total of 64 male Balb/c mice (16 in each group),
weighing 20–25 g, were obtained from the Experimental Animal Center
of Chongqing Medical University (Chongquing, China). All animals
were fed with a standard laboratory diet and water ad
libitum. They were housed in a specific pathogen-free room at a
temperature of 20–25°C and 50±5% relative humidity under a 12 h
dark/light cycle and had acclimatized for a minimum of l week prior
to use. All experimental procedures involving animals were approved
by the Animal Care and Use Committee of Chongqing Medical
University.
Induction of fulminant hepatitis
Balb/c mice were intraperitoneally injected with
vehicle or metformin (400 mg/kg, dissolved in normal saline),
administered 0.5 h prior to the injection of LPS (10 μg/kg)
combined with D-Gal (700 mg/kg). The dose of metformin was selected
based on previous experimental data (18). Subsequent to injection, the animals
were returned to their cages and allowed food and water ad
libitum. The experimental animals were allocated to four
groups: Control (CON), mice received vehicle administration only;
metformin (MET), mice treated with metformin without LPS/D-Gal
exposure; LPS/D-Gal, mice exposed to LPS/D-Gal; LPS/D-Gal + MET,
mice treated with metformin and exposed to LPS/D-Gal. Mice were
sacrificed by intraperitoneal injection of pentobarbital (50 mg/kg,
Sigma-Aldrich) at 1.5 h (n=8 per group) or 6 h (n=8 per group)
subsequent to LPS/D-Gal challenge, and blood samples and liver
tissues were harvested and stored at −80°C until required.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was isolated from liver samples using
TRIzol reagent (Takara Bio, Inc., Otsu, Japan). First-strand
complementary DNA was synthesized from 1 μg total RNA using
oligo-dT primer (Takara Bio, Inc.) and the M-MLV reverse
transcriptase (Takara Bio, Inc.). RT-qPCR was performed with SYBR
green PCR Master Mix (Takara Bio, Inc.). The sequences of the
primers used to amplify the target genes are presented in Table I. PCR was performed on a CFX96
Touch (Bio-Rad Laboratories, Inc., Hercules, CA, USA) using the
following PCR conditions: Denaturing at 95°C for 10 sec, annealing
at 58°C for 20 sec and elongation at 72°C for 20 sec. The mRNA
levels of TF, PAI-1, IL-6, erythropoietin (EPO), vascular
endothelial growth factor (VEGF) and matrix metalloproteinase-3
(MMP-3) were normalized to levels of β-actin.
 | Table ISequences of the primers for reverse
transcription-quantitative polymerase chain reaction. |
Table I
Sequences of the primers for reverse
transcription-quantitative polymerase chain reaction.
| Target gene | Forward primer | Reverse primer |
|---|
| TF |
5-CTTATCGGAAAGGCTCAA-3 |
5-CACCACTGCTCCCACAAT-3 |
| PAI-1 |
5-CATGTTTAGTGCAACCCTGGC-3 |
5-TGAGATGACAAAGGCTGTGGAG-3 |
| IL-6 |
5-AGTTGCCTTCTTGGGACTGATG-3 |
5-TCTCATTTCCACGATTTCCCAG-3 |
| EPO |
5-CAGCCACCAGAGACCCTTCA-3 |
5-TGTGAGTGTTCGGAGTGGAGC-3 |
| VEGF |
5-ACGATGAAGCCCTGGAGTGC-3 |
5-GCTCATCTCTCCTATGTGCTGGC-3 |
| MMP-3 |
5-CCACAGACTTGTCCCGTTTCC-3 |
5-GTGCTGACTGCATCAAAGAACAA-3 |
| β-actin |
5-CTGAGAGGGAAATCGTGCGT-3 |
5-CCACAGGATTCCATACCCAAGA-3 |
Western blot analysis
Total proteins from liver samples were prepared
using the Total Protein Extraction kit according to the
manufacturer's instructions. The total protein concentration was
determined using the BCA protein assay kit. Protein extracts were
fractionated on a 10% polyacrylamide-sodium dodecyl sulfate gel
(Beyotime Institute of Biotechnology) and then were transferred to
nitrocellulose membranes (Pierce Biotechnology, Inc.). The
membranes were blocked with 5% (w/v) nonfat milk in Tris-buffered
saline containing 0.05% Tween-20 (Enzo Life Sciences, Farmingdale,
NY, USA), prior to incubation with primary antibodies overnight at
4°C, followed by incubation with secondary antibodies for 1 h at
37°C. Antibody binding was visualized using an ECL system and a
short exposure of the membrane to X-ray films (Kodak, Rochester,
NY, USA).
ELISA
The protein levels of IL-6 in the plasma were
determined using the ELISA kits according to the manufacturer's
instructions (NeoBioscience Technology Company).
LA measurement
The LA contents in the liver tissue were determined
with an LA assay kit according to the manufacturer's instructions.
The LA values were assessed according to the absorbance measured at
530 nm (Varioscan Flash; Thermo Fisher Scientific, Waltham, MA,
USA). The levels of hepatic LA were normalized to the total protein
concentration in the same sample.
Statistical analysis
All data were presented as the mean ± standard
deviation. Statistical significance was determined by Student's
t-test for the comparison of two groups. Data were analyzed using
Statistical Package for Social Sciences (SPSS) software (version
16; SPSS, Inc., Chicago, IL, USA). P<0.05 was considered to
indicate a statistically significant difference.
Results
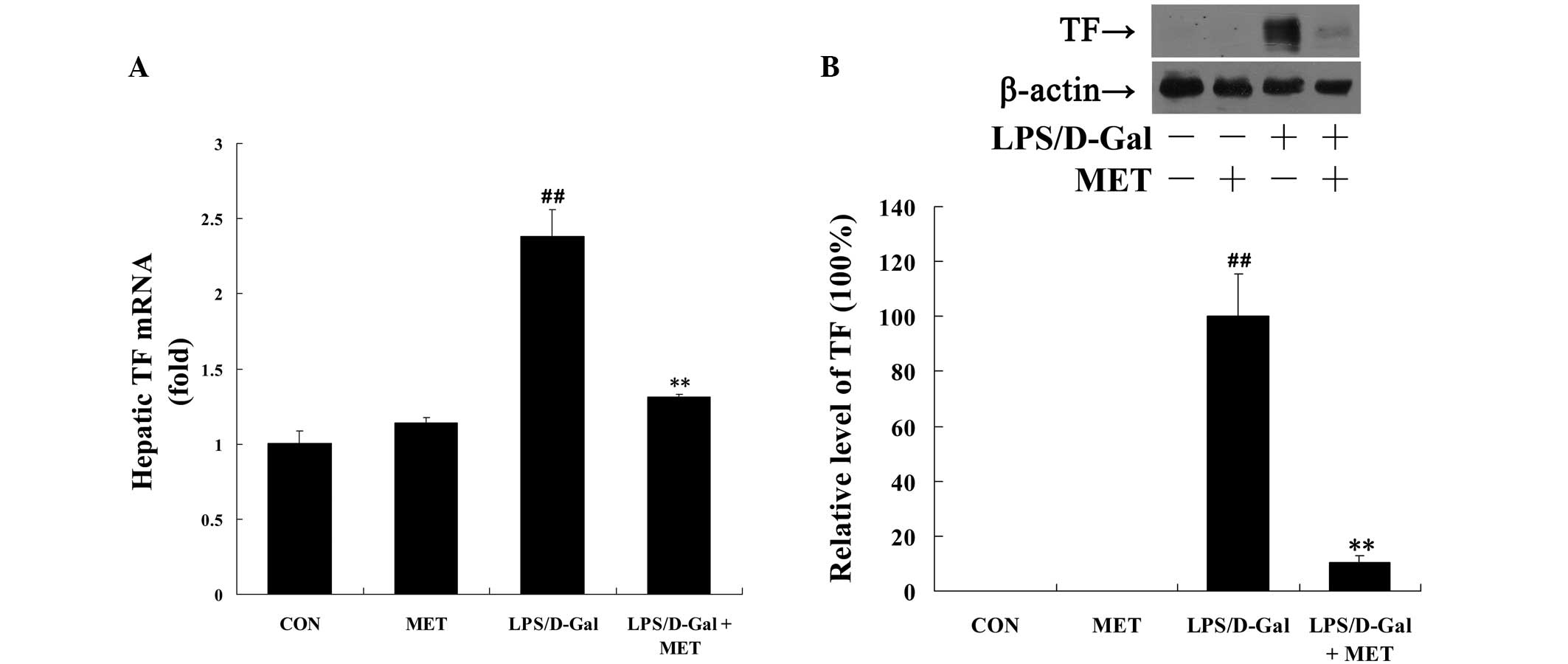
Metformin suppresses LPS/D-Gal-induced TF
and PAI-1 expression
TF is the initiator of the coagulation cascade,
which is rapidly induced by inflammatory stimuli (13). In the present study, LPS/D-Gal
exposure was demonstrated to markedly upregulate the mRNA and
protein levels of TF, compared with the control, which was
significantly reversed by metformin (P<0.01; Fig. 1). In addition, the
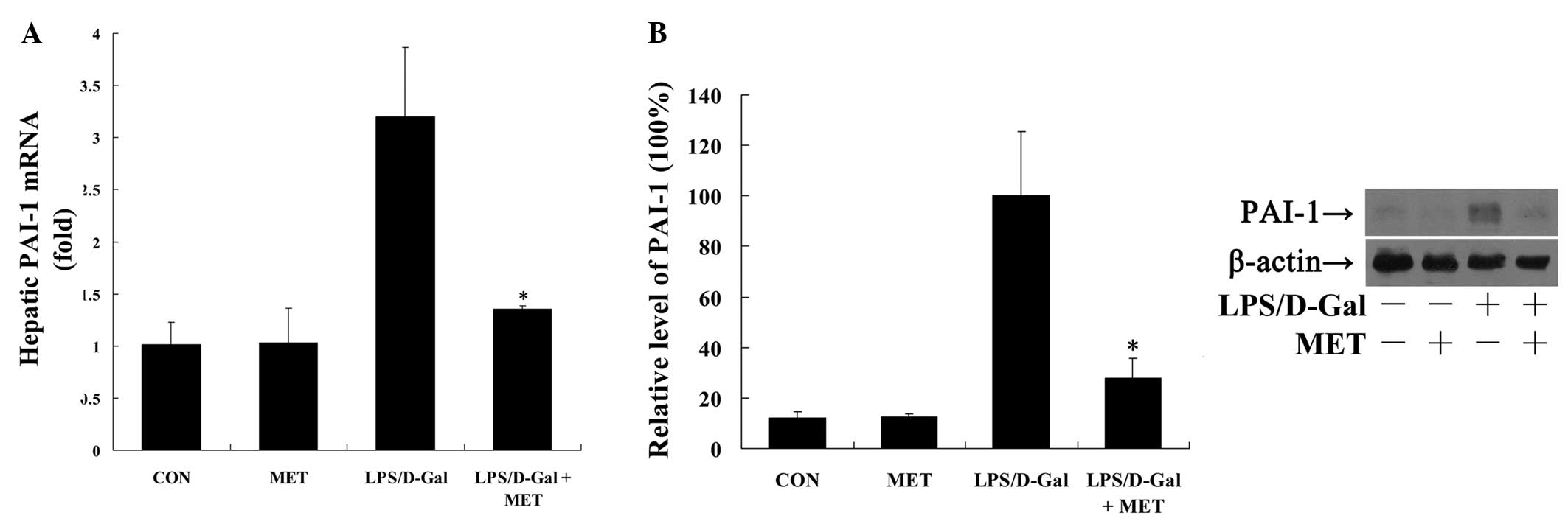
LPS/D-Gal-induced expression of PAI-1, an inflammation-induced
factor suppressing the fibrinolysis process (17), was also significantly inhibited by
metformin (P<0.05; Fig. 2).
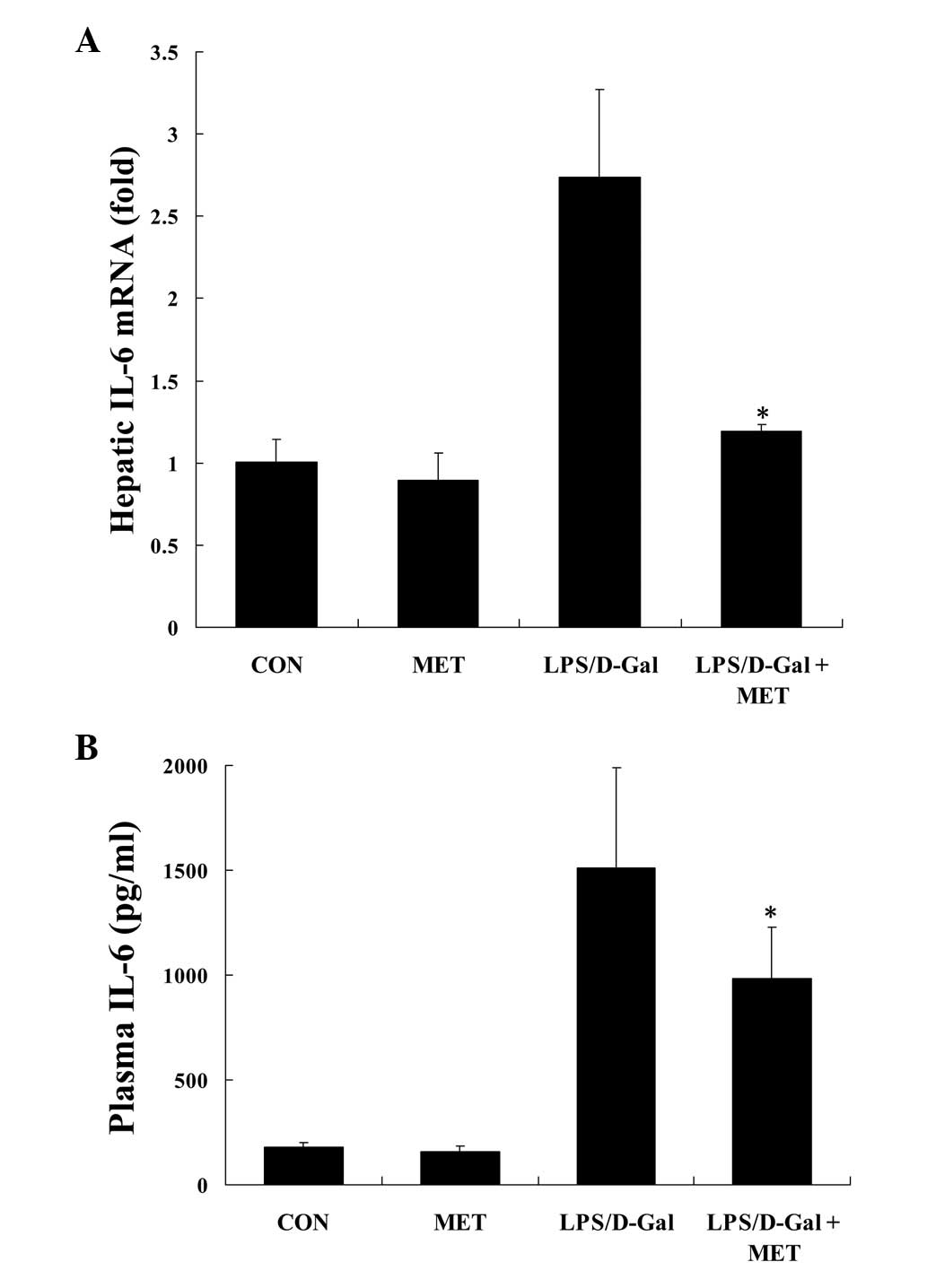
Metformin suppresses LPS/D-Gal-induced
IL-6 expression
IL-6 is a crucial pro-inflammatory cytokine linking
inflammation to coagulation (22).
In the current study, it was demonstrated that the challenge with
LPS/D-Gal induced the upregulation the mRNA levels of IL-6 in liver
tissue, whereas treatment with metformin significantly reduced the
level of IL-6 (P<0.05; Fig.
3A). In addition, similar inhibitory effects of metformin on
the plasma protein levels of IL-6 in LPS/D-Gal-exposed mice were
observed (P<0.05; Fig. 3B).
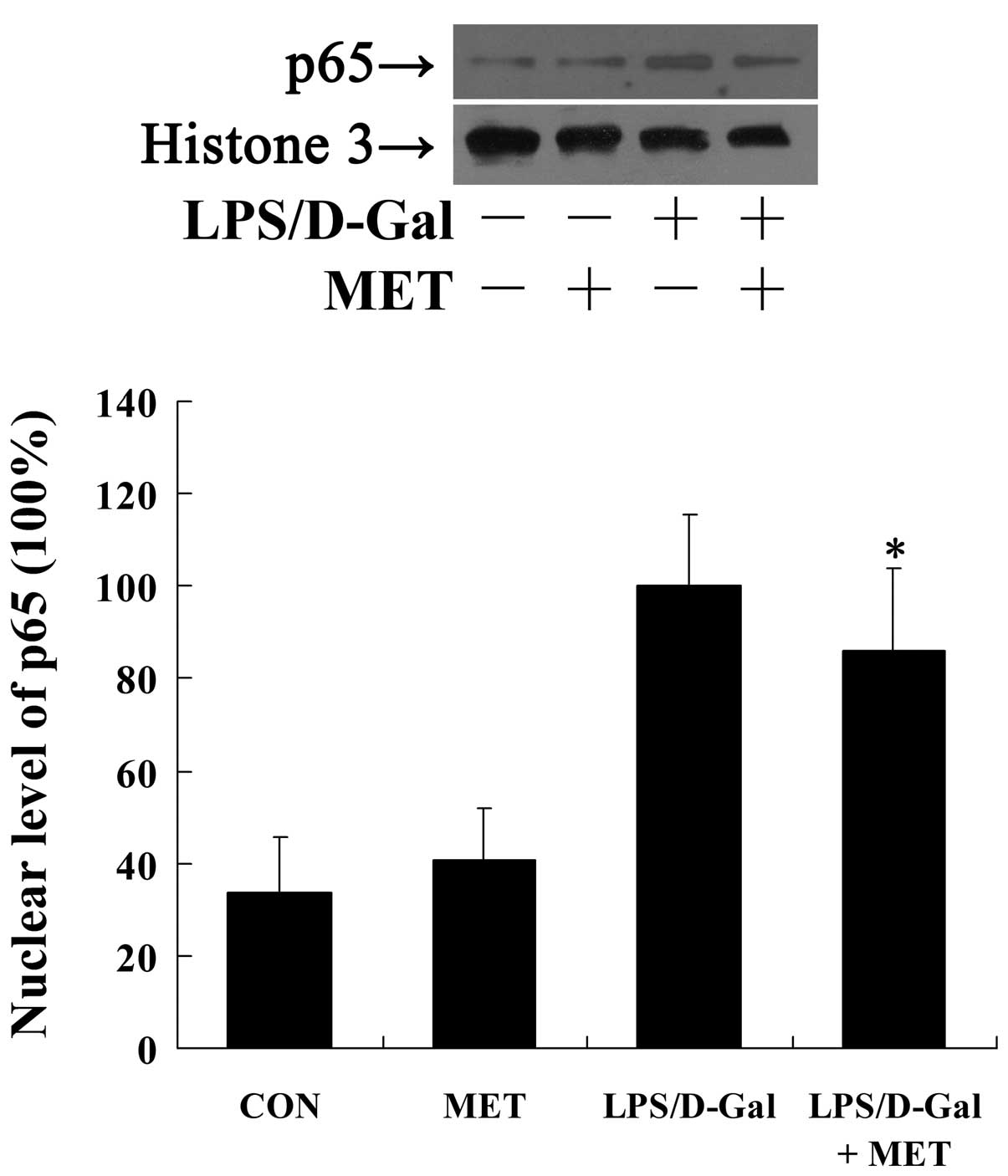
Metformin suppresses LPS/D-Gal-induced
NF-κB translocation
NF-κB is a pivotal transcriptional factor involved
in the transcriptional regulation of IL-6 and TF, and the nuclear
translocation of NF-κB is crucial for its activation (23,24).
In the present study, the exposure to LPS/D-Gal markedly induced
the translocation of NF-κB p65 into the nuclei indicated by the
increased nuclear level of p65 following LPS/D-Gal challenge,
whilst treatment with metformin significantly reduced the nuclear
level of p65 (P=0.023; Fig.
4).
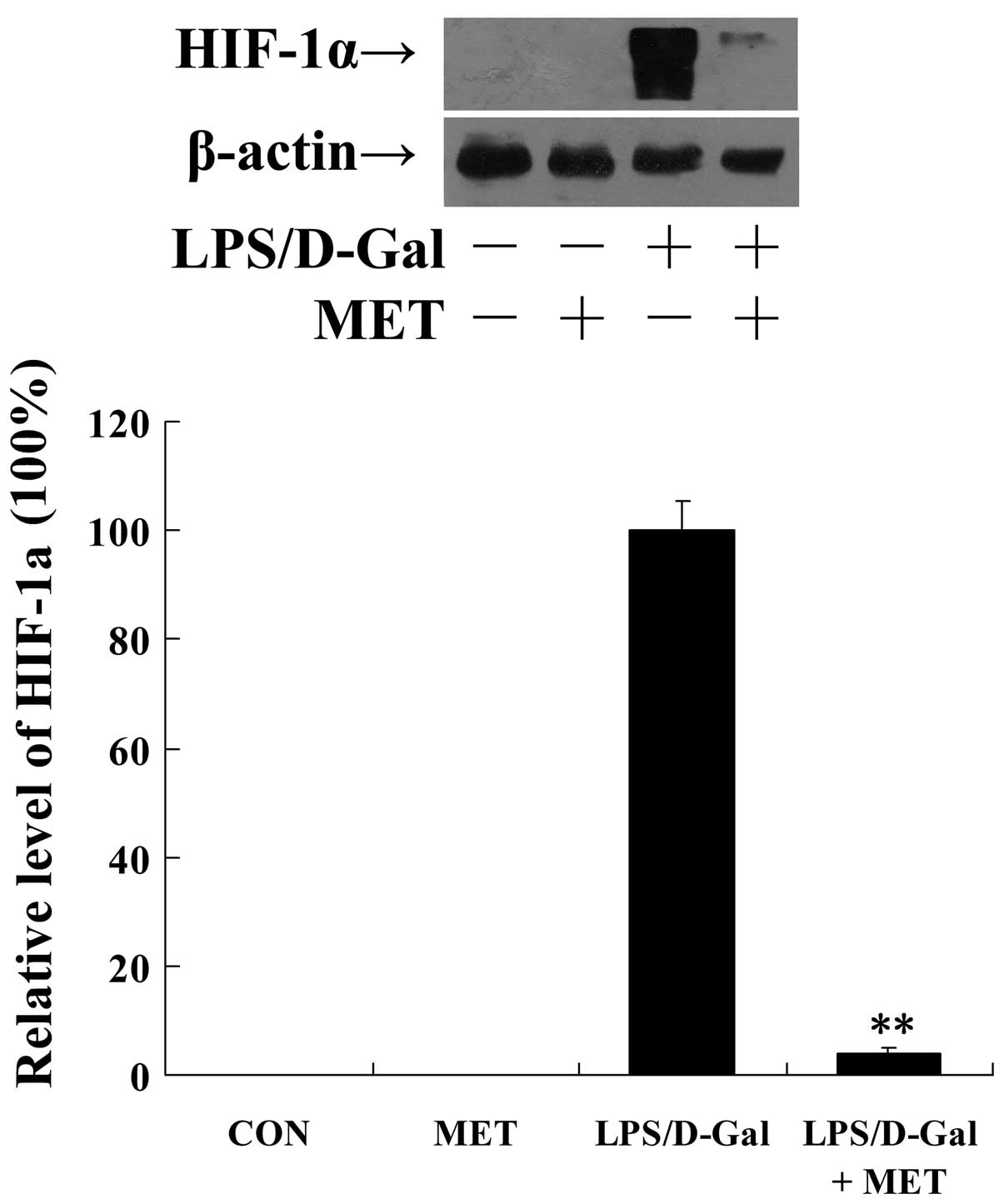
Metformin alleviates LPS/D-Gal-induced
hypoxia
Tissue hypoperfusion and hypoxia occur as a
consequence of coagulation activation (12). In the present study, the degree of
intrahepatic hypoxia was evaluated via the detection of the level
of HIF-1α. These data demonstrate that metformin significantly
suppressed the upregulation of HIF-1α protein levels (P<0.01;
Fig. 5). In agreement with this,
HIF-1-targeted genes, including EPO, VEGF and MMP-3 (25,26),
were upregulated following LPS/D-Gal exposure, however this
upregulation was suppressed by metformin (Fig. 6).
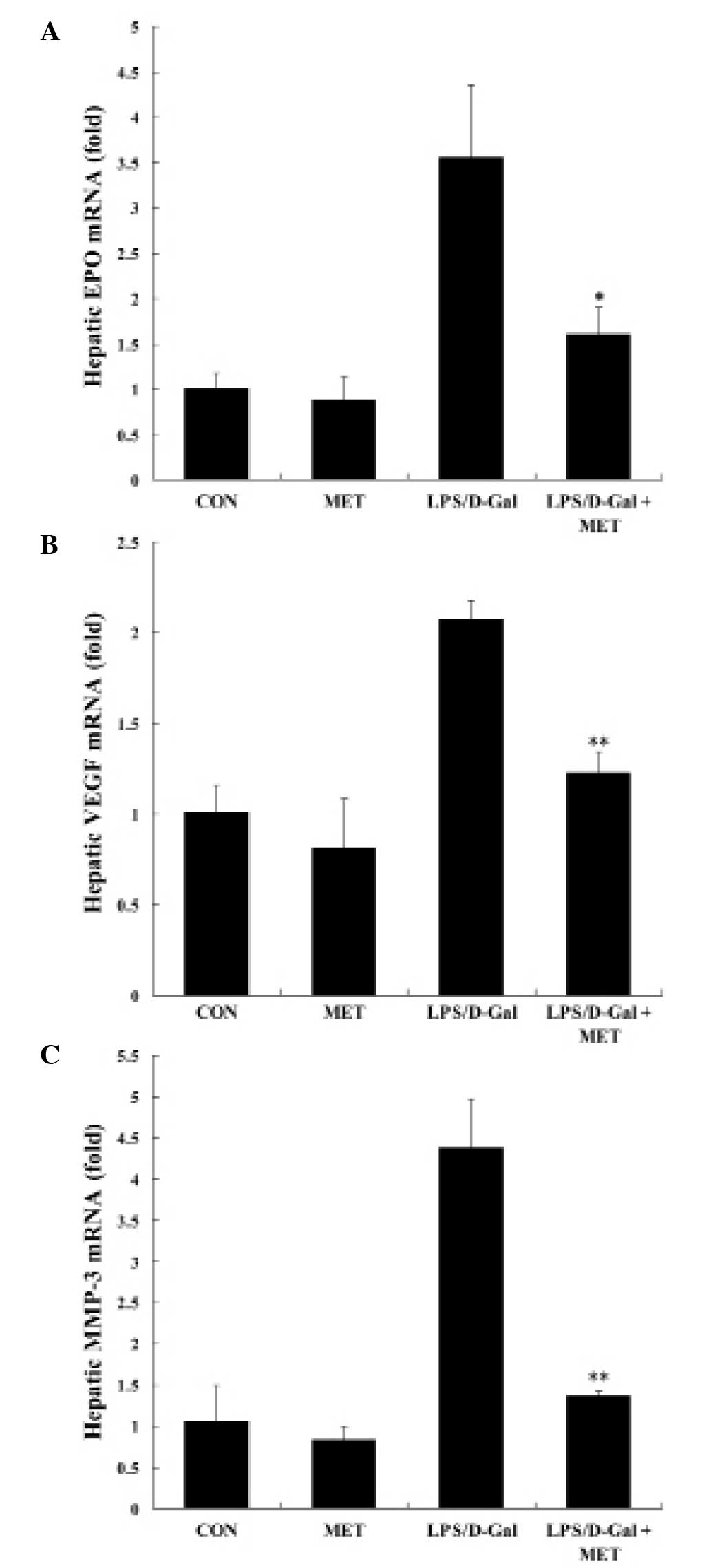 | Figure 6MET suppressed LPS/D-Gal-induced
upregulation of EPO, VEGF and MMP-3. Mice were treated with the
vehicle or MET (400 mg/kg) in the presence or absence of LPS/D-Gal
exposure. Liver samples were harvested 6 h subsequent to LPS/D-Gal
exposure and the mRNA levels of (A) EPO, (B) VEGF and (C) MMP-3
were determined. Data are presented as the mean ± standard
deviation, n=8. *P<0.05, **P<0.01 vs.
LPS/D-Gal group. MET, metformin; LPS, lipopolysaccharide; D-Gal,
D-galactosamine; EPO, erythropoietin; VEGF, vascular endothelial
growth factor; MMP-3, matrix metalloproteinase-3; CON, control. |
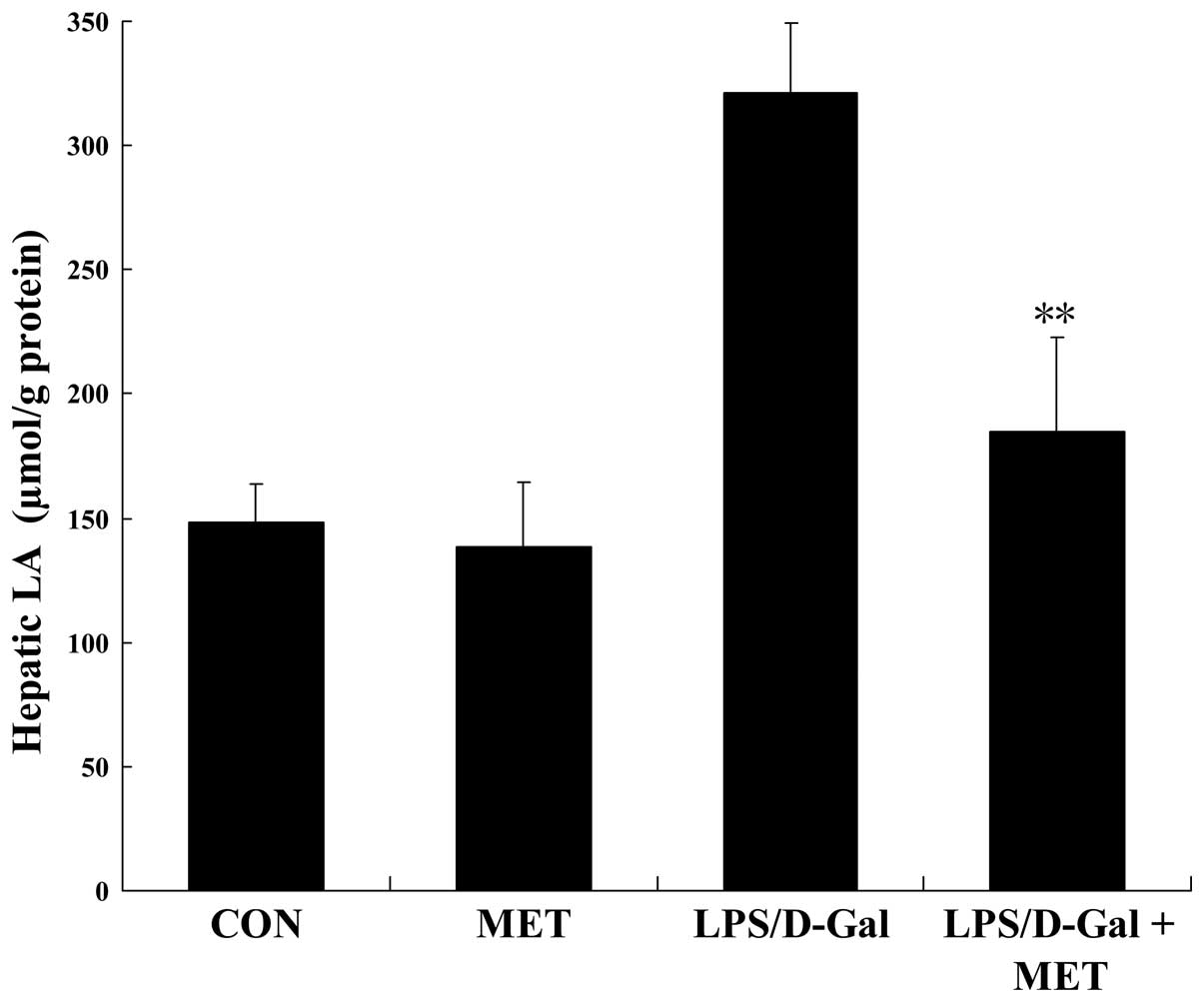
Metformin alleviates LPS/D-Gal-induced
metabolic disturbances
The metabolic disturbance resulting from hypoxia was
determined by detecting the level of LA, an anaerobic metabolic
product. The data collected indicate that LPS/D-Gal exposure
significantly increased the hepatic level of LA, which was
inhibited by metformin (Fig.
7).
Discussion
Metformin is used in the clinical treatment of
diabetes due to its hypoglycemic activity (27). In addition, studies have identified
the anti-inflammatory properties of metformin in vitro and
in vivo. In an LPS/D-Gal-induced hepatitis mouse model
(28–30), it was demonstrated that treatment
with the antidiabetic drug metformin provided therapeutic benefits.
This suggested that these protective effects may result from the
reduced production of the pro-inflammatory cytokine, TNF-α
(18), a major detrimental and
proapoptotic factor in this model of hepatitis (19). In the present study, it was
demonstrated that metformin significantly suppressed
LPS/D-Gal-induced coagulation activation, as indicated by the
reduced expression levels of TF and PAI-1, reduced level of HIF-1
and reduced content of LA in the liver.
The induction of TF serves a role in the activation
of coagulation under inflammatory circumstances (8). TF is constitutively expressed in the
majority of tissues, however is not expressed in cells in direct
contact with the blood (14). Once
the integrity of a vessel wall is disrupted, TF comes into contact
with other coagulation factors in the blood and initiates
coagulation (14). Additionally,
the expression of TF may be induced by inflammatory stimuli.
Therefore, the cells in direct contact with the blood, such as
monocytes and endothelial cells, may be induced to express TF under
inflammatory circumstances (13).
Several pro-inflammatory cytokines have been suggested to mediate
the induction of TF expression, however IL-6 may be the most
important mediator, as inhibition of IL-6 reverses TF-dependent
thrombin generation in experimental endotoxemia (22,31).
In the present study, the level of IL-6 and TF increased
significantly in mice challenged with LPS/D-Gal, however this
upregulation was suppressed by metformin, suggesting that the key
steps in the activation of the coagulation system were inhibited by
metformin.
The formation of thrombin may be counter-regulated
by the plasmin-catalyzed fibrinolysis process (32). Plasmin is generated from its
precursor plasminogen and this conversion is stimulated by two
activators, urokinase-type and tissue-type plasminogen activator
(32), which are inhibited by
PAI-1 (33). Additionally, PAI-1
is associated with inflammation due to the previously reported
potential upregulation of expression of PAI-1 by inflammatory
stimuli (34). In the present
study, the LPS/D-Gal-induced expression of PAI-1 was significantly
reduced by metformin, suggesting that the suppression of
fibrinolysis may be reversed by metformin. Previous studies have
demonstrated that TNF-α and IL-6 contribute to the increased
expression of PAI-1 under inflammatory conditions (34). In addition, the induced production
of TNF-α has been demonstrated to be significantly inhibited by
metformin (18), with the present
study indicating that the expression of IL-6 was also suppressed.
These anti-inflammatory effects of metformin may result in reduced
expression of PAI-1.
The transcription factor NF-κB controls the
expression of numerous pro-inflammatory genes including TNF-α and
IL-6 (35). In unstimulated cells,
NF-κB is sequestered by its inhibitor, inhibitory κB (IκB)
(36). Inflammatory stimuli may
induce the degradation of IκB and the release of NF-κB, resulting
in the translocation of NF-κB into the nucleus, where it activates
the transcription of target genes (36). In the present study, the increased
level of nuclear NF-κB in LPS/D-Gal-challenged mice was partially
reversed by metformin, which may contribute to the
anti-inflammatory effect on TNF-α and IL-6 expression.
Additionally, there is evidence indicating that NF-κB is involved
in the transcriptional regulation of TF and PAI-1 (24,34).
Thus, the inhibitory effects on NF-κB activation may underlie the
reduced induction of TF and PAI-1.
The activation of the coagulation system may result
in the reduction of blood flow, which leads to tissue hypoperfusion
and hypoxia (12). In the present
study, the degree of hepatic hypoxia was determined via the
detection of the stable protein level of HIF-1α. HIF is an
oxygen-sensitive transcription factor containing an oxygen-labile
α-subunit and a constitutively expressed β-subunit (37). Under normoxic conditions, the
hydroxylation of HIF-1α by the prolyl-4-hydroxylase
domain-containing enzymes (PHD) targets HIF-1α for
polyubiquitination and proteosomal degradation (38). In hypoxic conditions, PHD activity
is inhibited and HIF-1α accumulates and binds to the
hypoxia-responsive element sequences of target gene promoters
(38). In mice exposed to
LPS/D-Gal, levels of the stable protein of HIF-1α were markedly
increased, suggesting the presence of severe hypoxia in the liver
tissues. Treatment with metformin significantly reduced the level
of HIF-1α protein in LPS/D-Gal-challenged mice. Additionally, the
LPS/D-Gal-induced expression of HIF-1-targeted genes, including
EPO, VEGF and MMP-3 (25–26), was suppressed following metformin
treatment. These data were consistent with the reduced level of TF.
The HIF-1-driven expression of harmful products such as MMP-3 may
represent one of the mechanisms involved in hypoxia-induced tissue
damage (39).
Furthermore, tissue hypoxia and hypoperfusion may
lead to metabolic disturbance. Hypoxia may block mitochondrial
oxidative phosphorylation resulting in the generation of ATP from
the anaerobic metabolic pathway. In the anaerobic metabolic
pathway, pyruvate is converted into LA instead of acetyl-coenzyme A
(40). In the present study, the
level of LA increased significantly following LPS/D-Gal exposure,
however treatment with metformin reduced the level of LA. Increased
levels of LA have been suggested to be a pathological factor
responsible for liver injury via the stimulation of the release of
hydrolytic lysosomal enzymes into the cytosol and other mechanisms,
including the induction of endothelial dysfunction (41,42).
Taken together, the results of the present study
further indicate that metformin significantly suppresses
LPS/D-Gal-induced NF-κB activation, and IL-6, TF and PAI-1
expression. These effects are suggested to be associated with
alleviated hepatic hypoxia and metabolic disturbance. Therefore,
the present study identified an additional mechanism underlying the
hepatoprotective effects of metformin in mice with
LPS/D-Gal-induced fulminant hepatitis. The suppressive effects of
metformin on inflammation and inflammation-induced coagulation may
contribute to the protective benefits of metformin.
Acknowledgments
The current study was supported by a grant from the
National Natural Science Foundation of China (grant no. 81370179)
and a grant from Chongqing Municipal Education Commission (grant
no. KJ1400235).
Abbreviations:
|
D-Gal
|
D-galactosamine
|
|
EPO
|
erythropoietin
|
|
HIF-1α
|
hypoxia inducible factor 1α
|
|
IL-6
|
interleukin 6
|
|
IκB
|
inhibitory κB
|
|
LA
|
lactic acid
|
|
LPS
|
lipopolysaccharide
|
|
MMP-3
|
matrix metalloproteinase-3
|
|
NF-κB
|
nuclear factor κB
|
|
PAI-1
|
plasminogen activator inhibitor-1
|
|
TF
|
tissue factor
|
|
TNF-α
|
tumor necrosis factor α
|
|
VEGF
|
vascular endothelial growth factor
|
References
|
1
|
Schiødt FV and Lee WM: Fulminant liver
disease. Clin Liver Dis. 7:331–349. vi2003. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bernal W, Auzinger G, Dhawan A and Wendon
J: Acute liver failure. Lancet. 376:190–201. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Tsutsui H, Adachi K, Seki E and Nakanishi
K: Cytokine-induced inflammatory liver injuries. Curr Mol Med.
3:545–559. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hatano E: Tumor necrosis factor signaling
in hepatocyte apoptosis. J Gastroenterol Hepatol. 22(Suppl 1):
S43–S44. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kim SJ and Lee SM: NLRP3 inflammasome
activation in D-galactosamine and lipopolysaccharide-induced acute
liver failure: Role of heme oxygenase-1. Free Radic Biol Med.
65:997–1004. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Li Y, Xu Z, Yu Y, Yuan H, Xu H, Zhu Q,
Wang C and Shi X: The vagus nerve attenuates fulminant hepatitis by
activating the Src kinase in Kuppfer cells. Scand J Immunol.
79:105–112. 2014. View Article : Google Scholar
|
|
7
|
Levi M, Schultz M and van der Poll T:
Sepsis and thrombosis. Semin Thromb Hemost. 39:559–566. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Aksu K, Donmez A and Keser G:
Inflammation-induced thrombosis: Mechanisms, disease associations
and management. Curr Pharm Des. 18:1478–1493. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Semeraro N, Ammollo CT, Semeraro F and
Colucci M: Sepsis, thrombosis and organ dysfunction. Thromb Res.
129:290–295. 2012. View Article : Google Scholar
|
|
10
|
O'Brien M: The reciprocal relationship
between inflammation and coagulation. Top Companion Anim Med.
27:46–52. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
de Laforcade A: Diseases associated with
thrombosis. Top Companion Anim Med. 27:59–64. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gando S: Microvascular thrombosis and
multiple organ dysfunction syndrome. Crit Care Med. 38(Suppl):
S35–S42. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Egorina EM, Sovershaev MA and Hansen JB:
The role of tissue factor in systemic inflammatory response
syndrome. Blood Coagul Fibrinolysis. 22:451–456. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Mackman N: The many faces of tissue
factor. J Thromb Haemost. 7(Suppl 1): 136–139. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Schouten M, Wiersinga WJ, Levi M and van
der Poll T: Inflammation, endothelium, and coagulation in sepsis. J
Leukoc Biol. 83:536–545. 2008. View Article : Google Scholar
|
|
16
|
Bergmann S and Hammerschmidt S:
Fibrinolysis and host response in bacterial infections. Thromb
Haemost. 98:512–520. 2007.PubMed/NCBI
|
|
17
|
Hermans PW and Hazelzet JA: Plasminogen
activator inhibitor type 1 gene polymorphism and sepsis. Clin
Infect Dis. 41(Suppl 7): S453–S458. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yuan H, Li L, Zheng W, Wan J, Ge P, Li H
and Zhang L: Antidiabetic drug metformin alleviates
endotoxin-induced fulminant liver injury in mice. Int
Immunopharmacol. 12:682–688. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Silverstein R: D-galactosamine lethality
model: Scope and limitations. J Endotoxin Res. 10:147–162.
2004.PubMed/NCBI
|
|
20
|
Miyazaki M, Kato M, Tanaka M, Tanaka K,
Takao S, Kohjima M, Ito T, Enjoji M, Nakamuta M, Kotoh K, et al:
Antithrombin III injection via the portal vein suppresses liver
damage. World J Gastroenterol. 18:1884–1891. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Takano H, Inoue K, Shimada A, Sato H,
Yanagisawa R and Yoshikawa T: Urinary trypsin inhibitor protects
against liver injury and coagulation pathway dysregulation induced
by lipopolysaccharide/D-galactosamine in mice. Lab Invest.
89:833–839. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kerr R, Stirling D and Ludlam CA:
Interleukin 6 and haemostasis. Br J Haematol. 115:3–12. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Liang DY, Liu LM, Ye CG, Zhao L, Yu FP,
Gao DY and Wang YY, Yang ZW and Wang YY: Inhibition of UII/UTR
system relieves acute inflammation of liver through preventing
activation of NF-κB pathway in ALF mice. PLoS One. 8:e648952014.
View Article : Google Scholar
|
|
24
|
Jiang R, Wang NP, Tanaka KA, Levy JH,
Guyton RA, Zhao ZQ and Vinten-Johansen J: Factor Xa induces tissue
factor expression in endothelial cells by P44/42 MAPK and
NF-κB-dependent pathways. J Surg Res. 169:319–327. 2011. View Article : Google Scholar
|
|
25
|
Rhim T, Lee DY and Lee M: Hypoxia as a
target for tissue specific gene therapy. J Control Release.
172:484–494. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ahn JK, Koh EM, Cha HS, Lee YS, Kim J, Bae
EK and Ahn KS: Role of hypoxia-inducible factor-1alpha in
hypoxia-induced expressions of IL-8, MMP-1 and MMP-3 in rheumatoid
fibroblast-like synoviocytes. Rheumatology (Oxford). 47:834–839.
2008. View Article : Google Scholar
|
|
27
|
Bailey CJ and Turner RC: Metformin. N Engl
J Med. 334:574–579. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Tsoyi K, Jang HJ, Nizamutdinova IT, Kim
YM, Lee YS, Kim HJ, Seo HG, Lee JH and Chang KC: Metformin inhibits
HMGB1 release in LPS-treated RAW 264.7 cells and increases survival
rate of endotoxaemic mice. Br J Pharmacol. 162:1498–1508. 2011.
View Article : Google Scholar :
|
|
29
|
Hattori Y, Suzuki K, Hattori S and Kasai
K: Metformin inhibits cytokine-induced nuclear factor kappaB
activation via AMP-activated protein kinase activation in vascular
endothelial cells. Hypertension. 47:1183–1188. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Nath N, Khan M, Paintlia MK, Singh I, Hoda
MN and Giri S: Metformin attenuated the autoimmune disease of the
central nervous system in animal models of multiple sclerosis. J
Immunol. 182:8005–8014. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
van der Poll T, Levi M, Hack CE, ten Cate
H, van Deventer SJ, Eerenberg AJ, de Groot ER, Jansen J, Gallati H,
Büller HR, et al: Elimination of interleukin 6 attenuates
coagulation activation in experimental endotoxemia in chimpanzees.
J Exp Med. 179:1253–1259. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wyseure T and Declerck PJ: Novel or
expanding current targets in fibrinolysis. Drug Discov Today.
19:1476–1482. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Dupont DM, Madsen JB, Kristensen T, Bodker
JS, Blouse GE, Wind T and Andreasen PA: Biochemical properties of
plasminogen activator inhibitor-1. Front Biosci (Landmark Ed).
14:1337–1361. 2009. View
Article : Google Scholar
|
|
34
|
Kruithof EK: Regulation of plasminogen
activator inhibitor type 1 gene expression by inflammatory
mediators and statins. Thromb Haemost. 100:969–975. 2008.
|
|
35
|
Hoesel B and Schmid JA: The complexity of
NF-κB signaling in inflammation and cancer. Mol Cancer. 12:862013.
View Article : Google Scholar
|
|
36
|
Lee IT and Yang CM: Inflammatory
signalings involved in airway and pulmonary diseases. Mediators
Inflamm. 2013:7912312013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Ong SG and Hausenloy DJ: Hypoxia-inducible
factor as a therapeutic target for cardioprotection. Pharmacol
Ther. 136:69–81. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Brocato J, Chervona Y and Costa M:
Molecular responses to hypoxia-inducible factor 1α and beyond. Mol
Pharmacol. 85:651–657. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Ramakrishnan P, Hecht BA, Pedersen DR,
Lavery MR, Maynard J, Buckwalter JA and Martin JA: Oxidant
conditioning protects cartilage from mechanically induced damage. J
Orthop Res. 28:914–920. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Levy B: Lactate and shock state: The
metabolic view. Curr Opin Crit Care. 12:315–321. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Soric S, Belanger MP, Askin N and Wittnich
C: Impact of female sex hormones on liver tissue lactic acidosis
during ischemia. Transplantation. 84:763–770. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
James JH, Luchette FA, McCarter FD and
Fischer JE: Lactate is an unreliable indicator of tissue hypoxia in
injury or sepsis. Lancet. 354:505–508. 1999. View Article : Google Scholar : PubMed/NCBI
|